葡萄白藜芦醇合酶基因的克隆与生物信息学分析
2017-10-19,,,,,,
,,, ,,,
(1.山东大学生命科学学院,微生物技术国家重点实验室,山东济南 250100;2.重组蛋白基因检测技术国家地方联合工程实验室,山东博奥克生物科技有限公司,山东聊城 252000)
葡萄白藜芦醇合酶基因的克隆与生物信息学分析
翟逸1,2,刘钦松2,吴永坤2,马云2,张念强2,石党委2,祁庆生1,*
(1.山东大学生命科学学院,微生物技术国家重点实验室,山东济南 250100;2.重组蛋白基因检测技术国家地方联合工程实验室,山东博奥克生物科技有限公司,山东聊城 252000)
以葡萄总RNA为模板,利用RT-PCR的方法从葡萄中扩增获得一条白藜芦醇合酶基因完整的cDNA序列,命名为RS。利用生物信息学软件对其核酸和蛋白质序列进行分析,结果表明,该序列长1179 bp,与已报道的葡萄白藜芦醇合酶基因的序列相似性达到94%~99%,氨基酸序列相似性为96%~99%;RS基因编码392个氨基酸,氨基酸序列含有完整的芪合酶家族的特征序列GVLFGPGLT和活性中心序列GCYAGGTVLR;预测的分子量为42.78 kDa,理论等电点为6.57,不稳定参数为35.92,在分类上属于稳定性蛋白;二级结构主要以α-螺旋、无规则卷曲以及β-折叠为主,其中α-螺旋含量为44.13%、无规则卷曲含量为26.53%,β-折叠含量为17.66%。
葡萄,白藜芦醇合酶,基因克隆,序列分析
白藜芦醇(Resveratrol,Rs),是植物受到外来生物或非生物等环境条件胁迫时产生的一种具有芪类结构的非黄酮类多酚化合物,可使植物抵抗机械损伤、紫外线照射、细菌与真菌等病原的侵染。白藜芦醇主要存在于葡萄、花生、桑椹和虎杖等植物中[1],其化学结构如图1所示。白藜芦醇最初是作为一种植物“杀菌素”为人所知,随着研究的不断深入,研究者发现白藜芦醇还具有抗衰老、抗肿瘤、预防心血管疾病,抗神经性病变及提高机体免疫力等多种生物活性,在食品、保健品、化妆品及生物制药等行业中具有很大的应用潜力和广阔的发展前景[2-5]。
图1 反式白藜芦醇和顺式白藜芦醇结构图Fig.1 The structure of transresveratrol(a)andcis resveratrol(b)
在植物体中,白藜芦醇通过苯丙氨酸裂解酶(PAL)、肉桂酸-4-羟化酶(C4H),4-香豆酸-CoA连接酶(4CL)和白藜芦醇合酶(RS)四种酶来合成。RS是合成途径中的关键酶,它催化1分子的对香豆酰-CoA和3分子的丙二酰-CoA生成1分子的白藜芦醇[6-7]。目前,白藜芦醇主要是从葡萄皮、葡萄籽和中药虎杖等植物的组织中进行提取。但由于白藜芦醇在植物组织中的含量较低,再加上受原料来源、植物生长的季节性与地域条件的限制以及提取工艺等诸多因素的影响,导致从植物中提取、分离高纯度的白藜芦醇成本过高。白藜芦醇的化学合成又由于反应过程较复杂,对环境易造成污染等不足而限制了其进一步的应用。
近年来,随着合成生物学与代谢工程的迅速发展,利用微生物合成具有重要应用价值的植物源代谢产物越来越受到研究者的重视[8]。目前已有学者对生物合成白藜芦醇进行了相关研究,但相对葡萄来源的白藜芦醇合酶基因的研究相对较少[9-12]。本研究以葡萄总RNA为模板,利用RT-PCR扩增获得了一条1179 bp的完整编码序列RS,并对其进行了序列比对及生物信息学等相关分析,将其确定为葡萄的白藜芦醇合酶基因,为下一步利用基因工程技术进行白藜芦醇的生物合成提供了基因资源。
1 材料与方法
1.1材料与仪器
葡萄(Vitisvinifera)叶片 取自山东九州生物产业园有限公司的葡萄种植园;DP432植物总RNA提取试剂盒、大肠杆菌DH5a感受态细胞、分子量标准D2000 DNA Marker 天根生化科技(北京)有限公司;PrimeScriptTMII 1st Strand cDNA Synthesis Kit、TaKaRa LA Taq®、Takara MiniBEST Agarose Gel DNA Extraction Kit Ver.4.0、Takara pMDTM18-T Vector Cloning Kit、TaKaRa MiniBEST Plasmid Purification Kit Ver.4.0、T4 DNA Ligase 宝生物工程(大连)有限公司;LB培养基 蛋白胨10 g,酵母膏5 g,NaCl 10 g,蒸馏水1000 mL;其它试剂 分析纯。
TECHNE TC-512 PCR仪 英国Bibby Scientific Ltd;DYCP水平电泳仪 北京六一仪器厂;Tanon 1600R全自动数码凝胶成像分析系统 上海天能科技有限公司;5430R多功能台式高速离心机 上海艾本德生物技术国际贸易有限公司;YXQ-LS-50A立式压力蒸汽灭菌器、BGZ-240 电热鼓风干燥箱 上海博迅实业有限公司;DTY-5A智能恒温循环器 北京德天佑科技发展有限公司;JE203电子天平 上海浦春计量仪器有限公司;PHS-3C pH计 上海虹益仪器仪表有限公司;DTY-900 超净工作台 苏州苏洁净化设备有限公司。
1.2实验方法
1.2.1 葡萄叶片总RNA的提取 选取幼嫩、长势良好的葡萄叶片,摘下后迅速放入盛有液氮的小型容器中带回实验室。将葡萄叶片从容器中取出,放入预先以焦碳酸二乙酯(Diethy pyrocarbonate,DEPC)水浸泡并高温处理过的研钵中,加入适量液氮迅速研磨至粉末状,取适量转至无核酸酶的1.5 mL离心管中,根据植物总RNA提取试剂盒操作使用说明书提取葡萄叶片总RNA,-80 ℃冰箱保存备用。
1.2.2 PCR扩增引物的设计与合成 根据已报道的葡萄RS基因的序列(GenBank登录号:NM001281044.1),利用GeneTool分别设计扩增引物,上游引物RS-1F:5′ATGGCTTCAGTCGAGGAAT3′,下游引物R:RS-1R:5′ATTTGTAACCGTAGGAATGC3′。引物送生工生物工程(上海)股份有限公司进行合成。
1.2.3 白藜芦醇合酶基因RT-PCR扩增 以1.2.1中提取的葡萄叶片总RNA为模板,按照Prime ScriptTMII 1st Strand cDNA Synthesis Kit说明书进行逆转录,合成第一链cDNA;然后以合成的cDNA为模板,RS-1F:5′-ATGGCTTCAGTCGAGGAAT-3′、RS-1R:5′-ATTTGTAACCGTAGGAATGC-3′为引物,进行目的基因的PCR扩增,其反应体系为50.0 μL,包括5.0 μL缓冲Buffer,8.0 μL的dNTPs,各5.0 μL的RS-1F、RS-1R,5.0 μL的cDNA模板,21.5 μL灭菌ddH2O和0.5 μL 的LA Taq酶;PCR扩增条件:94 ℃预变性5 min;94 ℃变性40 s、56 ℃退火40 s、72 ℃延伸110 s,35个循环;最后72 ℃延伸10 min;反应完毕,取适量PCR扩增产物以1.0%琼脂糖凝胶进行电泳检测。
1.2.4 目的基因的TA克隆与测序 经电泳检测后,以琼脂糖凝胶回收试剂盒纯化回收目的片段。pMDTM18-T与纯化片段的连接体系为:pMDTM18-T Vector 1.0 μL,纯化目的片段4.0 μL,Solution I连接反应液5 μL。将上述反应体系缓慢混合均匀,在4 ℃条件下过夜连接。将10 μL的连接产物加入E.coliDH5α感受态细胞中,经热击转化后涂布氨苄抗性平板,37 ℃过夜培养;挑取白色单克隆进行液体培养、质粒提取,经PCR扩增验证重组质粒中插入片段大小正确后,送生工生物工程(上海)股份有限公司进行测序。
1.2.5 RS基因序列生物信息学分析 在NCBI中利用BLAST与保守结构域查询(Conserved Domain Search,CD Search)在线程序等对RS基因进行序列比对及保守结构域查询;DNAMAN 5.22与Clustal X等生物学软件进行多序列比对分析;RS蛋白的一级结构、二级结构及理化性质等预测通过ExPASY ProtParam Server在线服务器进行,三级结构预测采用自动建模的方式在SWISS-MODEL服务器上进行。
2 结果与分析
2.1葡萄叶片总RNA的提取
提取的葡萄叶片总RNA电泳检测结果如图2所示,从上往下依次为28 S、18 S和5 S的RNA。从图中可以看出,28 S、18 S条带完整、清晰,5 S条带则非常暗淡。这表明提取的葡萄总RNA无降解,质量较高,可满足下一步目的基因扩增的实验要求。
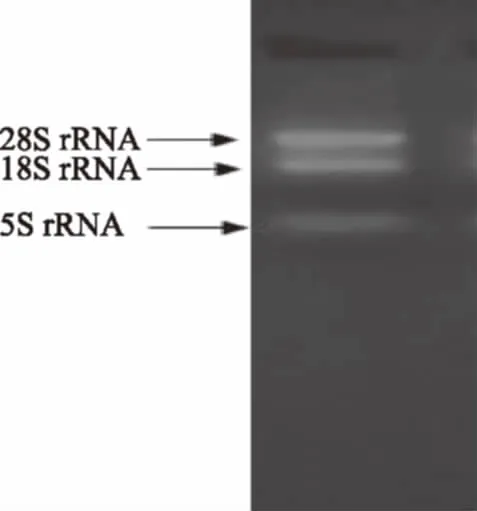
图2 葡萄叶片总RNA电泳检测Fig.2 The electrophoresis detection oftotal RNA of grape leaves
2.2白藜芦醇合酶基因RS的克隆
将提取的葡萄叶片总RNA逆转录合成第一链cDNA,以RS-1F和RS-1R为引物,cDNA为模板,经PCR扩增获得一条约1200 bp的特异性电泳条带,如图3所示,其大小与预期扩增产物(1179 bp)相符。
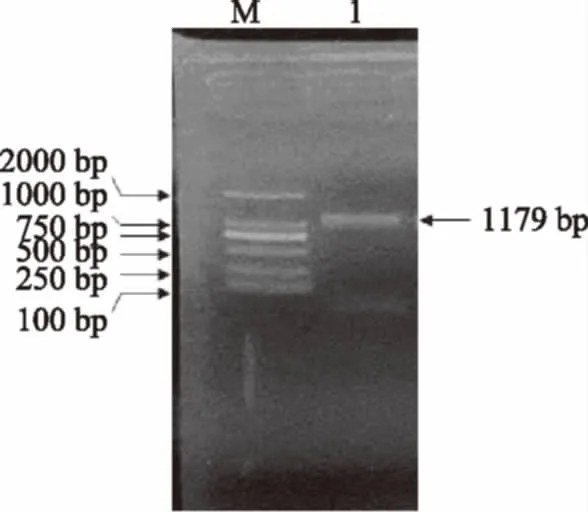
图3 白藜芦醇合酶基因RS的RT-PCR扩增Fig.3 RT-PCR amplification ofresveratrol synthase RS gene
2.3重组质粒的构建与筛选
在凝胶成像系统下将2.2中的特异性扩增电泳条带切下,纯化回收后与pMD18-T载体连接,转化E.coliDH5α并涂布在含有IPTG、X-gal和氨苄青霉素抗性的筛选平板上。培养后从平板上挑取白色单克隆转接于含氨苄青霉素抗性的液体LB中过夜培养并提取质粒。以提取的质粒为模板,RS-1F和RS-1R为引物,经PCR扩增获得一条与图3中特异性条带大小一致的电泳条带,如图4所示。这表明,目的片段已成功插入到克隆载体中,重组质粒构建成功。
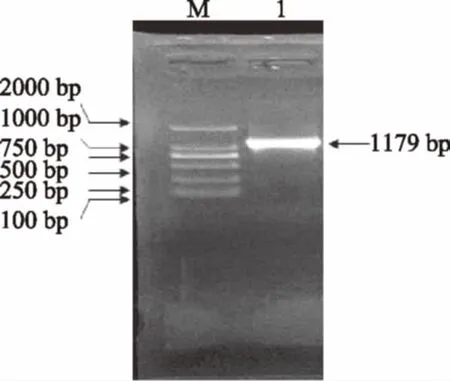
图4 阳性克隆的PCR扩增验证Fig.4 The verification of positive clone by PCR amplification
2.4RS基因的序列分析
阳性重组质粒的测序结果表明,插入片段包含一个完整的开放阅读框,片段的大小为1179 bp,将其命名为RS。在NCBI中BLAST(https://blast.ncbi. nlm.nih.gov)的序列比对结果表明,该序列与NCBI中已报道的葡萄白藜芦醇合酶基因的序列相似性达到94%~99%,其中与NM_001281044仅有3个碱基的差异,氨基酸序列相似性为96%~99%。这表明所克隆片段为葡萄的白藜芦醇合酶基因,其核酸与翻译的氨基酸序列如图5所示。

图5 RS基因的核酸序列和翻译的氨基酸序列Fig.5 The nucleic acid and translatedamino acid sequences of RS gene注:单划线序列表示芪合酶活性部位,标星号的为保守氨基酸;方框中序列表示芪合酶家族的特征序列。
2.5RS基因的生物信息学分析
2.5.1 一级结构预测与分析 葡萄白藜芦醇合酶RS基因编码的RS蛋白由392个氨基酸残基组成,预测的分子量为42.78 kDa,等电点为6.57。计算的不稳定参数为35.92,在分类上属于稳定性蛋白,这有利于后续进一步的研究工作。
2.5.2 RS基因结构域分析 白藜芦醇合酶属于芪合酶家族,通过NCBI在线软件对RS基因氨基酸序列的结构域分析结果如图5所示。该氨基酸序列具有完整的芪合酶家族的特征序列GVLFGPGLT,同时具有芪合酶家族活性中心序列GCYAGGTVLR,其中Cys164为芪合酶家族中保守的催化活性氨基酸,在酶的催化作用中具有重要的功能。
2.5.3 二级结构预测与分析 ExPASY ProtParam Server在线服务器对RS蛋白二级结构预测结果如图6所示。从图中可以看出,该RS蛋白的二级结构以α-螺旋、无规则卷曲以及β-折叠为主,其中α-螺旋含量为44.13%、无规则卷曲含量为26.53%,β-折叠含量为17.66%。

图6 RS蛋白的二级结构预测Fig.6 The predicted secondary structure of RS protein注:h:α-螺旋;c:无规则蜷曲;e:β-折叠。
2.5.4 三级结构预测与分析 将RS蛋白序列在SWISS-MODEL服务器中经BLAST比对搜索,获得了一条序列相似性为96.43%的模板3tsy.1.A,如图7所示。采用自动建模方式,以3tsy.1.A为模板预测RS蛋白的三级结构如图8。从图中可以看出,在该结构中含有较多的α-螺旋和无规则卷曲,这也与2.5.3中RS蛋白的二级结构预测结果相符合。
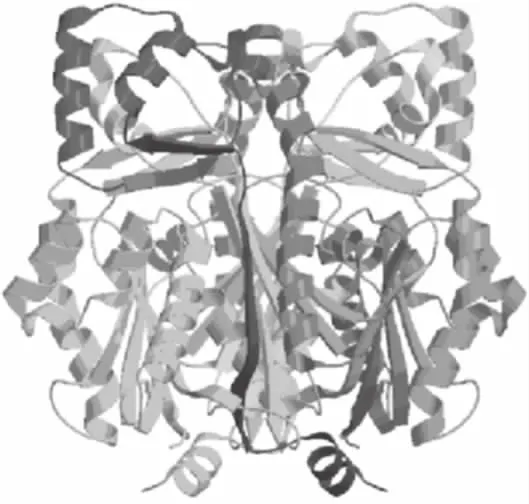
图7 模板3tsy.1.A三级结构Fig.7 Tertiary structure of template 3tsy.1.
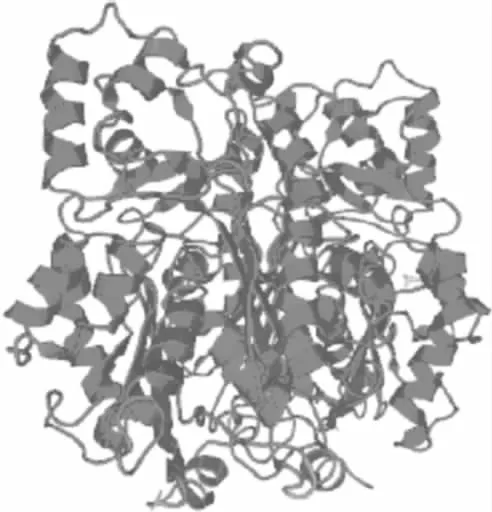
图8 预测的RS蛋白三级结构Fig.8 Predicted tertiary structure of RS protein
3 结论
白藜芦醇合酶(EC 2.3.1.95)又被称为3,4,5-三羟基二苯乙烯合酶,是芪合酶(Stiibene Synthase,STS)家族中的一种,它以4-香豆酰辅酶A和丙二酰辅酶A为底物合成白藜芦醇[13]。本研究通过RT-PCR扩增从葡萄总RNA中获得一条长度为1179 bp的基因片段,命名为RS,其在GenBank中的登录号为:KX688208。该基因片段的核酸和氨基酸序列与NCBI中已报道的葡萄来源的白藜芦醇合酶相似性分别达到94%~99%和96%~99%,并且具有典型的芪合酶家族特征序列GVLFGPGLT和活性中心序列GCYAGGTVLR。这些结果表明文中分离的RS基因为葡萄的白藜芦醇合酶基因,为下一步利用基因工程的方法进行生物合成白藜芦醇奠定了基础。
[1]Langcake P,Pryce R. The production of resveratrol byVitisviniferaand other members of the vitaccae as a response to infection or injury[J]. Physiological and molecular plant pathology,1976,9(1):77-86.
[2]Whitlock N C,Baek S J. The anticancer effects of resveratrol:modulation of transcription factors[J]. Nutrition and cancer,2012,64(4):493-502.
[3]Sun A Y,Wang Q,Simonyi A,et al. Resveratrol as a therapeutic agent for neurodegenerative diseases[J]. Molecular neurobiology,2010,41:375-83.
[4]Li W,Xiaoqian Ma X Q,Li N,et al. Resveratrol inhibits Hexokinases II mediated glycolysis in non-small cell lung cancer via targeting Akt signaling pathway[J]. Experimental Cell Research,2016,349(2):320-327.
[5]Ozcan P,Ficicioglu C,Kizikale Q,et al. Protective effect of resveratrol against oxidative damage to ovarian reserve in female Sprague-Dawley rats[J]. Reproductive biomedicine,2015,31(3):404-410.
[6]Vogt T. Phenylpropanoid biosynthesis[J]. Molecular Plant,2010,(3):2-20.
[7]Ferrer J L,Austin M B,Stewart J C,et al. Structure and function of enzymes involved in the biosynthesis of phenylpropanoids[J]. Plant physiology and biochemistry,2008,46:356-70.
[8]戴住波,朱欣娜,张学礼. 合成生物学在微生物细胞工厂构建中的应用[J]. 生命科学,2013,25(10):943-949.
[9]Watts K,Lee P,Schmidt D C. Biosynthesis of plant specific stilbene polyketides in metabolically engineeriedEschrichinacoli[J]. BMC Biotechnology,2006,6(22):1-12.
[10]梁景龙,郭丽琼,孙萍,等. 产白藜芦醇大肠杆菌基因工程菌的构建[J]. 食品工业科技,2013,34(23):138-141.
[11]汪建峰,张嗣良,王勇. 大肠杆菌中从头合成白藜芦醇途径的设计及优化[J]. 中国生物工程杂志,2014,34(2):71-77.
[12]Schwekendiek A,Pfeffer G,Kindl H. Pine stilbene synthase cDNA,a tool for probing environmental stress[J]. Febs Letters,1992,13,301(1):41-44.
CloningandsequenceanalysisofaresveratrolsynthasegenefromVitisvinifera
ZHAIYi1,2,LIUQin-song2,WUYong-kun2,MAYun2,ZHANGNian-qiang2,SHIDang-wei2,QIQing-sheng1,*
(1.State Key Laboratory of Microbial Technology,College of Life Science,Shandong University,Jinan 250100,China;2.State and Local Joint Engineering Laboratory of Recombinant Protein and Gene Detection Technology,Shan dong Boaoke Biotechnology Co.,Ltd.,Liaocheng 252000,China)
A complete cDNA sequence of resveratrol synthase gene was obtained fromVitisviniferatotal RNA by RT-PCR method,named RS. The results of multiple sequence alignment showed that RS was of 1179 bp with 94%~99% nucleic acid sequence similarity and 96%~99% amino acid sequence similarity to the reportedVitisviniferaresveratrol synthase,respectively. Domain analysis results revealed that the characteristics sequences GVLFGPGLT and the active site sequences GCYAGGTVLR of stilbene synthase family were found in cloned RS amino acids sequence. The predicted molecular weight was of 42.78 kDa and the theoretical isoelectric point was of 6.57. The calculated instability parameter was of 35.92,which indicated that this protein was s
Table in classification. The secondary structure of resveratrol synthase mainly containedα-helix,random curling andβ-sheet,which content was of 44.13%,26.53% and 17.66%,respectively.
Vitisvinifera;resveratrol synthase;gene clone;sequence analysis
TS201.3
A
1002-0306(2017)19-0096-04
10.13386/j.issn1002-0306.2017.19.018
2017-04-14
翟逸(1980-),男,博士,研究方向:微生物分子生物学与基因工程,E-mail:zhaiyi1218@163.com。
*通讯作者:祁庆生(1966-),男,博士,教授,研究方向:微生物合成与代谢调控,E-mail:qiqingsheng@sdu.edu.cn。
