热带、亚热带玉米自交系与温带玉米自交系的配合力和遗传参数分析
2017-10-20段智利杨晓婷陈洪梅希从芳汪燕芬李建宾
段智利,杨晓婷 *,陈洪梅,希从芳,汪燕芬,王 晶,李建宾 **
(1.云南农业大学农学与生物技术学院,云南 昆明 650201;2.云南省农业科学院粮食作物研究所,云南 昆明 650205)
热带、亚热带玉米自交系与温带玉米自交系的配合力和遗传参数分析
段智利1,杨晓婷1 *,陈洪梅2 **,希从芳1,汪燕芬2,王 晶1,李建宾1 **
(1.云南农业大学农学与生物技术学院,云南 昆明 650201;2.云南省农业科学院粮食作物研究所,云南 昆明 650205)
以4个优良热带、亚热带玉米自交系为测验种,9个优良温带玉米自交系为被测系,按NCⅡ遗传交配设计配制36个杂交组合,在云南省昆明、曲靖和德宏3个不同生态条件下进行田间鉴评,研究其9个性状的配合力及遗传参数。结果表明:热带自交系CRN3760-1-2-2-1-1-2的产量GCA效应值最高,其次是YML46、[阿1112×T(5003×云146)]##-1-1-1-1-B、2870263-4-2-1-B-B-B-B-B、IRF321、8723、23X等自交系,都具有显著的产量GCA正效应值,是组配强优势杂交组合的优良自交系。杂交组合CRN3760-1-2-2-1-1-2×23X、CRN3760-1-2-2-1-1-2×2870263-4-2-1-B-B-B-B-B、CRN3760-1-2-2-1-1-2 ×[阿1112×T(5003×云146)]##-1-1-1-1-B和YML46 × 8723具有显著的产量SCA效应值,有一定的增产潜力,可作为潜势组合在云南进一步鉴定其丰产性和稳定性。9个性状的遗传以加性基因效应为主,轴粗、百粒重和出籽率的狭义遗传力都大于55.2 %,其变异由遗传引起的比重较大,宜在早代进行选择,穗粗、秃尖和穗行数的狭义遗传力低于23.2 %,宜在晚代选择。
热带、亚热带玉米;种质;配合力;杂种优势;遗传参数
玉米种质资源狭窄已经成为世界玉米育种可持续发展的主要限制因素[1]。目前中国推广的杂交种中,80 %以上的亲本集中在Lancaster、Reid、唐四平头和旅大红骨等4大杂种优势群内,种质基础狭窄是中国玉米育种可持续发展的“瓶颈”[2-4]。
研究表明,温带和暖温带玉米生长期短、植株矮、雄穗小、经济系数高,但生产力受“源”的影响大,热带和亚热带玉米具有大量的有利遗传变异,生长期长,生物量高,籽粒脱水快,根系发达,茎秆强韧,有温带种质不具备的抗旱、抗病虫害、耐瘠、耐阴湿等特点,是温带玉米育种不可多得的异源种质[2-6]。但是,热带和亚热带玉米种质在温带种植存在强烈的温光反应,限制了其在育种中的直接利用[2,7]。为了解决这一难题,20世纪50年代以来,许多育种家不断探索热带、亚热带玉米种质资源利用的途径和方法,通过“逐步驯化”、“逐步导入”等选育优良自交系S37、CML140、CML94、B14、B37、B117、B121、YA3237、YA8201、GP2-1、GP5-2等[5,8-12]。玉米杂种优势主要取决于亲本系的GCA改良和系间SCA提高。为了获得最佳效果,必须对外来热带、亚热带玉米种子进行杂种优势群的准确划分,探明群体育种潜势,采取相应的育种策略和方法。根据Vasal等[13]、番兴明等[14]、陈洪梅等[15]和苏俊等[16]研究结果,热带、亚热带玉米群体Suwan1、CIMMYT、B110等和一些温带玉米自交系具有较高的配合力,为改良和利用热带、亚热带玉米种质奠定了较好的基础。
本研究以4个热带、亚热带玉米自交系和9个温带玉米自交系按NCⅡ交配设计进行杂交,并分析了其F1的产量、穗长、穗粗、百粒重等9个农艺性状及配合力、杂种优势,对深入利用热带、亚热带玉米种质,为云南省发掘新的玉米杂种优势模式,提供新的高产玉米杂交组合具有重要的指导意义。
1 材料与方法
1.1 试验材料与设计
选用的9个自交系BY815-1、吉853、ANL6、(自336×云146)-2-2-1-2-2-B、8723、IRF32、23X、2870263-4-2-1-B-B-B-B-B和[阿1112×T(5003×云146)]##-1-1-1-1-B为温带玉米优势群,4个测验种YML46、CRN3760-1-2-2-1-1-2、(素1×吉840)-2-2-2-1-1-1、 ACROSS856为热带、亚热带玉米自交系,其系谱来源见表1。
按不完全双列杂交(NCⅡ)交配设计,以4个热带、亚热带玉米自交系为母本、9个温带玉米自交系为父本进行杂交,配制36个杂交组合。
1.2 田间试验设计
36个杂交组合分别在昆明(海拔1960 m,120°45′ E,25°02′ N)、曲靖(海拔1862.5 m,103°40′ E,25°30′ N)和德宏(海拔913.8 m,98°35′ E,24°26′ N)等3个不同生态条件下进行田间鉴评。玉米田间栽培管理与当地大田一致。
采用随机区组设计,3次重复,1行区,行长5 m,小区面积3 m2,等行距种植,行距0.6 m,株距0.25 m,种植密度66 667株/hm2;试验设1个统一对照(CK)海禾2号。
1.3 调查项目
分别在开花授粉期和成熟期进行调查和取样。每小区连续选取有代表性的玉米5株(不取边株),成熟后收获果穗,进行室内穗部性状的调查,收集穗长(cm)、穗粗(cm)、轴粗(cm)、秃尖(cm)、穗行数(行)、行粒数(粒)、百粒重(g)、出籽率( %)、单株产量(g/株)等9个性状的数据。

表1 供试自交系的种质来源

表2 36个玉米杂交组合9个性状的方差分析
注:* 表示在0.05水平差异显著,**表示在0.01水平差异显著。下同。
Note: * Significant atP≤0.05, ** Significant atP≤0.01. The same as below.
1.4 统计分析
3个试验点玉米9个性状调查结果平均后,对平均单株产量数据进行方差分析,只有当差异检验为显著时,才进一步进行配合力分析。采用NCII交配设计分析方法进行配合力方差分析[17-20],并进一步估算一般配合力(GCA)和特殊配合力(SCA)的相对效应值。方差分析、配合力效应分析和采用对照优势对杂交组合进行杂种优势分析。
对照优势(HI):
HI(%)=(F1-X)/X×100
其中,F1为杂交组合单株产量,X为统一对照种海禾2号的单株产量,其他用SAS统计软件和Excel软件完成。
2 结果与分析
2.1 方差分析
9个性状的方差分析结果(表2)看出,被测系和测验种的9个性状差异均达极显著水平,说明所测各个性状在测验种和被测系亲本间存在极显著差异,因而可进一步估算亲本的一般配合力(GCA)效应和组合双亲的特殊配合力(SCA)效应;测验种×被测系间,除了穗粗达到显著水平外,其他性状均达到极显著水平,表明被测系与测验种间这些性状的特殊配合力存在差异。
2.2 杂交组合单株产量与杂种优势分析
根据1年3个试验点产量对照优势(表3)分析,19.4 %的组合对照优势大于3.2 %,其中HI大于10 %的组合有4个,分别为T2× Q7、T2× Q8、T2 ×Q9和T1 ×Q5,这4个组合的单株产量和HI依次是218.133 g和19.199 %、214.067 g和16.976 %、205.967 g和12.550 %以及202.033 g和10.401 %;其余组合没有对照优势。HI小于-20 %的组合有4个,分别为T4 × Q2、T4× Q1、T4 × Q6以及T4×Q4,这4个组合的平均产量及HI依次是91.800 g和-49.836 %、114.300 g和-37.541 %、134.867 g和-26.302 %以及135.200 g和-26.120 %。

表3 36个玉米杂交组合的产量及产量对照优势
2.3 一般配合力效应分析
一般配合力(GCA)效应是由亲本基因型的加性效应基因所决定的,是可以遗传的部分,一般配合力高的相应性状的遗传力也高[17,20-21]。
13个自交系9个性状的一般配合力效应值见表4。热带玉米测验种T2和T1以及温带玉米被测系Q9、Q8、Q6、Q5和Q7具有较高的单株产量GCA效应值,说明这些自交系含有较多与产量相关的有利基因,属于优良的高产玉米育种材料,可在今后的育种工作中加以利用。尤其是T2的产量GCA效应值最高,且属于热带材料,在中国温带玉米地区将具有较高的利用潜力。
自交系T1、T2和Q5、Q1、Q3、Q9、Q8等的穗长GCA效应值较高,对于改良玉米果穗的长度,提高产量具有重要意义。穗粗GCA效应值较高的自交系有T1、T3、T2和Q2、Q1、Q7,可作为培育粗穗型玉米的种质加以利用。果穗秃尖GCA较低的自交系有T1、Q8、Q2、Q9、Q7和Q4,这些自交系一般能减少F1果穗的秃尖,提高F1杂种果穗的结实性。穗行数GCA效应值较高的自交系有T1、T3和Q6、Q3和Q9,可以用这些自交系改良穗行数较少的优良玉米自交系。行粒数GCA效应值较高的自交系有T2、T1和Q9、Q1、Q6、Q5,这几个自交系能增加F1
杂交种的行粒数,从而有效增加F1产量。百粒重GCA效应值较高的自交系有T2和Q8、Q2、Q7、Q5、Q4,这些自交系能增加F1的千粒重。其中热带玉米测验种T2的百粒重GCA效应值高达6.110,可作为培育大籽粒玉米种质加以利用。出籽率GCA较高的自交系有T2、T3和Q7、Q8、Q9、Q5、Q6,这几个自交系能增加F1杂交种的出籽率,其中热带玉米测验种T2的出籽率GCA效应值高达2.879,可作为培育较高出籽率的玉米种质加以利用。
2.4 特殊配合力效应分析
特殊配合力(SCA)的高低决定于亲本基因型的非加性基因效应,不能在上下代中稳定遗传,但它可以指导杂种优势的利用和杂交种的选育[22]。本研究13个自交系9个性状的特殊配合力效应值见表5。从表5可以看出,不同组合同一性状的SCA相对效应值有很大的差异。
36个组合中,产量SCA相对效应值(Sij)为正的组合19个,占组合总数的52.8 %,其中产量SCA相对效应值达极显著水平的组合有8个,依次为T2×Q7(Sij= 25.212**)、T3×Q6(Sij= 23.435**)、T4×Q9(Sij= 19.692**)、T1×Q5(Sij= 19.461**)、T3×Q4(Sij= 17.243**)、T2×Q8(Sij= 16.519**)、T4×Q3(Sij= 16.234**)和T1×Q1穗长和行粒数SCA效应值为正的组合各有18个,其中达到显著水平以上的组合分别有8个和7个,组合T1×Q3的穗长和行粒数的SCA效应都达显著水平;秃尖SCA效应为负的组合有18个,其中达显著水平以上的组合有7个,T1×Q5、T4×Q4和T4×Q9的秃尖和行粒数SCA效应都达显著水平;穗粗和穗行数SCA效应值为正的组合分别有18个和19个,但仅有1个组合的穗行数SCA效应值达到显著水平以上,所有组合穗粗的SCA效应值均未达显著水平;穗轴粗SCA为负效应的组合有20个,达到显著水平以上的组合有6个;百粒重SCA效应值为正的组合有19个,达到显著水平以上的组合有7个,其中效应值最大的组合为T1×Q2,其次是T3×Q4和T4×Q8;出籽率SCA效应值最大的组合为T2×Q8,其次是T1×Q1和T3×Q9。

表4 9个性状的一般配合力(GCA)效应值
(Sij= 15.845**),产量显著高于海禾2号的组合有4个,依次是T3×Q2(Sij= 12.551*)、T2×Q1(Sij= 11.096*)、T1×Q6(Sij= 9.961*)和T4×Q8(Sij= 9.749*),说明这12个组合对应的自交系之间具有较强的杂种优势,这些组合可作为潜势组合进一步鉴定其丰产性、稳定性和适应性。
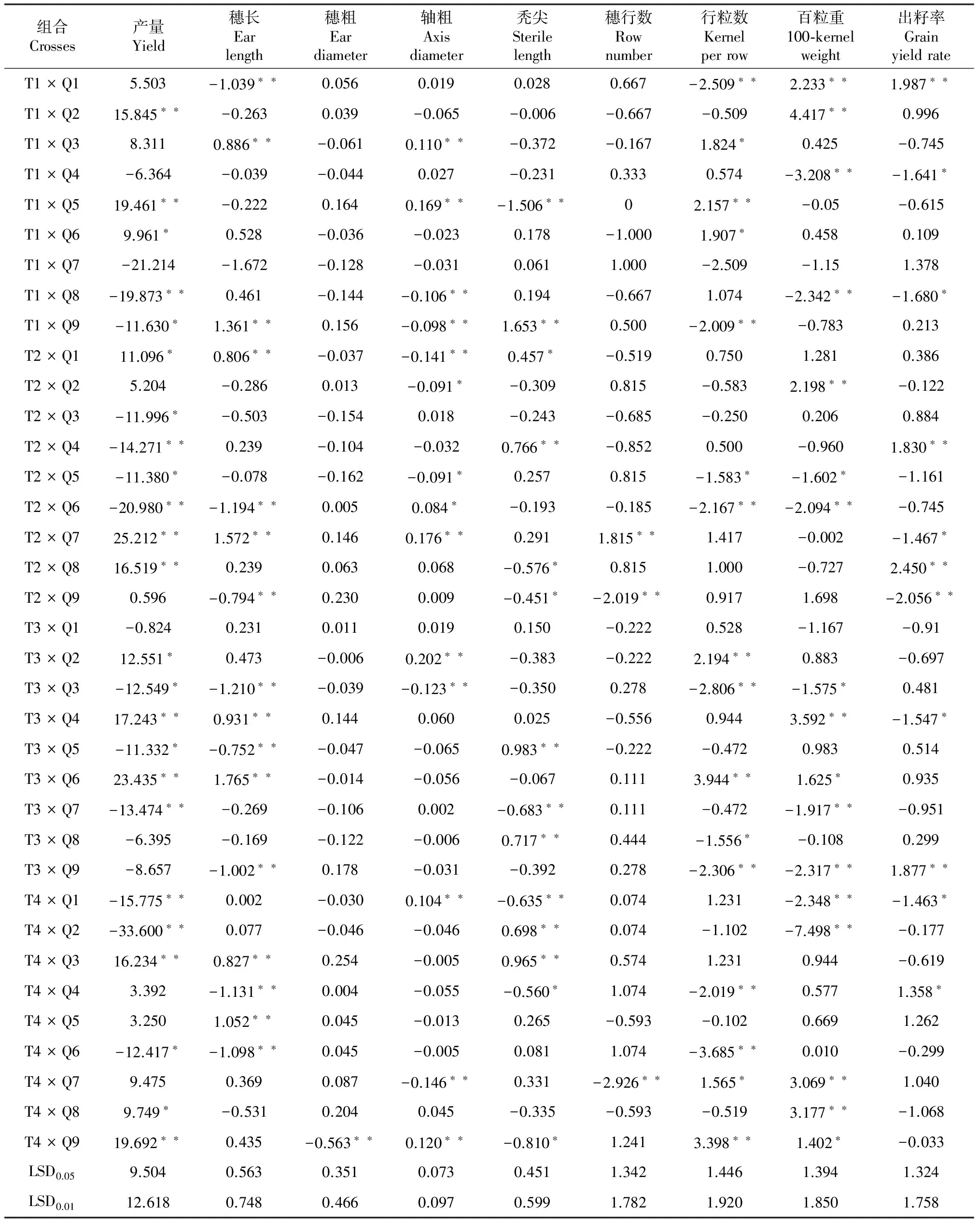
表5 特殊配合力相对效应值
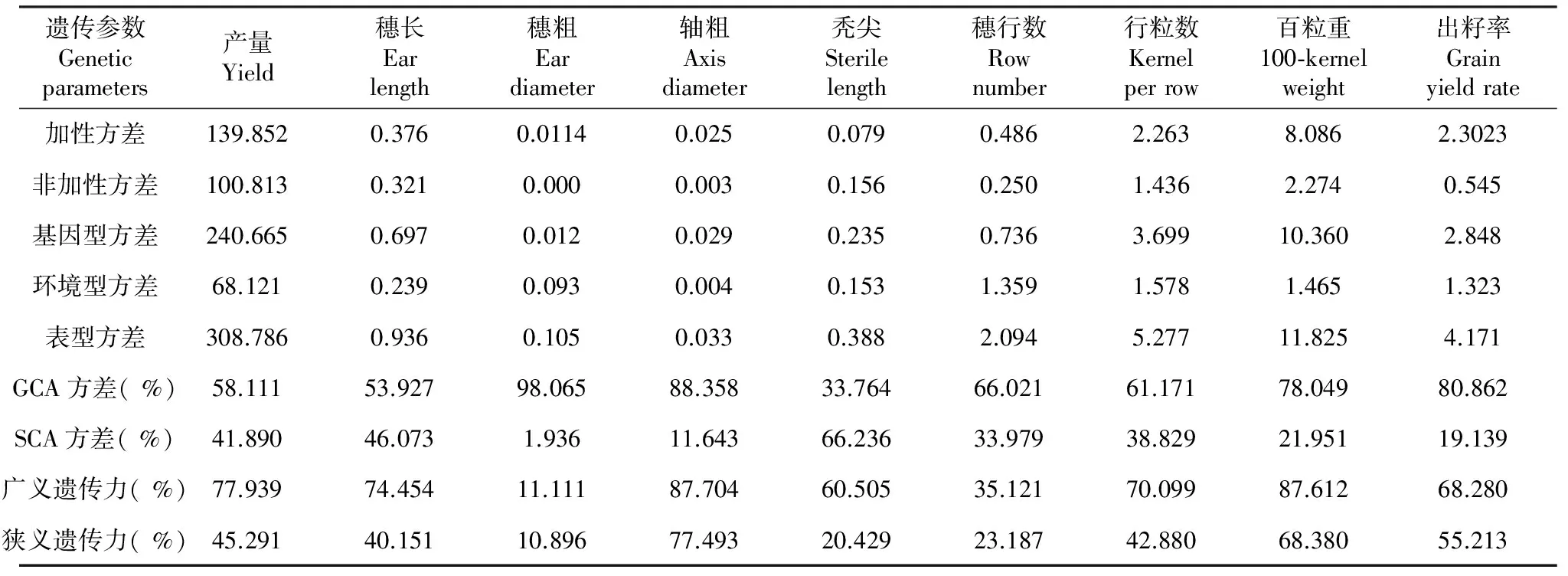
表6 各性状遗传参数估计值
从表5还可看出,百粒重SCA正效应(或负效应)值达到显著水平以上的组合,其产量效应值都为正值(或负值),且有62.5 %~77.8 %的组合产量SCA效应值达显著水平以上,说明组合百粒重SCA效应对其产量SCA效应起到决定性作用。
2.5 性状遗传参数分析
从表6可以看出,同一性状的遗传参数值差异较大。除了秃尖的GCA方差小于SCA方差、加性方差小于非加性方差外,其余8个性状的GCA方差都大于SCA方差、加性方差也大于非加性方差,说明除了秃尖主要受非加性基因的控制外,其余产量、百粒重等8个性状则主要受加性基因控制。
穗粗、轴粗、百粒重和出籽率等性状的GCA方差值是SCA方差值的5.5~104.7倍,说明其基本受加性基因控制,穗长、产量、行粒数和穗行数等性状的GCA方差是SCA方差的1.17~1.94倍,说明其主要受加性基因的控制,也受非加性基因的影响。可见,在组合选配上,应重视选择双亲皆表现优良的亲本,才能充分发挥优良基因的加性效应;在杂交组合选配过程中,穗粗、轴粗、百粒重和出籽率应着重双亲的选择;产量、穗长和穗行数应兼顾双亲和杂交组合的表现;秃尖则可放宽对双亲的选择,着重对杂交组合的表现进行评价。
从表6还可看出,9个性状的广义遗传力都比狭义遗传力高,且基因型方差大于环境型方差;各性状的广义遗传力大小依次为:轴粗﹥百粒重﹥产量﹥穗长﹥行粒数﹥出籽率﹥秃尖﹥穗行数﹥穗粗。此结果说明,上述9个性状的表型主要受遗传基因控制,但无法区分是加性基因还是非加性基因,需要进一步分析其狭义遗传力。
狭义遗传力反应的是该性状受加性基因控制的遗传表现,对育种更有指导意义。本试验研究的9个性状中,轴粗的狭义遗传力最高,其次是百粒重和出籽率,其狭义遗传力都大于55.2 %,说明这几个性状不易受环境条件影响,其变异由遗传引起的比重较大,易在早代对其进行选择;而穗粗、秃尖和穗行数的狭义遗传力小于23 %,说明其主要受非加性基因控制,易受环境影响,应放到晚代进行选择;产量、行粒数和穗长的狭义遗传力在40 %~45 %,在受到加性基因控制的同时,还受环境条件的影响,不宜过早或过晚选择。
3 讨 论
3.1 热带、亚热带玉米自交系的应用潜力
利用热带、亚热带种质资源可以拓宽温带玉米种质的遗传基础,构建新的杂种优势模式,提高品种的生态适应性,创制出强优势的杂交玉米新组合[1-5]。Lewis等[23]、苏俊等[16]以温带系 × 热带系合成杂交种,再用系谱法选系,获得含热带种质的优良新自交系。李芦江等[12]、番兴明等[5]认为,在利用人工合成改良群体选系前,应对群体的育种潜势进行评价,并根据群体遗传特性和自交后代配合力表现,采取相应的策略和方法。一般说来,玉米穗部性状和产量的GCA效应值越大,越有利于改善杂交组合的穗部性状和产量,越容易组配出高产杂交组合[20,24]。本试验结果,4个热带,亚热带玉米自交系的产量GCA效应值为CRN3760-1-2-2-1-1-2 > YML46 > (素1×吉840)222111 > ACROSS8569,其中CRN3760-1-2-2-1-1-2和YML46的产量GCA效应值为极显著正值,而ACROSS8569的产量GCA效应值为极显著负值,说明CRN3760-1-2-2-1-1-2和YML46作亲本,容易组配出产量较高的组合,在生产上有较高的利用价值。
本研究在云南省的昆明、曲靖和德宏等3个生态条件下完成,综合3个试验点的结果,热带、亚热带玉米自交系能适应云南省的生态环境,温带玉米自交系与其杂交后大部分产量表现较好,其中,12个组合CRN3760-1-2-2-1-1-2 × 23X、(素1×吉840)222111 × IRF321、ACROSS8569 ×[阿1112×T(5003×云146)]##-1-1-1-1-B、YML46 × 8723、(素1×吉840)222111×(自336×云146)22122-B、CRN3760-1-2-2-1-1-2×2870263-4-2-1-B-B-B-B-B、ACROSS8569 × ANL6、YML46 × 吉853、(素1×吉840)222111 × 吉853、CRN3760-1-2-2-1-1-2×BY815-1、YML46×IRF321、ACROSS8569 × 2870263-4-2-1-B-B-B-B-B的SCA效应值达到显著水平以上,具有强的杂种优势,可作为潜势组合进一步研究其丰产性、稳定性和适应性。
3.2 温带种质改良系的利用
Holland和Goodman[25]的研究结果表明,改良过的热带种质与温带种质杂交可能出现很高的配合力。荣廷昭等[2]、番兴明等[14]、陈洪梅等[15]认为以热带、亚热带玉米种质为供体回交改良温带玉米自交系是利用热带、亚热带玉米种质的有效方法。本试验中4个热带、亚热带测验种各自具有不同的优良性状,可以用来改良温带玉米自交系。CRN3760-1-2-2-1-1-2的产量和百粒重GCA效应值最高,与温带自交系23X组配的产量SCA效应值最高,可以用其人工合成群体,从中选育出优良自交系,还可以用此自交系改良温带玉米的百粒重。YML46有较高的产量GCA效应值和最高的穗行数效应值,可以用来改良温带玉米种质的穗行数和产量;ACROSS8569的轴粗GCA效应值为极显著正值,可以用其来改良温带种质的轴粗;(素1×吉840)222111的轴粗和出籽率GCA效应值为极显著正值,可以用其来改良温带种质的轴粗和出籽率。
3.3 主要农艺性状的遗传参数
在本研究中,轴粗、百粒重、出籽率的广义和狭义遗传力都高,这些性状低世代可以选择优良个体,也较为可靠,通过重组育种可选育出农艺性状好的品种;而对狭义遗传力较低的秃尖、穗粗和穗行数易受环境影响,不易在早期表现出来,应放到晚代进行选择。这一结果与张亚勤等[20]和李勇成等[26]试验结果一致。
[1]Hoisington D, Khairallah M, Reeves T, et al. Plant genetic resources: what can they contribute toward increased crop productivity[J].Proceedings of the National Academy of Sciences of USA,1999,96(11):5937-5943.
[2]荣廷昭,潘光堂,黄玉碧,等.热带玉米种质在温带玉米育种的应用[J].作物杂志,1998(S):12-14.
[3]张世煌,彭泽斌,李新海.玉米杂种优势与种质扩增、改良和创新[J].中国农业科学,2000,33(S):34-39.
[4]荣廷昭,李晚忱,潘光堂.新世纪初发展我国玉米遗传育种科学技术的思考[J].玉米科学,2003(专刊):42-53.
[5]李 雁,肖植文,伏成秀,等. 云南 28 个玉米杂交组合主成分分析及综合评价[J].西南农业学报,2015,28(1):34-40.
[6]张建宏,郑宏鹏,冯丽赟,等.玉米育种问题及对策探讨[J].山西农业科学,2014,42(7):768-770.
[7]王翠玲,程芳芳,孙朝晖,等.玉米光周期敏感性的遗传特性及相关基因的研究进展[J].玉米科学,2008,16(1):11-14,19.
[8]倪昔玉,刘礼超,雷本鸣.山区玉米育种用优良自交系苏37(S37)的选育研究[J].四川农业大学学报,1996,14(3):366-370.
[9]Hallauer A R, Lamkey K R, White P R. Registration of B117,B118,B119,B120, and B121[J].Crop Sci,2004,44:2280-2281.
[10]胡学爱,杨 荣,刘世建,等.温-热带种质玉米自交系YA3237 和YA3729 选育与应用[J].玉米科学,2011,19(2):42-44,49.
[11]胡学爱,刘世建,杨 荣,等.温带玉米育种利用热带种质的新方法[J].中国农技推广,2013,29(SI):80-84.
[12]李芦江,杨克诚,潘光堂,等.玉米人工合成群体的改良与利用[J].玉米科学,2011,19(1):1-7.
[13]Vasal S K, Srinivasan G, Gonzalez F C, et al. Heterosis and combinging ability of CIMMYT`s tropical × subtropical maize germplasm[J].Crop Sci,1992,32:1483-1489.
[14]番兴明,谭 静,杨峻芸,等.外来热带、亚热带玉米自交系与温带玉米自交系产量配合力分析及其遗传关系的研究[J].中国农业科学,2002,35(7):743-749.
[15]陈洪梅,汪燕芬,姚文华,等.导入热带种质的温带玉米自交系的利用潜力[J].作物学报,2011,37(10):1785-1793.
[16]苏 俊,李春霞,龚士琛,等.热带、亚热带玉米种质在北方早熟春玉米育种中的利用研究[J].玉米科学,2010,18(4):1-6,12.
[17]郭还威,王黄英.玉米不完全双列杂交测定自交配合力的研究[J].玉米科学,1993(4):17-18.
[18]祁志云,杨 华,李淑君,等.玉米种质杂种优势群划分及注意性状相关性与通径分析[J].西南农业学报,2013,26(4):1316-1321.
[19]苟才明,黄 宁,徐克成,等.17个玉米新选系的杂优类群及主要性状相关与通径分析[J].西南农业学报,2013,26(1):17-21.
[20]张亚勤,杨 华,邱正高,等.几个玉米自交系的主要农艺性状的配合力和遗传参数分析[J].西南农业学报,2013,26(2):406-414.
[21]王兵伟,黄安霞,覃永嫒,等.20个糯玉米自交系几个主要性状的配合力分析[J].西南农业学报,2013,25(1):22-27.
[22]高之仁.数量遗传学[M].第1版.成都:四川大学出版社,1986.
[23]Lewis R S, Goodman M M. Incorporation of tropical maize germplasm into inbred lines derived from temperate × temperate-adapted tropical line crosses: agronomic and molecular assessment[J].Theor Appl Genet,2003,107:798-805.
[24]高祥扩,杨克昌,黄清梅,等.玉米4 个穗部性状的遗传及相关分析[J].玉米科学,2012,20(4):9-14.
[25]Holland J B, Goodman M M. Combining ability of tropical maize accessions with US germplasm[J].Crop Sci,1995,35:767-773.
[26]李勇成,番兴明,胡 云,等.外来热带、亚热带玉米自交系与云南地方玉米自交系的配合力研究[J].玉米科学,2009,17(3):45-50.
(责任编辑 王家银)
StudyonCombiningAbilityandGeneticParametersbetweenTropical,SubtropicalMaizeInbredLinesandTemperateInbredLines
DUAN Zhi-li1, YANG Xiao-ting1 *, CHEN Hong-mei2 **, XI Cong-fang1, WANG Yan-fen2, WANG Jing1, LI Jian-bin1 **
(1.College of Agriculture and Biotechnology, Yunnan Agricultural University, Yunnan Kunming 650201, China; 2.Institute of Food Crop, Yunnan Academy of Agricultural Sciences, Yunnan Kunming 650205, China)
In this study, 36 crosses were made with four tropical, subtropical maize inbred lines as test-crossed and nine temperate inbred lines as tested ones by NCII genetic design, and were evaluated in Kunming, Qujing and Dehong of Yunnan province. General combining ability(GCA),Special combining ability(SCA) and genetic parameters of nine agronomic traits of these inbred lines and crosses were recorded. The results showed that the CRN3760-1-2-2-1-1-2 (tropical maize inbred line) has the highest yield GCA, next were YML46,[A1112×T(5003×Yun146)]##-1-1-1-1-B, 2870263-4-2-1-B-B-B-B-B, IRF321, 8723 and 23X, and they were had significant atP≤0.05. Above inbred lines could be used in developing the crosses with higher heterosis. Four cross combinations with having significant higher yield SCA and high yield potential were selected, which were CRN3760-1-2-2-1-1-2×23X, CRN3760-1-2-2-1-1-2 × 2870263-4-2-1-B-B-B-B-B, CRN3760-1-2-2-1-1-2 ×[E1112×T(5003×Yun146)]##-1-1-1-1-B and YML46 × 8723, and their performance of high yield and yield stability would be further tested in Yunnan. The hereditary of the nine agronomic traits were mainly under control by the additive gene effect. The narrow heredity of axis diameter, 100-kernel weight and grain rate were higher than 55.2 %, which showed that the variation of these traits were affected mainly by hereditary, and they should be selected at early generation. The narrow heredity of ear diameter, sterile length and row number were lower than 23.2 %, and they should be selected at the late generation.
Tropical and subtropical maize; Germplasm; Combining ability; Heterosis; Genetic parameter
1001-4829(2017)3-0497-08
10.16213/j.cnki.scjas.2017.3.003
S513
A
2015-03-05
国家重点基础研究发展计划(973计划)(2011CB 100402);云南省重点新产品开发计划项目(2012BB012);国家科技支撑计划课题(2012BAD40B01)
段智利(1978-),女,硕士,副研究员,主要从事作物遗传育种研究,E-mail:292769220@qq.com;*为共同第一作者,杨晓婷(1989-),女,云南大理人,硕士研究生,主要从事作物遗传育种研究,E-mail:1034512548@qq.com,**为通讯作者,陈洪梅,E-mail:chenhm9072@sina.com,李建宾,E-mail:rikehe@hotmail.com。
