本内苏铁:最早出现的两性"花"
2017-11-02许旭
许旭
本内苏铁:最早出现的两性"花"
许旭
1.苏铁植物门类
在中生代,裸子植物达到繁盛的顶峰期,其中苏铁类植物(Cycadophyta)广泛分布全球,最具代表性,成为恐龙森林的主要的构建者,中生代又被称为"苏铁植物时代"。
苏铁植物包含有苏铁纲(Cycadopsida)和本内苏铁纲(Bennettitopsida)。苏铁纲植物虽在侏罗纪和早白垩世极为繁盛,分布遍及全球,但现今已处于次要地位(图1),仅存留10属近180种,分布于热带、亚热带地区,很多种类被广泛用于园林绿化及作为庭院植物。本内苏铁则常见于晚三叠世至早白垩世植物群,在西欧、英格兰、格陵兰岛、加拿大、阿拉斯加、墨西哥南部、南非、印度、澳大利亚、中国等地层中广泛分布。最近发现个别分子在冈瓦纳地区或可残存至渐新世,至今已全部灭绝。发掘本内苏铁的秘密,完全依靠于对化石证据的研究,这使本内苏铁变得神秘而又奇特。

图1:现生的苏铁纲植物 (http://bbs.zol.com.cn/dcbbs/d34039_1326.html)
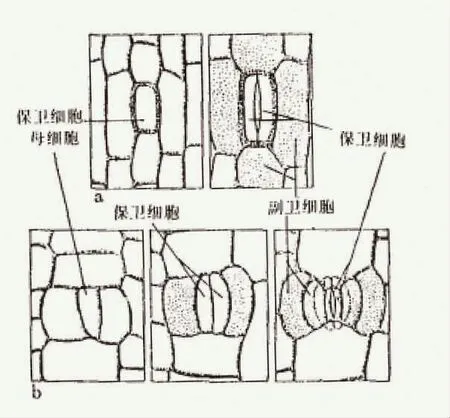
图2:裸子植物气孔主要类型,a-单唇型,b-复唇式 (A.S.Foster and E.M.Gifford,1974)
本内苏铁纲有学者也称为拟苏铁纲(Cycadeoidopsida),仅一目:本内苏铁目(Bennettitales)或拟苏铁目(Cycadeoidales)。本内苏铁从外观上与苏铁纲难以区别:茎不分枝或稍有分枝,有的矮小粗壮或成块状,也有的成细枝状,粗壮的茎上有连续叶痕武装的盔甲;叶为一次羽状复叶,革质而具旱生结构。疏木质木材,单沟型花粉,直生胚珠等也是两者相似之处。尽管如此,本内苏铁复唇式的气孔器与两性"花"的生殖器官等特征,仍可使其与苏铁纲严格区分开来。
2.本内苏铁植物结构

图3:昆虫与本内苏铁植物相互作用(http://www.telegiz.com/articles/2402/20160205/jurassic-butterfly-appeared-40-millionyears-earlier-butterflies.htm)

图4:本内苏铁目植物常见叶形 (Watson and Sincock,1992)
本内苏铁纲的气孔(图2b)为复唇式(syndetocheilic),也称本内苏铁式,其始原细胞即形成保卫细胞,也形成副卫细胞,这种气孔结构也出现在现存裸子植物买麻藤属(gnetum)、千岁兰属(welwitschia)、一些被子植物和原始的石松植物上。同时本内苏铁叶片的表皮细胞明显排列成行且细胞壁弯曲。而苏铁纲的气孔器(图2a)为单唇式(haplocheilic),也称苏铁式,即表皮始原细胞仅发育成保卫细胞,而副卫细胞则由表皮细胞形成,这类气孔结构也连续出现在种子蕨、科达、松柏、银杏属(Ginkgo)和麻黄属(Ephedra)的表皮上。在某些木兰植物中,叶片的气孔类似于本内苏铁,而花上的气孔则是苏铁的类型。有学者认为,气孔类型的不同,意味着进化程度的不同。

图5:云南省禄丰县一平浪植物群中的异羽叶(Anomozamites)
而本内苏铁纲与苏铁纲的最大区别就是雌雄同株,具有与被子植物十分相近的两性"花"。这并不是真正的花,而是两性的生殖器官融合到了一起,产生花粉的小孢子叶着生在具有胚珠的花托基部周围,长成类似于"花"的结构。它可能的彩色类花瓣结构可吸引早期的昆虫为它传粉(图3)。这种特殊的生殖器官是植物进化史上最早出现的两性"花",而它在研究被子植物起源的课题上尤为重要,Wieland(1906)就曾将本内苏铁假定为被子植物的祖先。而苏铁纲植物就显得更为原始:雌雄异株,雌雄孢子叶球形体都比较大,多数着生于茎的顶端。
本内苏铁常见的叶片化石属(图4)有侧羽叶(Pterophyllum)、毛羽叶(Ptilophyllum)、似查米亚(Zamites)、耳羽叶(Otozamites)、网羽叶(Dictyozamites)、异羽叶(Anomozamites)(图 5)和尼尔桑带羽叶(Nilssoniopteris)。这些叶片大部分为一次羽状复叶,裂片规则,平行脉或放射脉。本内苏铁的孢子叶属为角羽叶(Deltolepis)和苏铁鳞片(Cycadolepis)。
本内苏铁茎干的内部有很大的髓部居于中心,髓部被一个圆筒状且发育良好的木质部围绕。次生木质部通常紧实,类似于松柏。初生木质部内始式,即原生木质部从维管柱最内面的原形成层细胞开始发育,并向外分化,这也是最进化的类型。叶迹源于初生木质部并直接穿过次生木质部进入皮层,分支形成若干痕迹以补给叶基。不出现环形叶迹,大量紧凑的次生木质部和次生木质部的梯形管胞,是本内苏铁区别于苏铁纲的重要性征。
3.本内苏铁植物类型

图 6:Williamsonia3D 重建图(https://fr.wikipedia.org/wiki/Bennettitales)

图7:Williamsonia sewardiana重建图(Courtesy T.Delevoryas.)
本内苏铁目植物根据地质年代、生态、生殖器官组织结构的不同可分为拟苏铁科(Cycadeoidaceae) 和威廉姆逊科(Williamsoniaceae)。
(1) 威廉姆逊科(Williamsoniaceae)
威廉姆逊科较为古老,出现在晚三叠世至侏罗纪的地层中。包含在这个科的植物拥有细长分支的茎干,这代表了某些种子蕨的原始分支类型。这些植物推测高达2米,单性球果或有的与拟苏铁科一样为两性"花"。按照生殖器官特征,本科植物分为以下属型:①威廉姆逊属(Williamsonia)
威廉姆逊属(图6)是单一孢子囊的,短的侧枝上长有具胚珠的大孢子叶球("雌花"),Williamsonia一名也是指代"雌花"的形态属,花柄上有圆锥形花托,胚珠为种间鳞片包围,花托下部具鳞叶。学者在一些胚珠的空腔里还发现有雌配子体和双子叶的胚芽。"雄花"的形态、结构类似拟苏铁科,常用Weltrichia指代,小孢子叶(雄蕊)的下部连结成漏斗状,但它如何着生于枝干上尚不清楚。
在威廉姆逊科,本属可谓是最为吸引目光,这是部分是由于Sahi(1932)富有想象力而美丽的复原图 -Williamsonia sewardiana(图 7),整株植物的重建均基于碎片化的化石证据。这是在印度侏罗纪矿化保存的植物,它1.5到2米高,呈小树状,树干表面具永久性的螺旋排列叶基,在顶部形成毛羽叶型(Ptilophyllum)羽状复叶形成的树冠,枝干则为Bucklandia indica型,长有最终球果的枝条被苞片覆盖,出现在叶基之中,球果类型为Williamsonia sewardiana。另外,本科复原植物还有长有似查米亚(Zamites)叶片类型的 Williamsonia gigas;Harris(1969)的重建植物:树干为Bucklandia pustulosa,生殖器官为 Williamsonia leckenbyi,叶片为 Ptilophyllum pectin;枝干上长有若干广泛螺旋排列的耳羽叶(Otozamites)与似查米亚(Zamites)类型叶片的Ischnopython。

图 8:Wielandiella angustifolia 复原图 (Pott,2014)
本属时代为侏罗纪至早白垩纪。
②魏兰德属(Wielandiella)与小威廉姆逊属(Williamsoniella)

图9:Cycadeoidea Gigantea 3D复原图(https://www.cgtrader.com/3d-models/plant/other/xfrogplants-cycadeoidea-gigantea)

图10:Cycadeoidea的球果重建:or为胚珠花托;mi为小孢子叶(Crepet,1974)
魏兰德(Wielandiella)是威廉姆逊科第一个发现的化石记录,发现于瑞士晚三叠世地层,小威廉姆逊(Williamsoniella)则发现于早侏罗世,它们纤细而光滑的枝呈二歧式分枝,前者叶为异羽叶(Anomozamites)型,后者叶为尼尔桑带羽叶(Nilssoniopteris)型,但二者均具两性"花",小孢子叶的基部亦相连,但不成漏斗状。各大孢子叶间有间生鳞片。而魏兰德属(图8)最近又在我国内蒙古中侏罗世地层中发现,由Pott(2015)命名并描述为新种:Wielandiella viillosa。
(2)拟苏铁科(Cycadeoidaceae)
本科出现于全球晚侏罗世至白垩纪地层中,树干多不分支,矮胖,不超过50cm高,约40cm宽,椭球形或球形。有很多化石爱好者找到拟苏铁科的树干化石,认为这些有趣的标本是蜂窝化石,珊瑚,蘑菇或其他多种多样的结构。
拟苏铁属(Cycadeoidae)是本科研究最清楚的属(图9):茎成椭球形或圆柱状,上覆永久性叶基及小鳞片。叶为似查米亚(Zamites)型,螺旋状长在树干顶端。"花"长在叶腋间,在一个较粗的轴上,最外面为密螺旋状排列的羽状营养苞片,苞片上密覆茸毛,苞片之上环状排列基部联合的雄蕊(小孢子叶),轴的顶端为"雌花"(大孢子叶球)。"雌花"是由许多螺旋状排列的具柄胚珠和顶端膨大的间生鳞片组成。间生鳞片有保护胚珠的作用。
很幸运的是,由于保存精美的矿化化石,Crepet(1974)揭示了小孢子叶早期的生长过程(图10):早期的小孢子叶围聚在球果顶端的分生组织周围,当他们成熟时,羽状的小孢子叶便会外卷,带有聚合囊的小羽片向平行于叶轴的平面翻折。这些小羽片的轴在胚珠花托的顶端融合。而胚珠在前配子体阶段四分裂的大孢子的化石记录也有被发现,可惜的是它的成熟过程并没被人类了解。
4.本内苏铁的起源与灭绝
图11:本内苏铁球果起源于生殖枝的演化假说图示
仅仅从单独的植物学特征来判断,苏铁纲和本内苏铁纲植物确实是密切相关的两个类群,但考虑到球果的类型,它们的关系就比较遥远了。而对于这最好的理解就是将它们看作是有着共同祖先-古生代某类种子蕨的两个分离的类群。根据拟苏铁属和威廉姆逊属树干的内部构造,将髓木类的种子蕨作为是它们共同的来源是我们最好的选择。不规律叶痕位置是髓木类种子蕨、苏铁纲,本内苏铁纲共同的维管束性征。而大小孢子囊的球果可能来源于具有大小孢子囊的种子蕨,简单来讲,就是带有小孢子和胚珠的生殖叶片压缩聚合成球果,而本内苏铁的球果就是一个被压缩的腋生生殖枝(图11)。有学者指出:本内苏铁的聚合囊结构来源于拥有髓木类树冠植物的棒状聚合囊的融合。但胚珠的双层珠被,却与髓木类不同,更相近于有花植物和开通目(Caytoniales)。种间鳞片和胚珠的起源,至今众说纷纭,没有一个统一的答案。但如果忽略这个问题,我们便可以更轻松的说:髓木目就是苏铁植物门的起源。
学者们却一直存在这样的怀疑:本内苏铁目比起和苏铁目,与买麻藤目(Gnetales)和有花植物更具亲缘关系,将它们放在苏铁植物门并不符合自然的规律。这为弄清楚本内苏铁植物的分类位置又增添了一层疑云。
Crepet(1974)指出:本内苏铁球果上昆虫钻孔的痕迹和小孢子叶成熟不张开的证据,表明本内苏铁存在昆虫传粉和自花传粉两种机制,这两种传粉机制都可能对本内苏铁植物白垩纪的迅速衰退起了作用:一方面,与异花传粉的杂交植物相比,自花传粉的植物更加不能适应剧烈的气候变化;另一方面,随着被子植物的迅速发展,进化出了更能吸引昆虫的器官;昆虫传粉者转向了被子植物,对本内苏铁植物造成了沉重的打击。
