基于Meta分析的国内鱼、鳖源嗜水气单胞菌毒力基因研究
2017-10-20牛志伟吕小丽韩书煜黎姗梅梁静真韦慕兰邓小红
牛志伟,吕小丽,黄 钧*,韩书煜,黎姗梅,梁静真,韦慕兰,邓小红
(1. 广西大学动物科学技术学院/广西水生动物病害诊断实验室,广西 南宁 530005;2. 陆川县水产技术推广站,广西 陆川 537700;3. 广西壮族自治区水产技术推广总站,广西 南宁 530021;4. 都安瑶族自治县水产技术推广站,广西 都安 530700;5. 全州县水产技术推广站,广西 全州 541500)
基于Meta分析的国内鱼、鳖源嗜水气单胞菌毒力基因研究
牛志伟1,吕小丽2,黄 钧1*,韩书煜3,黎姗梅3,梁静真1,韦慕兰4,邓小红5
(1. 广西大学动物科学技术学院/广西水生动物病害诊断实验室,广西 南宁 530005;2. 陆川县水产技术推广站,广西 陆川 537700;3. 广西壮族自治区水产技术推广总站,广西 南宁 530021;4. 都安瑶族自治县水产技术推广站,广西 都安 530700;5. 全州县水产技术推广站,广西 全州 541500)
通过借鉴Meta分析法分析中国知网数据库(CNKI)中有关鱼源、鳖源嗜水气单胞菌的7种毒力基因的相关研究数据,探讨鱼源、鳖源嗜水气单胞菌毒力基因的分布特点及为研制嗜水气单胞菌多价DNA疫苗提供候选基因。分析结果表明:嗜水气单胞菌毒力基因的分布受养殖对象的种类和生长环境影响;南方和北方地区鱼源嗜水气单胞菌以毒力基因astA、altA、aerA和hlyA分布较为广泛,南方地区鳖源嗜水气单胞菌毒力基因hlyA、actA和aerA分布均较为广泛,可分别作为制备鱼源和鳖源嗜水气单胞菌多价DNA疫苗候选基因。
嗜水气单胞菌;鱼源;鳖源; 毒力基因;Meta分析;DNA疫苗;
嗜水气单胞菌(Aeromonashydrophila)属于气单胞菌科(Aermonadaceae)气单胞菌属(Aeromonas),是革兰氏阴性短杆菌,广泛分布在各种水体中,容易导致多种水产养殖对象患病,给我国水产养殖业造成巨大的经济损失。有关研究发现嗜水气单胞菌的致病性和携带的毒力基因密切相关:朱大玲等[1]通过比较基因检测的结果与嗜水气单胞菌对鲫鱼的致病性,发现丝氨酸蛋白酶(ahpA)阳性的嗜水气单胞菌皆为毒力菌株;付巧芳[2]通过比较嗜水气单胞菌毒力基因的分布情况及其对异育银鲫的致病性,结果发现气溶素(aerA)、丝氨酸蛋白酶(ahpA)基因为嗜水气单胞菌致病的主要毒力因子。国内研究与致病性相关的毒力基因主要有气溶素(aerA)、溶血素(hlyA)、细胞性肠毒素(actA)、粘附素(ahaI)、热敏感肠毒素(altA)、丝氨酸蛋白酶(ahpA)、金属蛋白酶(astA)7种,有关嗜水气单胞菌携带不同毒力基因对水产动物致病力的影响且将上述基因全部同时检测的研究目前尚未见报道[3]。
DNA疫苗是将编码特异性抗原多肽或蛋白的基因构建在含有调控元件的表达性DNA质粒中,可直接导入机体并在机体内表达目的抗原多肽或蛋白,诱导机体产生针对目的蛋白的免疫应答以达到预防和治疗的目的[4]。其优点是易于构建、稳定性好、便于储存,且能刺激机体产生体液及细胞免疫[5],目前,国内对嗜水气单胞菌DNA疫苗的研究比较少,朱大玲[6]构建了气溶素APT区域的DNA疫苗,但是免疫效果不明显,其他研究集中在鉴定具有免疫原性的蛋白。目前鉴定出具有免疫原性的蛋白基因有气溶素基因[7](aerA)、溶血素基因[7-8](hlyA) 热敏感丝氨酸蛋白酶[8](ahpA)、细胞性毒肠毒素[9](actA)、金属蛋白酶[9](astA)、热敏感肠毒素[9](altA)、粘附素基因[10](ahaI)、胞外蛋白[11](OMP)、鞭毛蛋白FlaB基因[12]等。
通过借鉴Meta分析的方法,搜集前人研究嗜水气单胞菌毒力基因的相关文献,提取并分析数据,以期找到鱼源、鳖源嗜水气单胞菌毒力基因在我国南北地区(以淮河划分)以及不同水产养殖动物之间的分布差异,为研制嗜水气单胞菌DNA疫苗提供参考。
1 资料与方法
1.1 文献的检索
利用中国知网数据库(CNKI)进行水产和渔业科学领域相关文献检索,使用的3种检索策略分别为:①嗜水气单胞菌*(气溶素+溶血素+细胞兴奋性肠毒素+粘附素+细胞性肠毒素+胞外蛋白酶+抗金属蛋白酶);②嗜水气单胞菌*(aerA+hlyA+actA+ahaI+altA+ahpA+astA);③嗜水气单胞菌*(毒力基因+毒力因子)。检索文献发表的时间范围为建库至2016年10月。
1.2 纳入和排除标准
纳入标准:①研究对象为嗜水气单胞菌的毒力基因;②菌株来源地区详细(能区分来自南方或者北方);③菌株源自养殖对象类别详细(能区分来自鱼、鳖、蛙等);④研究菌株样本量≥3株;⑤毒力基因检测方法为PCR子检测;⑥细菌鉴定方法为生化鉴定和分子鉴定结合。
排除标准:①重复发表或者同一作者,同一研究课题组在相同年度发表的相似研究;②地区描述不能区分南北地区;③菌株来自多种养殖对象,但没说明具体来自那种养殖对象;④毒力基因源于不同属的菌株,没有说明毒力基因具体来源菌株;⑤研究菌株样本量<3株;⑥比较不同方法检测毒力基因差异的研究;⑦数据不详的会议文件。
1.3 文献的筛选和提取资料
由2名科研人员根据上述的纳入和排除标准,分别独立进行筛选和提取,在提取过程中遇到争议,则用讨论法决定是否提取数据。提取的主要数据包括:作者、题目、毒力基因名称、地区和检出数量等数据。
1.4 统计分析
使用Excel 2007和SPSS.18等软件进行数据处理和分析,通过卡方分析法分析不同来源(相同地区不同宿主或相同宿主不同地区)菌株毒力基因携带率的差异。
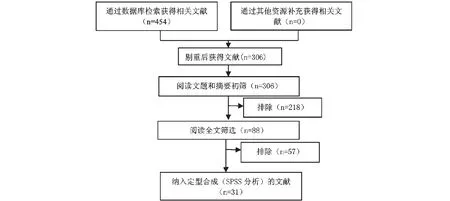
图1 文献筛选流程及结果Fig.1 Screening process and result of literature
2 结果与分析
2.1 纳入文献结果
共检索到454篇文献,排除重复发表或重复下载的文献148篇,阅读题目摘要排除218篇,阅读全文排除文献57篇(图1),最终纳入8个鳖源嗜水气单胞菌毒力基因研究(来源均为南方地区),25个鱼源嗜水气单胞菌毒力基因研究(南方17个,北方8个)共31篇文献33个研究(表1)数据来源见表2。
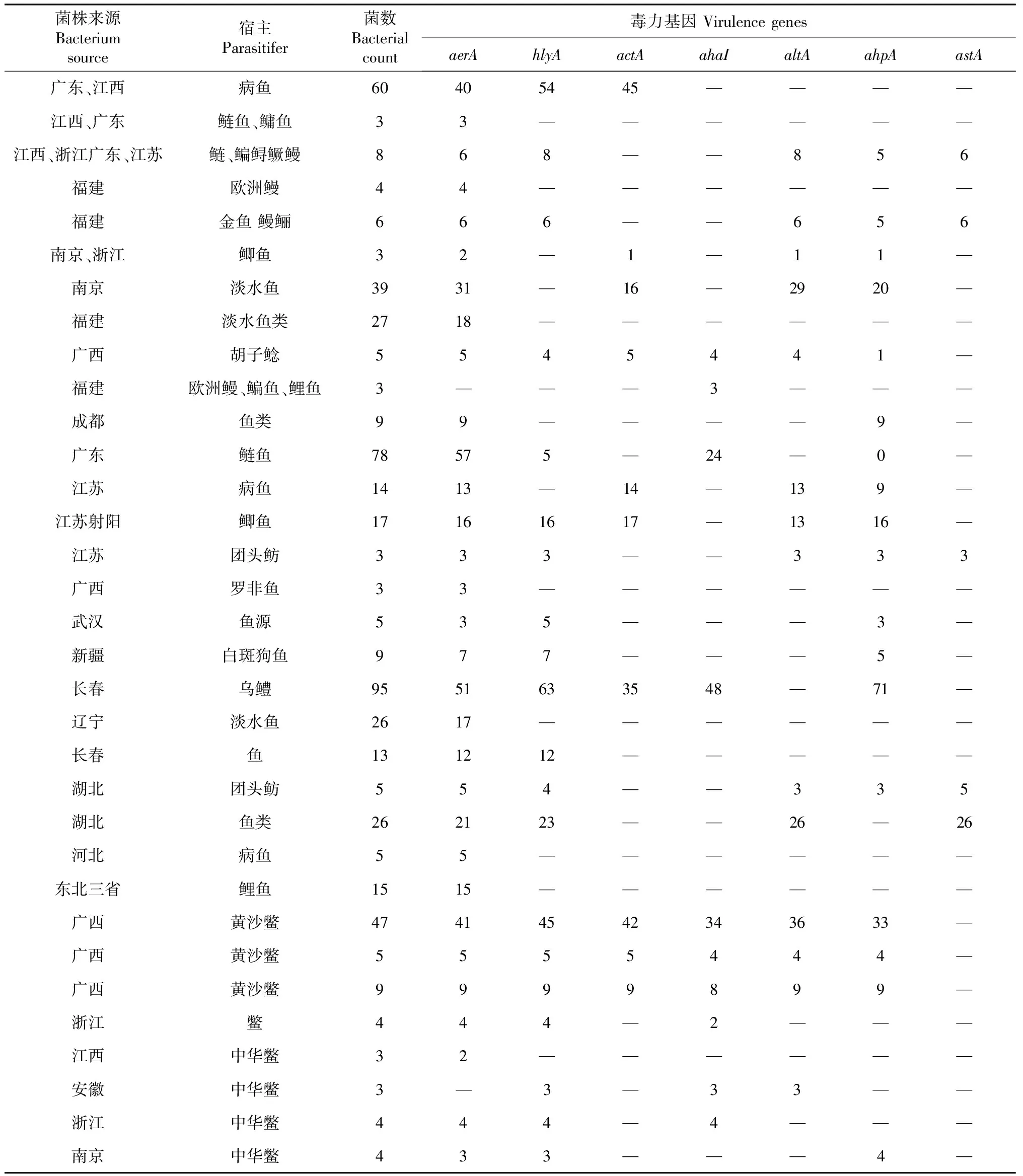
表1 纳入的毒力基因研究文献
注:“—”表示该研究未检测此毒力基因,若菌株来源为作者所在实验室,则视实验室所在地区为菌株来源地区。
Note: ‘—’,indicated that virulence genes was not detected, If the strain from the author's laboratory, depending on the area of author's laboratory as the strain source region.

表2 数据来源
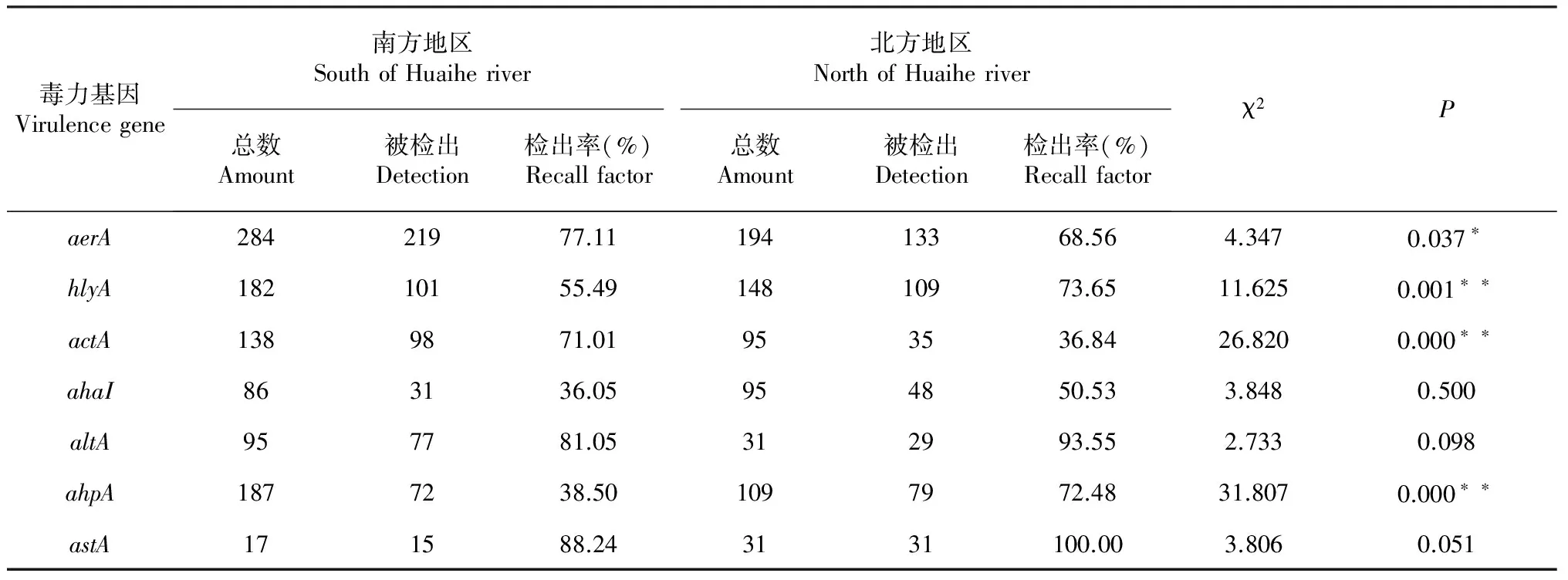
表3 鱼源毒力基因检出情况及卡方分析
注:“*”代表差异显著,”**”代表差异极其显著,P<0.0001时记为0.000,下同。
Note: ‘*’Indicated significant difference, ‘**’ Indicated difference is extremely significant,P<0.0001 denoted asP=0.000, The same as below.

表4 淮河南鱼源和鳖源毒力基因分布情况及卡方分析
2.2 鱼源嗜水气单胞菌毒力基因分布情况
计算纳入研究数据的毒力基因检出率结果(表3)显示,南方地区嗜水气单胞菌7种毒力基因分布以astA为主,检出率为88.24 %;altA次之(81.05 %);ahaI最少(36.05 %);aerA、actA、hlyA和ahpA检出率分别为77.11 %、71.01 %、55.49 %和38.50 %。北方地区也以astA为主,检出率为100 %;altA次之(93.55 %);actA最少(36.84 %);hlyA、ahpA、aerA和ahaI检出率分别为:73.65 %,72.48 %,68.56 %和50.53 %。卡方分析表明,在南方和北方地区之间,毒力基因aerA检出结果差异达显著水平;hlyA、actA和ahpA达极显著水平,ahaI、altA和astA差异不显著。
2.3 鳖源毒力基因分布情况及与鱼源毒力基因检出情况比较分析
从鳖源嗜水气单胞菌毒力基因分布情况可以看出(表4),鳖源嗜水气单胞菌毒力基因以hlyA为主,检出率为96.05 %;actA次之(91.80 %);ahaI最低(76.39 %);aerA、altA和aphA检出率分别为89.47 %,81.25 %和76.92 %。南方鳖源和鱼源之间,嗜水气单胞菌毒力基因aerA检出结果差异达显著水平、hlyA、actA、ahaI和ahpA检出结果差异均达极显著水平,altA检出结果差异不显著。
3 讨 论
不同的毒力基因具有不同的主要功能,动物被大量致病性嗜水气单胞菌感染时,肠内肠毒素含量急速上升,且与细胞发生不可逆的结合,使体液严重损失,最终导致宿主死亡[13]。抗金属蛋白酶可直接破坏宿主组织获取营养导致致宿主死亡[14]。细菌病原的感染是一个复杂的、动态的多因子作用过程,它要求病原菌众多毒力基因产物的协调一致,并能与宿主的免疫系统一同进化,在大多数情况下,病原菌会根据感染部位所遇到的环境随时调整各毒力基因的表达[15]。
疫情发生与宿主动物带菌率高且菌株毒力基因阳性率高有密切关系[16]。本研究对在前人的研究数据进行统计分析发现,不同地区致病鱼源嗜水气单胞菌毒力基因检出率较高的毒力基因是altA和astA,并且差异不显著,说明altA和astA可能是鱼源嗜水气单胞菌最基本的毒力基因或者大多嗜水气单胞菌在致病过程中依靠astA获取营养,且侵害部位主要是肠道。其余4种毒力基因aerA、actA、hlyA、ahpA的检出率在不同地区分布差异显著,说明这4种毒力基因分布或在不同菌体之间传播受环境影响较大。通过比较分析相同地区鱼源和鳖源嗜水气单胞菌毒力基因分布情况,发现相同地区不同来源嗜水气单胞菌毒力基因分布大多不同,且差异显著,说明嗜水气单胞菌针对不同养殖对象选择的入侵对策不同,龟源嗜水气单胞菌毒力基因以hlyA和actA为主和鱼源的不同,可能针对宿主本身所含的营养物质或防御措施不同,细菌选择的分解方式和入侵策略不同所致。侧面反映出嗜水气单胞菌侵袭鳖的症状以溶血和肠炎为主[17],剩余五种毒力基因aerA、hlyA、actA、ahaI、ahpA检出率鳖源均高于鱼源且差异显著,说明嗜水气单胞菌入侵鳖需要更多的毒力基因协作或者鳖的防御能力高于鱼。
目前正在研究的疫苗包括:全菌灭活苗、减毒活疫苗。因为嗜水气单胞菌血清型众多,不同地区、不同鱼种之间流行的菌株差异明显等原因,致使嗜水气单胞菌疫苗不能商品化而不能普及[18],传统的亚单位疫苗的最大局限性是所用的抗原不能在宿主细胞中表达,因而不能诱导细胞免疫,DNA疫苗兼有重组亚单位疫苗的安全性和减毒活疫苗的全面免疫应答的高效力[19],如果可以将分布广泛的几种毒力基因的功能区域,并利用基因工程技术将几种毒力基因的功能区域合在一起制备DNA疫苗,对于解决嗜水气单胞菌血清型众多,疫苗难以商品化的难题,是最理想的方法[20]。通过统计分析发现,致病性鱼源嗜水气单胞菌分布比较广泛的毒力基因是altA、astA、aerA、hlyA,鳖源嗜水气单胞菌分布比较广泛的毒力基因是hlyA、actA、aerA,若能找到这5种毒力基因的功能区域,并分别制备成多价DNA疫苗,对攻克鱼源、鳖源嗜水气单胞菌及普及疫苗具有一定意义。
由于本研究是在前人研究基础上进行的二次研究,对于前人研究的毒力基因是随机检测还是存在人为选择难以把握,计算出的检出率可能偏高,所以只做定型分析,通过整合前人的研究数据也增大了菌株样本含量,具有一定的统计学意义。
4 结 论
嗜水气单胞菌对养殖对象的致病机理随环境和养殖对象种类的影响而不同,鱼源致病性嗜水气单胞菌毒力基因altA、astA、aerA、hlyA分布广泛,鳖源致病性嗜水气单胞菌毒力基因hlyA、actA、aerA分布比较广泛,可分别作为制备鱼源和鳖源嗜水气单胞菌多价DNA疫苗候选基因。
[1]朱大玲,李爱华,汪建国,等. 嗜水气单胞菌毒力与毒力基因分布的相关性[J]. 中山大学学报(自然科学版),2006,45(1):82-85.
[2]付乔芳. 嗜水气单胞菌毒力特性研究[D]. 上海:上海海洋大学. 2011.
[3]牛志伟,吕小丽,范华龙,等.中国耐喹诺酮类嗜水气单胞菌毒力基因及耐药基因检出情况的系统评价[J].南方农业学报,2016,47(9):1587-1595.
[4]聂建辉,王佑春. DNA疫苗的研究现状[J]. 中国药事,2007,21(2):114-118.
[5]谭 伟,谢丽基,谢芝勋. 禽流感病毒疫苗研究进展[J]. 南方农业学报,2014,45(8):1492-1497.
[6]朱大玲. 嗜水气单胞菌毒力基因及基因工程疫苗[D]. 北京:中国科学院研究生院(水生生物研究所),2006.
[7]刘明智,叶 星,田园园,等. 嗜水气单胞菌气溶素和溶血素基因的克隆与结构预测[J]. 江西农业大学学报,2010,32(2):356-362.
[8]潘晓艺,郝贵杰,姚嘉赟,等. 嗜水气单胞菌TPS-30株丝氨酸蛋白酶基因与溶血素基因在大肠杆菌中的融合表达[J]. 水生生物学报,2010,34(3):591-597.
[9]倪晓丹. 嗜水气单胞菌临床分离株主要毒素基因型分析及胞外蛋白免疫蛋白组学研究[D]. 江苏省南京市:南京农业大学,2008.
[10]庄培德. 嗜水气单胞菌主要粘附素基因克隆表达及其产物特性分析[D]. 福建省福州市:福建农林大学,2007.
[11]蒋 蔚. 嗜水气单胞菌J-1株OmpA蛋白原核表达、免疫原性分析及适于四膜虫的OmpA表达载体构建[D].江苏省南京市:南京农业大学,2007.
[12]沈雪飞. 嗜水气单胞菌鞭毛蛋白的纯化及重组蛋白FlaB多抗的制备[D].吉林省长春市:吉林大学,2014.
[13]方一风,潘晓艺,蔺凌云,等. 嗜水气单胞菌对喹诺酮类药物耐药的分子机制[J]. 微生物学报,2014,54(2):174-182.
[14]李槿年,余为一,魏梅芳. 36株气单胞菌外毒素溶血性和致病性的测定[J]. 中国预防兽医学报,2000,22(2):51-53.
[15]陈师勇,莫照兰,张振冬,等. 细菌毒力基因体内表达检测技术研究进展[J]. 遗传,2005,27(3):505-511.
[16]王 耀. 肉制品和乳制品中致病菌检测技术体系建立及李氏菌分型鉴定与溯源研究[D].沈阳:沈阳农业大学,2011.
[17]赵蓓蓓. 鳖源嗜水气单胞菌ZHYYZ-1表型、致病机制及抑菌中草药复方筛选的研究[D]. 舟山:浙江海洋学院,2012.
[18]龚新勇,欧阳岁东,主 性. 鱼嗜水气单胞菌疫苗研究进展[J]. 贵州畜牧兽医,2004,28(3):19-20.
[19]李 萍, 严家新. DNA疫苗研究进展[J]. 国际生物制品学杂志, 1995(3):100-104.
[20]朱大玲,李爱华,钱 冬,等. 嗜水气单胞菌毒力基因的研究进展[J]. 水生生物学报,2004(1):80-84.
(责任编辑 汪羽宁)
StudyonVirulenceGenesofAeromonashydrophilafromFishandTurtleinChinaBasedonMetaAnalysis
NIU Zhi-wei1,LV Xiao-li2,HUANG Jun1*,HAN Shu-yu3,LI Shan-mei3,LIANG Jing-zhen1,WEI Mu-lan4,DENG Xiao-hong1
(1.Institute of Animal Science and Technology of Guangxi University/ Guangxi Aquatic Animal Disease Diagnostic Laboratory, Guangxi Nanning 530005, China; 2.Guangxi Luchuan Aquaculture Technology Extension Station, Guangxi Luchuan 537700, China; 3.Guangxi Zhuang Autonomous Region, Aquaculture Technology Extension Station, Guangxi Nanning 530005, China; 4.Guangxi Duan Yao Autonomous County Aquaculture Technology Extension Station, Guangxi Du’an 530700, China; 5.Guangxi Quanzhou Aquaculture Technology Extension Station, Guangxi Quanzhou 541500, China)
The data of 7 kinds of virulence genes ofAeromonashydrophilafrom fish and turtle in CNKI database were analyzed based on Meta-analysis, and the distribution characteristics of virulence genes ofAeromonashydrophilafrom fish and turtle were discussed to provide candidate gene for generate multivalent DNA vaccine ofAeromonashydrophila. The results showed that the distribution of virulence genes fromAeromonashydrophilawas affected by the types of cultured species and growth environment. The virulence genes ofastA,altA,aerAandhlyAfromAeromonashydrophilaof fish were widely distributed in the north and south of China. The virulence genes ofhlyA,actA, andaerAfrom turtle sourceAeromonashydrophilawere widely distributed in the south of China, which could be used as candidate genes for generating multivalent DNA vaccine ofAeromonashydrophilain fish and turtle respectively.
Aeromonashydrophila; Fish derived; Turtle derived; Virulence genes; Meta analysis; DNA vaccine
1001-4829(2017)3-0711-06
10.16213/j.cnki.scjas.2017.3.041
S941.42+9
A
2016-11-08
广西水产畜牧兽医局专项项目(桂渔牧财[2014]52号,桂渔牧财[2015]97号)
牛志伟(1990-),男,河北邢台人,硕士研究生,研究方向水生动物病害防控及健康养殖,E-mail:905478661@qq.com,*为通讯作者,E-mail:hj1351@163.com
