干旱胁迫对刺槐、皂荚叶片渗透调节物质含量及保护酶活性的影响
2017-10-23陈丽培刘瑞霞杨玉珍王国霞雒红宇
陈丽培,刘瑞霞,杨玉珍,王国霞,雒红宇
(郑州师范学院 生命科学学院,河南 郑州 450044)
干旱胁迫对刺槐、皂荚叶片渗透调节物质含量及保护酶活性的影响
陈丽培,刘瑞霞,杨玉珍,王国霞,雒红宇
(郑州师范学院 生命科学学院,河南 郑州 450044)
以刺槐(Robiniapseudoacacia)与皂荚(Gleditsiasinensis)的1年生苗为材料,通过盆栽控水试验,研究干旱胁迫对2种苗木叶片中渗透调节物质含量、丙二醛(MDA)含量及保护酶活性的影响。结果表明:随着干旱胁迫强度的增大,2种苗木叶片的脯氨酸(Pro)含量、可溶性糖含量比正常水分时升高,相同水分梯度下刺槐Pro、可溶性糖含量升高幅度较大;皂荚叶片中可溶性蛋白含量增加,刺槐叶片中可溶性蛋白含量降低;MDA含量随水分胁迫程度的加剧均表现出升高趋势,且皂荚在相同水分梯度下增幅较大;超氧化物歧化酶(SOD)活性增加,但刺槐增加幅度大于皂荚;皂荚叶片中过氧化物酶(POD)活性增加、过氧化氢酶(CAT)活性降低,而刺槐叶片中CAT活性增加、POD活性降低。可知,2种苗木通过渗透调节物质的积累、抗氧化酶活性的升高来增强对干旱胁迫的适应性。总体来看,刺槐耐干旱能力大于皂荚。
皂荚; 刺槐; 干旱胁迫; 渗透调节物质; 保护酶活性
水分是植物的重要组成物质,也是影响植物生长发育的重要因子之一[1]。水分亏缺时,植物会自动开启保护酶体系及渗透调节功能来适应逆境。目前,我国对植物干旱胁迫的生理响应研究较多,主要针对水分不足时渗透调节物质及保护酶活性的变化。陈吉虎等[2]通过研究干旱胁迫下5个树种苗木抗氧化酶活性的变化,确定它们的抗旱能力。项忠阳[3]通过测定水分胁迫下刺槐和紫穗槐相对含水量、水分饱和亏缺、脯氨酸(Pro)含量、丙二醛(MDA)含量、超氧化物歧化酶(SOD)活性的变化,揭示出紫穗槐的抗旱性强于刺槐。徐莲珍等[4]研究3种苗木叶片在遭受干旱胁迫时渗透调节物质含量、保护酶活性的变化,探讨了生理指标与其抗旱性的关系。皂荚与刺槐同为豆科(Leguminosae)落叶乔木,为具有多种用途的生态经济林树木。它们根系发达,具有较强繁殖能力和耐干旱能力,为防护林及行道树的主要树种。在现有抗旱机制研究基础上,采用盆栽试验,以郑州市1年生皂荚与刺槐苗为供试材料,探讨干旱胁迫条件下其叶片渗透调节物质含量及抗氧化酶活性等的变化,为2种苗木的栽培和推广,以及北方干旱地区选取优良抗旱树种提供理论依据。
1 材料和方法
1.1材料与处理
以皂荚(Gleditsiasinensis)、刺槐(Robiniapseudoacacia)2个树种1年生苗为试验材料。2016年3月初将苗木移栽到外口直径48 cm、内口直径42 cm、高33 cm、底部直径27 cm的塑料盆中,花盆土壤为营养土和校园苗圃表土1∶1的混合土。将花盆放在防雨棚中遮雨并进行正常水分管理。2016年5月4日选取长势基本一致的苗木进行试验。试验前通过称质量的方法设定4个水分梯度,土壤含水量分别为75%~80%(正常供水)、55%~60%(轻度胁迫)、40%~45%(中度胁迫)、30%~35%(重度胁迫)。每天18:00左右补充蒸发掉的水量,使土壤水分一直稳定在设定的范围内。控水处理21 d时,于6:00—7:00选取完整、无病虫害叶片,测定Pro、可溶性蛋白、可溶性糖、MDA含量及SOD、过氧化物酶(POD)、过氧化氢酶(CAT)活性。测量时每个水分梯度的2种苗木随机各选取8株,进行3次重复。
1.2测定指标及方法
Pro含量、可溶性蛋白含量、可溶性糖含量测定分别采用酸性茚三酮显色法[5]、考马斯亮蓝G-250法[6]、蒽酮比色法[7];MDA含量测定采用TBA比色法[8];SOD活性、POD活性、CAT活性测定分别采用氮蓝四唑(NBT)比色法[8]、愈创木酚显色法[5]和紫外吸收法[6]。
2 结果与分析
2.1干旱胁迫对2种苗木叶片渗透调节物质的影响
2.1.1 Pro Pro作为植物细胞内主要渗透调节物质,干旱条件下可以降低细胞渗透势,增强从周边环境吸水能力,提高植物抗旱性[9]。由图1可知,2种苗木叶片Pro含量随着干旱胁迫程度的加剧均呈升高趋势。水分轻度胁迫时,刺槐、皂荚Pro含量为170.68、108.12 μg/g,分别比正常水分升高了69.60%、38.97%。水分中度胁迫时,皂荚Pro含量达到最大值(209.80 μg/g),比正常水分增加了169.67%,之后缓慢降低,重度胁迫时Pro含量为198.60 μg/g,仍比正常水分升高155.27%。刺槐Pro含量中度胁迫时为334.03 μg/g,比正常水分增加了233.41%,之后缓慢上升,重度胁迫时达到最高值(343.9 μg/g),比正常水分增加了241.75%。刺槐、皂荚由轻度胁迫至中度胁迫时增加的Pro含量为正常水分时的162.34%、130.60%。可见,2种苗木在水分亏缺时的Pro含量积累顺序为中度胁迫>轻度胁迫。重度胁迫时,Pro合成受到抑制,变化不明显。另外,在相同水分梯度下,刺槐Pro含量升高幅度大于皂荚,刺槐Pro在干旱条件下渗透调节能力更强。

图1 干旱胁迫对刺槐、皂荚叶片Pro含量的影响
2.1.2 可溶性糖 可溶性糖主要包括葡萄糖、蔗糖、海藻糖等,它们不仅为植物的生长代谢提供物质和能量,还在苗木抵抗干旱时起着重要的调节作用。在干旱胁迫下,2种苗木叶片可溶性糖的含量均高于正常供水情况,且都表现出先升高再降低的趋势(图2)。刺槐、皂荚轻度胁迫时可溶性糖含量分别为71.74、58.28 mg/g,比正常水分时升高了70.24%、52.85%。随着胁迫程度加剧,2种苗木的可溶性糖含量在中度胁迫时均达到峰值87.15 mg/g和66.83 mg/g,比正常水分时增加了106.81%、75.27%。重度胁迫时刺槐、皂荚可溶性糖含量下降为79.36、53.96 mg/g,仍比正常水分时升高88.32%、41.52%。刺槐、皂荚由轻度胁迫至中度胁迫时增加的可溶性糖含量为正常水分的37.05%、22.42%。由此可知,2种苗木干旱胁迫时可溶性糖含量在轻度胁迫时积累最多,这与Pro相反。相同的胁迫程度时,刺槐可溶性糖含量升高幅度明显大于皂荚,这与Pro相似。总的来说,可溶性糖作为渗透调节物质,其含量的增加,提高了刺槐、皂荚水分亏缺时的渗透调节能力。
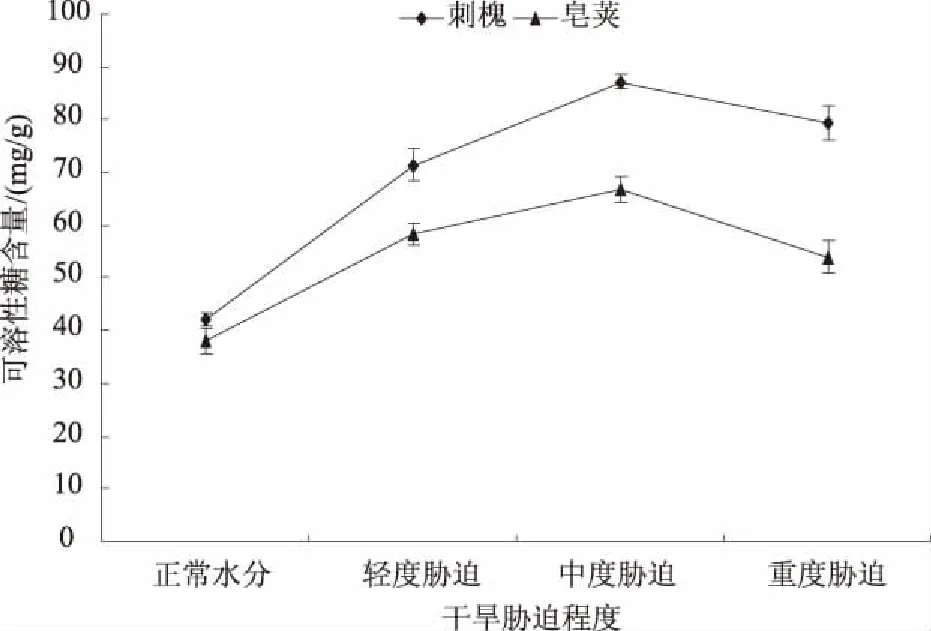
图2 干旱胁迫对刺槐、皂荚叶片可溶性糖含量的影响
2.1.3 可溶性蛋白 可溶性蛋白是构成生命的基础物质[10]。植物受到干旱胁迫时,可溶性蛋白含量的升高,有利于增加细胞渗透物质含量,稳定细胞体内渗压平衡,保护植物细胞不受伤害。图3表明,刺槐叶片的可溶性蛋白含量随土壤含水量的降低呈不同程度的下降趋势。水分轻度胁迫时,可溶性蛋白含量由29.40 mg/g降为28.20 mg/g,变化较小。从中度胁迫开始,降幅增大,到重度胁迫时降为最低值。中度胁迫、重度胁迫时可溶性蛋白含量分别比正常水分时降低了18.23%、44.86%。这可能是由于植物在水分胁迫时mRNA转录及翻译过程受到破坏[11],新的蛋白质合成受抑制,同时原来蛋白质分解速度加快所致。皂荚叶片可溶性蛋白含量表现出升高的趋势。轻度胁迫时可溶性蛋白含量为26.29 mg/g,比正常水分升高了10.76%。由轻度胁迫至中度胁迫时增加的可溶性蛋白含量为正常水分时的30.43%,而由中度胁迫至重度胁迫时增加的可溶性蛋白含量为正常水分时的14.00%。由此得出,由轻度胁迫至中度胁迫时皂荚可溶性蛋白累积量最多。皂荚叶片可溶性蛋白含量的升高,提高了皂荚叶片的保水力。
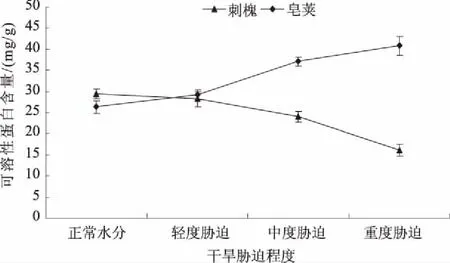
图3 干旱胁迫对刺槐、皂荚叶片可溶性蛋白含量的影响
2.2干旱胁迫对2种苗木叶片保护酶活性的影响
2.2.1 SOD SOD为自由基清除剂,广泛存在植物体中,能清除干旱胁迫下产生的有害物质,减轻活性氧对细胞膜脂的伤害,保证植物各种代谢活动的正常进行[12]。从干旱胁迫下2种苗木叶片SOD活性测定结果(图4)可看出,干旱胁迫下,刺槐和皂荚幼苗叶片SOD活性均高于正常水分时活性,且均表现出先升高再降低的趋势。轻度胁迫时2种苗木SOD活性分别为980.46、607.03 U/(g·h),分别比正常水分时升高了74.20%、50.70%。中度胁迫时SOD活性变化较小,缓慢上升为1 080.08、608.56 U/(g·h),增加的SOD活性分别为正常水分时的17.70%、0.38%。重度胁迫下,刺槐和皂荚SOD活性迅速下降,重度胁迫时降低的SOD活性为正常水分时的38.61%、47.55%。可知,在一定干旱范围内植物会主动产生SOD,有效地清除活性氧,对细胞起保护作用。但超过一定范围时,SOD活性下降。另外,虽然2种苗木SOD活性变化趋势一致,但变化幅度却有差异。在轻度胁迫时,SOD活性升高幅度为刺槐大于皂荚,重度胁迫时刺槐SOD活性降低幅度小于皂荚,重度胁迫两者变化都不明显。总的来说,干旱胁迫下刺槐SOD活性强于皂荚。
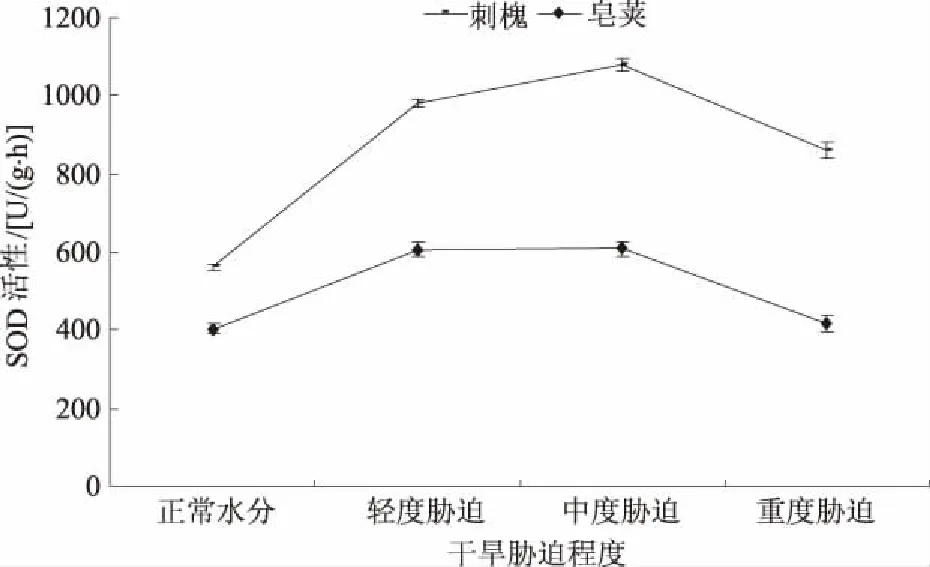
图4 干旱胁迫对刺槐、皂荚叶片SOD活性的影响
2.2.2 POD POD是一种活性较高的酶,干旱条件下,植物体内POD活性的升高有利于提高细胞的抗氧化能力。水分胁迫下,刺槐叶片POD活性呈不同程度下降趋势(图5)。其中,轻度胁迫时,POD活性下降较小,由881.16 U/(g·min)降低为868.58 U/(g·min),降低了1.43%。从中度胁迫开始下降明显,中度胁迫、重度胁迫时POD活性分别比正常水分降低了14.17%、40.48%。皂荚叶片POD活性表现出先升高再降低的趋势,但是干旱胁迫下的POD活性始终高于正常水分时活性。轻度胁迫时,POD活性由917.43 U/(g·min)升高为1 018.70 U/(g·min),升高了11.04%,之后迅速升高,中度胁迫时达到最大值1 385.28 U/(g·min),比正常水分时升高了51.00%。中度胁迫增加的POD活性为正常水分的39.96%。重度胁迫时POD活性有所下降,但仍比正常水分时增加了32.10%。表明POD在皂荚叶片中发挥着重要保护作用。水分亏缺时,POD活性增加幅度顺序为中度胁迫大于轻度胁迫。
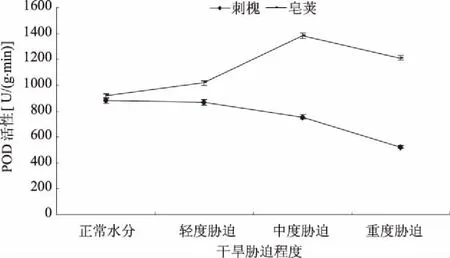
图5 干旱胁迫对刺槐、皂荚叶片POD活性的影响
2.2.3 CAT CAT作为一种末端氧化酶,在干旱胁迫时和POD作用一致,催化植物细胞中的H2O2使其分解,保护细胞免受高含量H2O2的伤害。图6表明,随着干旱胁迫程度的加剧,刺槐、皂荚叶片CAT活性均呈先升高再降低的趋势。其中,刺槐叶片在轻度胁迫时,CAT活性增加了25.73%,之后迅速升高,中度胁迫时达到峰值[646.23 U/(g·min)],比正常水分时升高了67.90%。中度胁迫CAT活性增加量为正常水分时的42.13%。重度胁迫时,CAT活性降低为596.06 U/(g·min),仍比正常水分时增加了54.86%。皂荚CAT活性在轻度胁迫时最高[262.92 U/(g·min)],比正常水分升高了1.88%。之后呈不同程度下降趋势。重度胁迫时降为最低值206.57 U/(g·min),比正常水分降低了19.96%。皂荚叶片CAT活性在干旱情况下变化较小,基本保持在206.57~262.92 U/(g·min)。刺槐叶片CAT活性变化趋势明显,干旱条件下,CAT活性增加幅度顺序为中度胁迫>轻度胁迫,这与皂荚叶片POD活性变化一致。虽然刺槐CAT活性在重度胁迫时下降,但仍然高于正常水分活性。可见,刺槐叶片CAT在干旱条件下起着重要的抗氧化作用。
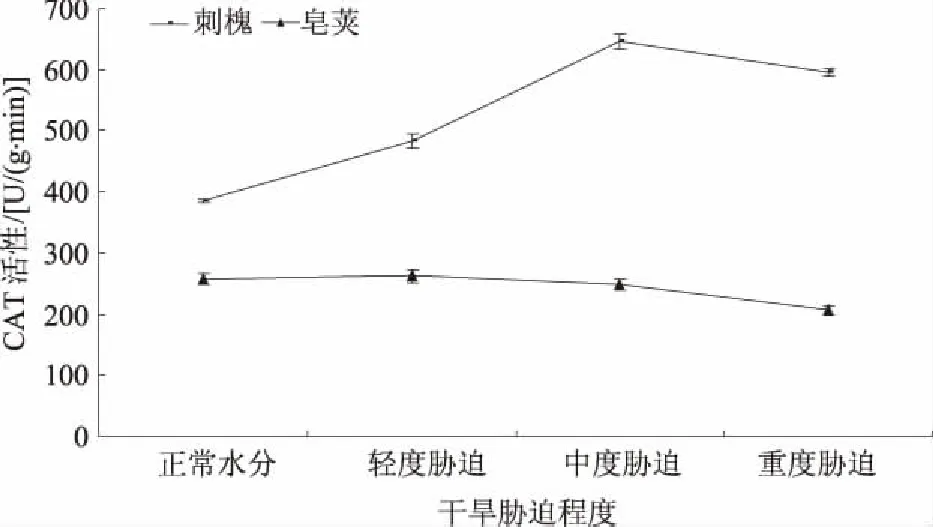
图6 干旱胁迫对刺槐、皂荚叶片CAT活性的影响
2.3干旱胁迫对2种苗木叶片MDA含量的影响
MDA是膜脂过氧化产物之一,对细胞膜破坏性很强[13]。干旱胁迫时,MDA含量的变化可作为反映植物受胁迫严重程度的重要指标。由图7可知,随着干旱胁迫程度的加剧,2种苗木叶片MDA含量均表现出上升的趋势,但变化幅度有所差异。刺槐、皂荚在轻度胁迫时MDA含量分别为7.64、10.96 nmol/g,比正常水分时升高了6.11%、11.04%。从中度胁迫起MDA含量升高幅度变大,2种苗木中度胁迫时增加的MDA含量为正常水分的17.08%、30.40%。重度胁迫时刺槐、皂荚苗木MDA含量均达到最大值11.06、19.20 nmol/g,增加的MDA含量为正常水分的30.42%、53.00%。可见,在干旱条件下2种苗木MDA含量的增幅顺序为重度胁迫>中度胁迫>轻度胁迫,这与SOD活性增幅顺序相反(图4)。另外,在相同水分梯度时,皂荚苗木MDA含量升高幅度大于刺槐,皂荚膜脂过氧化程度强于刺槐。
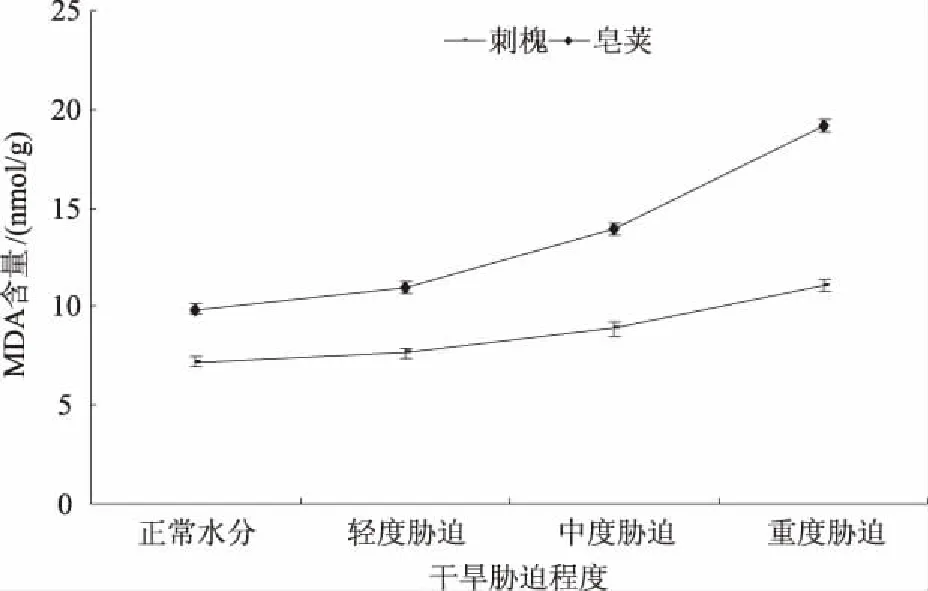
图7 干旱胁迫对刺槐、皂荚叶片MDA含量的影响
3 结论与讨论
渗透调节是植物长期进化的一种干旱适应机制。植物遭受水分胁迫时,通过主动产生一些渗透调节物质来降低渗透势,增强植物的抗干旱能力。Pro、可溶性糖、可溶性蛋白作为干旱胁迫的渗透调节物质,其含量是衡量植物抗旱性的重要生理指标。关于干旱胁迫对渗透调节物质的影响研究较多,一般认为,植物缺水时,Pro、可溶性糖、可溶性蛋白含量均表现一定程度的升高,从而增加植物的干旱适应性。王海珍等[14]研究干旱胁迫对灰胡杨(PopuluspruinosaSchrenk)和胡杨(PopuluseuphraticaOliv)幼苗渗透调节物质影响时发现,干旱胁迫下灰胡杨、胡杨幼苗通过渗透调节物质的积累来抵御水分胁迫;李燕等[15]认为,皂角幼苗表现出一定的抗旱性,可能与渗透调节物质含量的升高有关;李州等[16]、刘晓东等[17]研究发现,白三叶(Trifoliumrepens)、玉带草(Phalarisarundinaceavar.picta)叶片随着水分胁迫程度的加剧,叶片内游离Pro含量呈升高趋势,从而使叶片保持相对较高的渗透调节能力,以保证水分的合理利用;包卓等[18]对5种园林绿化植物研究表明,干旱胁迫下,具有较强抗旱能力的蛇莓(Duchesneaindica)和蛇莓委陵菜(Potentillacentigrana)叶片可溶性糖和可溶性蛋白含量增加。本研究中,刺槐、皂荚在土壤含水量降低过程中,叶片Pro含量表现出升高的趋势。可溶性糖含量随着干旱胁迫程度的增强均呈先升高再降低的趋势。重度胁迫后2种苗木可溶性糖含量有所下降,但仍比正常水分升高了88.32%、41.52%。说明Pro和可溶性糖在刺槐、皂荚干旱时均具有较强的调节作用。但是水分亏缺时不同植物对Pro和可溶性糖的敏感程度不同。由试验结果可知,在相同干旱梯度下,刺槐的Pro、可溶性糖含量升高幅度均大于皂荚,可见干旱胁迫对Pro、可溶性糖含量的影响顺序为刺槐大于皂荚。另外,2种苗木在水分不足时Pro大量积累发生在中度胁迫,而可溶性糖在轻度胁迫时大量积累。可以得出,可溶性糖积累早于Pro,可溶性糖对干旱胁迫的敏感度大于Pro。可溶性蛋白作为植物正常情况下催化各种生理生化过程的酶类,在干旱情况下参与植物的渗透调节。本研究中,刺槐可溶性蛋白含量在干旱情况下逐渐降低,重度胁迫降为最低值,显著低于正常水分。皂荚叶片可溶性蛋白含量变化趋势刚好相反,随土壤含水量的降低而升高。说明不同树种遭受干旱胁迫时参与渗透调节的物质不同,可溶性蛋白是皂荚叶片的渗透调节物质之一。有学者发现,干旱胁迫时可溶性蛋白含量的升高与植物合成新的蛋白质有关[12]。孙国荣等[19]也认为,干旱胁迫下可溶性蛋白含量变化常常伴随其组分的变化。本研究中皂荚苗木蛋白质含量的增加是否与合成新的逆境蛋白质有关,尚需进一步研究。总的来说,因刺槐、皂荚树种的差异,Pro、可溶性糖、可溶性蛋白等参与渗透调节的物质、调节强度存在差别。2种苗木叶片中主要渗透调节物质Pro、可溶性糖的调节能力顺序为刺槐大于皂荚。
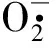
植物在干旱条件下会发生细胞膜过氧化作用而受到损伤,MDA是膜脂过氧化作用的最终产物,其含量的升高,使细胞膜透性增加,影响细胞的正常代谢过程[13]。本研究中,随着土壤含水量的降低,2种苗木叶片的MDA含量持续上升,且重度胁迫时增幅最大,轻度胁迫增幅最小。这可能是由于重度胁迫时SOD活性较低,清除活性氧能力弱,MDA含量升高较多。反之,轻度胁迫时SOD活性大幅度升高,有效清除了细胞中的自由基,使细胞膜脂过氧化程度降低,MDA含量增加较少。SOD活性与MDA含量呈一定负相关性。另外,在相同水分梯度下,皂荚MDA含量升高幅度大于刺槐。说明干旱条件下皂荚细胞膜受损程度大于刺槐,刺槐叶片的抗氧化能力强于皂荚。
本研究对2种苗木的渗透调节物质、保护酶活性进行了初步探讨,仅这些方面来说,刺槐苗木叶片的耐旱性大于皂荚。但是影响植物耐旱性的指标是多方面的,其复杂的机制尚需进一步研究。
[1] 孙映波,尤毅,朱根发,等.干旱胁迫对文心兰抗氧化酶活性和渗透调节物质含量的影响[J].生态环境学报,2011,20(11):1675-1680.
[2] 陈吉虎,余新晓,孙明高,等.北方旱区不同树种抗氧化酶活性变化及与抗旱性的关系[J].干旱地区农业研究,2006,24(5):120-124.
[3] 项忠阳.刺槐和紫穗槐抗旱性比较试验[J].防护林科技,2014(4):25-27.
[4] 徐莲珍,蔡靖,姜在民,等.水分胁迫对3 种苗木叶片渗透调节物质与保护酶活性的影响[J].西北林学院学报,2008,23(2):12-16.
[5] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2001.
[6] 高俊凤.植物生理学试验技术[M].西安:世界图书出版社,2000:137-202.
[7] 西北农业大学植物生理生化教研室.植物生理实验指导[M].西安:陕西科学技术出版社,1987:51-55.
[8] 中国科学院上海植物生理研究所,上海市植物生理学会.现代植物生理学实验指南[M].北京:科学出版社,2004.
[9] 邹原东,韩振芹,陈秀新,等.干旱胁迫对蓝羊草渗透调节物质和抗氧化酶活性的影响[J].北方园艺,2013(23):71-75.
[10] 茹广欣,郝绍菊,茹桃勤,等.干旱梯度下刺槐无性系生理指标的变化与品种抗旱性关系的研究[J].河南科技学院学报(自然科学版),2006,34(1):37-40.
[11] 史玉炜,王燕凌,李文兵,等.水分胁迫对刚毛柽柳可溶性蛋白、可溶性糖和脯氨酸含量变化的影响[J].新疆农业大学学报,2007,30(2):5-8.
[12] 史胜青.四种树种苗木抗旱机制的研究[D].保定:河北农业大学,2003.
[13] 郭智涛.四种刺篱植物的抗旱性研究[D].保定:河北农业大学,2014.
[14] 王海珍,徐雅丽,张翠丽,等.干旱胁迫对胡杨和灰胡杨幼苗渗透调节物质及抗氧化酶活性的影响[J] .干旱区资源与环境,2015,29(12):125-130.
[15] 李燕,孙明高,孔艳菊,等.皂角苗木对干旱胁迫的生理生化反应[J].华南农业大学学报,2006,27(3):66-69.
[16] 李州,彭燕,苏星源,等.不同叶型白三叶抗氧化保护及渗透调节生理对干旱胁迫的响应[J].草业学报,2013,22(2):257-263.
[17] 刘晓东,李洋洋,何淼,等.PEG模拟干旱胁迫对玉带草生理特性的影响[J].草业学报,2012,29(5):687-693.
[18] 包卓,孟祥英,张晓松,等.干旱胁迫对5种园林绿化植物光合速率和渗透调节的影响[J].江苏农业科学,2010(3):225-227.
[19] 孙国荣,张睿,姜丽芬,等.干旱胁迫下白桦实生苗叶片的水分代谢与部分渗透调节物质的变化[J] .植物研究,2001,21(3):413-415.
[20] 李合生.现代植物生理学[M].北京:高等教育出版社,2002:415-419.
Effects of Drought Stress on Osmotic Adjustment Substances Content and Activity of Protect Enzymes in the Leaves ofRobiniapseudoacaciaandGleditsiasinensis
CHEN Lipei,LIU Ruixia,YANG Yuzhen,WANG Guoxia,LUO Hongyu
(School of Life Sciences,Zhengzhou Normal University,Zhengzhou 450044,China)
This paper studied the effects of drought stress on the content of osmotic adjustment substances,MDA and protective enzyme activity in the leaves of one-year-oldGleditsiasinensisandRobiniapseudoacaciathrough the experiment of controlling water by potted planting.The results indicated that with the aggravation of drought stress,the content of Pro and soluble sugar in the leaves of the two seedlings were higher than that of normal water,Robiniapseudoacaciahad greater increasing amplitude under the same water gradient.The content of soluble protein in the leaves ofGleditsiasinensisincreased,but in the leaves ofRobiniapseudoacaciadecreased.The content of MDA increased with the aggravation of water stress,butGleditsiasinensisincreased significantly under the same water gradient.The activity of SOD increased,however the increasing amplitude ofRobiniapseudoacacia>Gleditsiasinensis.POD activity increased and CAT activity decreased in leaves ofGleditsiasinensis,while theRobiniapseudoacacialeaves were just the opposite.The results showed that the two seedlings enhanced the adaptability of drought stress through the accumulation of osmotic adjustment substance and the increase of antioxidant enzyme activity.Overall,the tolerance to drought ofRobiniapseudoacacia>Gleditsiasinensis.
Gleditsiasinensis;Robiniapseudoacacia; droughtstress; osmotic adjustment substances; protective enzyme activity
S792.27
A
1004-3268(2017)10-0122-06
2017-05-20
河南省高等学校重点科研项目计划(16A180054)
陈丽培(1981-),女,河南郑州人,讲师,硕士,主要从事森林培育研究。E-mail:chenlp0524@126.com
*通讯作者:杨玉珍(1965-),女,河南郑州人,教授,博士,主要从事森林培育研究。E-mail:yzhyang@163.com
