玉米群体生长与光截获的动态模拟及应用
2017-10-13曹亦兵黄收兵王媛媛夏雨晴孟庆锋陶洪斌王璞
曹亦兵,黄收兵,王媛媛,夏雨晴,孟庆锋,陶洪斌,王璞
玉米群体生长与光截获的动态模拟及应用
曹亦兵,黄收兵,王媛媛,夏雨晴,孟庆锋,陶洪斌,王璞
(中国农业大学农学院,北京 100193)
【目的】群体光截获对玉米生长发育及产量形成均有重要的影响,合理的群体结构直接决定着玉米产量的增加与稳定。群体结构的测量较为繁琐,费时费力,且不能与当地太阳辐射情况充分结合,从而无法对群体结构和功能做出整体的评价。因此,有必要构建合理、简便的玉米群体光截获动态模型,结合群体不同生育期的形态指标与光辐射情况,对群体结构做出快速而精准的评价,为不同区域玉米品种选择、播种密度确定、以及后期管理等提供参考意见。【方法】将玉米群体结构模型与光分布模型相结合,建立简易可靠的群体光截获动态模型。该模型可模拟玉米整个生育期群体光截获的动态变化,并对不同生育期群体结构做出相应的评价。同时,在中国农业大学吴桥实验站开展了两个田间试验,对模型的准确性和应用功能进行了验证。试验均为三因素,分别为品种、密度和氮肥处理,其中试验一:设置2个品种(郑单958,丹玉405),3个密度(4.5×104,6.0×104,7.5×104株/hm2)和3个氮肥处理(不施氮;施氮180 kg·hm-2,于播前、大喇叭口期和吐丝期以1﹕4﹕1的比例施入;施氮270 kg·hm-2,于播前、大喇叭口期和吐丝期以4﹕4﹕1的比例施入);试验二:设置1个品种(郑单958)、1个密度(8.25×104株/hm2)和6个播期(4月20日,5月5日与20日,6月4日与18日,7月3日)。在两个试验中,对玉米不同生育期的群体形态指标、干物重、及产量均进行了测定。【结果】吐丝期和灌浆中期的群体光截获率模拟值与测量值的相关系数分别为0.91和0.85,均达到显著相关。运用该模型对两个田间试验数据的模拟结果显示,光截获率(量)随灌浆期的推进呈先增后减的趋势,吐丝后50 d光截获衰减加速。在试验地区,密度超过6×104株/hm2时,光截获随密度变化不大;播期为6月中上旬时形成的玉米群体具有较为理想的光截获能力,结构功能潜力较大。产量与灌浆期有效光辐射呈正相关,但相关性随生育期推进逐渐减弱,表明灌浆后期产量的增加对群体光截获的需求减弱。在玉米生长发育后期物质转移对产量的贡献可能发挥着更加重要的作用。产量与群体光截获相关性的动态变化与品种有关,紧凑株型优于平展型。【结论】选择株型紧凑且生育后期群体结构维持较好的玉米品种,通过增加密度可进一步增加夏玉米产量。调整播期增强灌浆初期光合效率也是有效的增产方法之一。
玉米;群体结构;模型;光截获; 密度;播期;产量
0 引言
【研究意义】群体结构是影响玉米光合特性和产量的重要指标之一,理想的群体结构有助于光线在冠层合理分布,显著提高作物群体光合特性和生物学产量[1-3]。群体结构是一个复杂的综合指标,包括叶面积指数、叶面积分布、茎叶夹角、叶向值和株高等,且诸多指标均随生育进程的推进不断变化[4]。群体结构是一个动态变化的过程,任何一个指标均可影响群体光分布的情况。另外,太阳高度和方位也是影响作物群体光空间分布的重要因素之一[5]。因此,运用传统方法测定和描述群体结构及其光分布特征是一个复杂、耗时且精确度难以控制的过程。【前人研究进展】基于试验数据和模型模拟的研究手段能够在一定程度上简化群体结构特征和光分布,同时提高效率和精确度。现有一系列模型,例如群体结构-功能模型[6-7]、3D结构模型[8-9]和生长模型[10]等,均将光分布和光截获作为重要的模拟部分,并试图将不同的模型相结合,以此准确地模拟群体光合特性。其中,群体叶面积指数或叶片空间分布是以上模型考虑的重要参数之一。例如,在群体结构-功能模型中,群体叶片的空间分布,尤其是穗位层叶片可决定冠层总体的光合性能和物质生产[11]。在3D结构模型中,冠层不同层次的群体结构,单个叶片的空间取向均是重点模拟的指标,以期准确地模拟群体光分布情况,并对群体结构做出评价[9]。但是,上述模型均是模拟某个生育期或某个时间段的群体结构,而对其进行整个生育期动态变化的模拟却鲜有报道。在模拟群体光分布的过程中,太阳高度和方位随玉米生育进程的动态变化和对玉米群体光分布的影响没有重视。这些因素均是模拟群体整个生育期光合性能的重要因子,并最终影响产量模拟的准确性。其次,优化群体光合性能的目的是增加干物质累积,促进产量的提高,但是上述模型没有建立群体光分布与产量的具体关系,从而难以评价不同类型群体结构及其光分布对产量的贡献。同时,现有的群体光分布模型逐渐趋于复杂,所需参数较多,任何一个参数的微小变动均可影响模拟结果的准确性和稳定性,使模拟过程更加繁琐。【本研究切入点】如何在保证模拟结果准确性的同时减少输入参数,提高模型的简便性和可操作性,对于指导农业生产非常重要。在简单的田间试验或以往的试验数据基础上,借助简易模型能够快速地对玉米群体结构做出评价,对于不同区域玉米品种选择、播种密度、以及后期管理同样有非常重要的意义。【拟解决的关键问题】本研究利用玉米群体结构模型与光分布模型,拟建立一个简便可靠的玉米群体光截获动态模型,以期对不同生育阶段群体结构做出评价。
1 材料与方法
1.1 模型设计
该模型将Chelle等[12]和Maddonni等[13]提出的光辐射模型与Wilson[14]和Welles等[15]提出的冠层结构模型进行改进和整合。Maddonni等[13]提出群体有效光截获公式(the fraction of incident photosynthetically active radiation intercepted by canopy,fIPAR)可表达为:
f=1--K×LAI(1)
式中,为消光系数,为叶面积指数。
Welles等[15]提出光线在群体中的穿透率为:
(,)=exp[-(,)(,)] (2)
式中,(,)为光辐射的群体结构区域,为太阳天顶角,为太阳方位角,为叶片密度(每立方米群体中的叶片面积),(,)为光线在冠层中的路径。在测量过程中,太阳方位角将被平均,可在公式中忽略不记,所以公式(2)可简化为:
()=exp[-()()] (3)
由公式(1)、(3)可得,
()=1-f=1--K×LAI(4)
进而,由公式(3)、(4)可得,
exp[-()()]=-K×LAI(5)
最终得到公式(6):

其中,在单一的群体结构中,叶片密度与冠层高度和叶面积指数有关,可表示为:
(7)
光线在冠层群体中的路径()与冠层高度和太阳天顶角相关,可表示为:
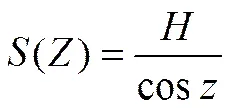
由公式(6)、(7)、(8)可知,消光系数可转换为:
(9)
根据Wilson[13]和Welles等[14]提出的冠层结构模型,光线在冠层群体中的辐射区域可表示为:

式中,叶茎夹角,为太阳天顶角。
由公式(9)、(10),将消光系数进一步转换为:

因此,冠层群体光截获模型最终表达为:
(12)
1.2 模型验证与应用
1.2.1 模型验证 试验一于2010—2011年在中国农业大学吴桥实验站(37°41′N,116°37′E)进行,其数据用于模型建立及验证。试验采取裂区设计,品种和密度为主因素,氮肥为副因素,每个处理3次重复,小区面积为60 m2。供试品种为郑单958(ZD 958)和丹玉405(DY 405);种植密度为4.5×104,6.0×104,7.5×104株/hm2,行距均为60 cm;氮肥处理为不施氮、施氮180 kg·hm-2(分别在播前、大喇叭口期和吐丝期以1﹕4﹕1的比例施入)、施氮270 kg·hm-2(分别在播前、大喇叭口期和吐丝期以4﹕4﹕1的比例施入)。同时,施用磷肥(重过磷酸钙)105 kg P2O5·hm-2、钾肥(K2SO4)120 kg K2O·hm-2和ZnSO415 kg·hm-2作为基肥。本试验于6月20播种,10月7日收获。
1.2.2模型应用 试验二于2009年在中国农业大学吴桥实验站进行,其数据以及部分试验一数据用于验证模型的应用效果。试验设置6个播期,具体播种时间及相应吐丝期和收获期见表1,采取随机区组设计,每个处理3次重复,小区面积为50 m2。供试品种为郑单958,设置密度为8.25×104株/hm2,行距为60 cm;氮肥用量180 kg N·hm-2(于播前和大喇叭口期分别以1﹕2的比例施入),其他肥料用量和施入时间同试验一。

表1 试验二的播种、吐丝和收获时间(月/日,2009)
1.2.3 测量指标和数据来源 叶面积指数(LAI):分别于吐丝期、灌浆期由传统长宽法测量全株叶面积。每个小区取样3株,单叶叶面积=长×宽×0.75,单株叶面积为单叶叶面积的总和。LAI=单株叶面积×单位土地面积内株数/单位土地面积。群体绿叶面积衰老速度用曲线方程=ae-cx/(1+e-cx)模拟,其中为灌浆期某一时刻相对吐丝期的绿叶面积(RGLA,%),为吐丝后天数,为RGLA的理论初始值(100%),为叶片衰老的启动参数,为衰老参数[16]。该方程可用于模拟玉米灌浆期叶面积指数的动态变化。
叶茎夹角:于吐丝期利用量角器侧量叶脉与茎边缘的夹角,每个小区测量10株。
透光率:于吐丝期、灌浆期采用LAI-2000(Li-Cor,Lincoln,NE,USA)在群体底层(地面)测量所得,每个小区不同位置测量4次。
光截获率(%)=1-透光率。
产量:每小区未取样区域收获玉米5行,每行4 m,称量所收果穗总鲜重。按平均鲜穗重从所收果穗中随机选取10穗,测定籽粒含水率,然后换算成实际产量(按14%折算含水率)。
太阳天顶角:数据来自NOAA ESRL(http://www. esrl.noaa.gov/gmd/grad/solcalc/azel.html)。同时测定吐丝期和灌浆期取样当天中午12点的太阳天顶角。
光合有效辐射:数据来自“中国气象科学数据共享服务网”泊头站(http://cdc.cma.gov.cn/),站点编号为54 618,38°05′N, 116°33′E,海拔132 m,距吴桥试验站约50 km。
光合有效辐射截获量:群体光合有效辐射截获量(MJ·m-2,下文用有效光截获表述)=f×,灌浆期光合有效辐射累计量(MJ·m-2)=Ʃ(day),灌浆期群体光合有效辐射累计截获量(MJ·m-2)=Ʃ f(day)×(day)。式中,f为群体截光率(%),f(day)为灌浆期每日群体光截获率,为光合有效辐射量(MJ·m-2),(day)为灌浆期每日光合有效辐射量,f和f(day)均为模拟值。
1.3 数据处理与分析
本研究运用SAS 9.4程序“Proc. Corr.”分析模型模拟值与田间试验测量值之间的相关性以及产量与光截获的相关性,整个模型在Excel 2010中运行。
2 结果
2.1 光截获模型的验证
为验证模型的准确性,使用试验一的研究结果分 别对玉米吐丝期和灌浆中期各处理光截获率的测量值与模拟值进行相关分析(图1)。结果表明,两者呈显著相关关系,且吐丝期相关系数高于灌浆中期,分别为0.91和0.85。

**, P<0.01显著。下同
2.2 模型应用
2.2.1 密度与群体光截获的关系 施氮量为180 kg·hm-2(播前﹕大喇叭口期﹕吐丝期=1﹕4﹕1)是吴桥地区较为常用的施氮方式,因此以本处理来反映吐丝后群体光截获随生育进程的变化情况。不同密度下,两品种群体光截获率(图2)和光截获量(图3)在整个灌浆期均呈现先升高后降低的趋势。光截获率在吐丝后30 d达到峰值。由于受阴雨寡照影响,有效光辐射在吐丝后20—30 d处于较低水平,导致光截获量波动较大。群体光截获率和截获量均在吐丝后50 d开始加速衰减。在4.5×104株/hm2密度下,光截获率(量)明显低于6.0×104和7.5×104株/hm2种植密度;而在6.0×104和7.5×104株/hm2密度下光截获量非常接近(图2—3)。
2.2.2 播期与群体光截获的关系 利用已经建立的模型,对试验二播期试验的数据进行分析整理。模拟结果显示,不同播期群体有效光截获率(量)在整个灌浆期呈现先升高后降低的趋势(图4),其中,光截获率随播期的推迟逐渐升高,即播期1—3群体处于较低的光截获水平,而播期4—6群体的光截获水平则较高,且播期4—6的数值非常接近(图4-A)。受不同阶段有效光辐射的影响,虽然整体规律表现一致(先升后降),但单个播期群体在灌浆期不同阶段的光截获量(MJ·m-2)波动较大(图4-B)。整个灌浆期光合有效辐射累积量(MJ·m-2)随播期推迟呈逐渐降低的趋势,而光合有效辐射累积截获量(MJ·m-2)随播期推迟呈先升高后降低的趋势,其中播期3的数值最高(图5)。

图2 灌浆期郑单958(ZD 958)和丹玉405(DY 405)在3个密度下的群体光截获率模拟值

图3 灌浆期郑单958(ZD 958)和丹玉405(DY 405)在3个密度下的群体光合有效辐射截获量模拟

图4 不同播期下郑单958群体灌浆期光合有效辐射截获率模拟值(A)与十天平均光截获量模拟值(B)随生育进程的变化

图5 灌浆期不同播期群体光合有效辐射累计量与累计截获量
2.2.2 产量与群体光截获的关系 运用本模型对两个不同试验的产量和光合有效辐射数据进行综合分析后发现,籽粒产量与光合有效辐射均呈正相关关系(图6—7)。其中,播期试验(试验二)的相关性高于密度和氮肥试验(试验一)。由图6可知,产量与灌浆期光合有效辐射累计量的相关性(0.86)高于与光辐射截获量的相关性(0.80),且均达到显著相关。由图7可知,产量与灌浆期不同阶段(10 d平均值,试验一)群体有效光辐射截获量同样呈现正相关性,但相关系数较低且不显著,并随灌浆期的推进逐渐降低。郑单958相关系数的变动范围为0.31—0.21,丹玉405为0.55—0.33,后者在不同灌浆阶段均高于前者。
3 讨论
3.1 模型验证与分析
由不同时期群体光截获率模拟值与测量值的显著相关性可知(图1),该模型的准确性已达到较为理想的水平,可以用于评价玉米群体结构。对吐丝期和灌浆中期进行比较发现,测量值均高于模拟值,其主要原因是测量值未能排除底部衰老叶片对光截获的影响[4, 17]。底部衰老或干枯叶片可以拦截部分光照,从而影响光截获的测量值。两者的相关性在灌浆中期低于吐丝期,有以下四点原因:随着生育期的推进,叶片出现下披的现象,导致叶茎夹角略有增大[18],由于模型中未涉及此参数的校正导致模拟值略有偏差;随着底部衰老叶片增多,其对光截获率测量值的影响增大,而在模型中叶面积参数是有效绿叶面积更接近真实状态;未考虑雄穗对光辐射的截获作用[19-20],在一定程度上可能影响模型的准确性;另外,模型中叶面积指数为模拟值,并非每个生育期的真实测量值,因此造成了一定的模拟偏差。总体而言,与群体光截获的实测方法相比,该模拟方法不仅节省大量的时间和人力,而且可通过对群体不同生育期光截获的模拟分析,对群体结构的动态变化进行评价。

图6 籽粒产量与灌浆期光合有效辐射累积量和累积截获量的相关性(试验二)

图7 不同品种群体产量与灌浆期不同阶段光合有效辐射截获量的相关性
与现有的玉米群体光截获模型相比[6-10, 21],本模型将群体叶面积作为一个整体,模拟了整个群体的光截获状态,简化了模拟步骤,省略了大量参数,使模拟过程更加简洁,并且实现了群体光截获随生育期变化的动态模拟。在接下来的研究中,可以进一步细化光辐射在群体不同层次的分布,为深入研究光照、不同部位叶片对干物质生产及产量的影响奠定基础。
3.2 模型应用及分析
光照在群体内的分布和截获是评价作物群体结构的重要参数,但至今仍没有研究提出针对不同群体结构的光截获评价范围[6, 22-23],也鲜有光截获模型涉及籽粒产量[4]。可见精确模拟群体光截获具有一定难度,主要因为涉及群体光截获的影响因素较多,例如品种、密度、播期等。品种是调控群体结构的重要措施之一,本文中丹玉405比郑单958更为紧凑(18.0°和22.4°,吐丝期整株平均值),在高密度下有进一步提高光截获的潜力(图2—3)。但是,当密度大于6×104株/hm2时,群体光截获率(量)变化很小,尤其是华北平原地区最常用的品种郑单958,表明在当地通过增加密度进一步提升产量的空间有限,这点与徐丽娜等[24]的研究结果一致。在播期试验中,产量与群体光辐射截获量的相关性处于较低的水平=0.8(图6),表明品种郑单958在8.25×104株/hm2(播期试验的种植密度)密度下的群体结构并不理想;可能由于群体结构过于郁闭,截获的有效光辐射不能被充分利用。另外,产量与群体的光合能力(群体功能)、后期物质转运同样密切相关[25],这些因素也可影响产量与群体光辐射的相关性。
通过早播或者晚收能够提高灌浆期光合有效辐射累计量,从而达到高产的目的[26-27]。本研究中播期早于5月20日的群体光截获率处于较低的水平,说明群体结构的优势并没有得到充分发挥;播期晚于6月4日的群体光截获率则较高,且不同播期间数值非常接近。上述结果说明,晚于6月4日播种更利于群体结构功能的充分发挥。鉴于当地一年两熟的耕作制度(冬小麦-夏玉米),夏玉米整个生育期被控制在6月中下旬至9月底,即100 d左右[27-29]。夏玉米播种太早将影响冬小麦的产量,而播种太晚会显著降低夏玉米的产量[30]。因此,大幅度调整夏玉米播期的可能性不大。另外,吐丝开花期高温、阴雨等逆境条件同样制约着夏玉米播期的选择[26]。适当早播(6月中上旬)在提高群体光辐射利用效率的同时,也提高了避免高温、寡照等逆境的可能性。
由不同阶段光截获与产量的相关性可知(图7),灌浆初期光照对产量的影响大于灌浆的其他时期,可能与当地吐丝期前后的阴雨寡照环境有关。徐丽娜等[24]、陈建忠等[31]指出在该试验区域(黑龙港流域)7—8月的降雨量可达全年的70%—80%,日照时数为5.8 h左右。这段时间正值夏玉米吐丝、灌浆初期,“源”的物质生产能力不足,将直接影响夏玉米产量的进一步增加。
群体光截获与产量的相关性在整个灌浆期一直处于较低的水平,表明光照不足只是制约当地夏玉米增产的因素之一。由于灌浆后期群体结构维持功能衰减加快(吐丝后50 d左右,图2)以及光辐射量的减小,灌浆初期的相关性大于灌浆后期,表明在灌浆中后期物质生产对产量的贡献逐渐减弱,而物质转移随生育期进行变得愈加重要。
4 结论
通过利用玉米群体结构模型与光分布模型建立了简便有效的玉米群体光截获动态模型,该模型准确的描述玉米冠层的光辐射截获能力,并可结合当地的光辐射环境,对吐丝后整个生育期的群体光截获做出动态模拟,细化光辐射在群体内的分布。该模型结合吴桥地区大田试验分析发现,根据不同玉米品种形态特征,该试验地区夏玉米最适种植密度为6.0×104—7.5×104株/hm2,最适播期为6月中上旬。在现有的耕作措施和品种条件下,通过增加密度进一步增加夏玉米产量的空间有限,但通过调整播期增强灌浆初期光合效率,或选择株型紧凑且能在生育后期维持较好群体结构的品种是有效的增产方法。
致谢:感谢徐丽娜、朱金城博士在数据分析与处理方面给予的帮助与支持!
[1] 董树亭, 胡昌浩, 岳寿松, 王群瑛, 高荣岐, 潘子龙.夏玉米群体光合速率特性及其与冠层结构、生态条件的关系. 植物生态学与地植物学学报, 1992, 16(4): 372-378.
DONG S T, HU C H, YUE S S, WANG Q Y, GAO R Q, PAN Z L. The characteristics of canopy photosynthesis of summer corn and its relationship with canopy structure and ecological conditions., 1992, 16(4): 372-378. (in Chinese)
[2] 吕丽华, 陶洪斌, 夏来坤, 张雅杰, 赵明, 赵久然, 王璞. 不同种植密度下的夏玉米群体结构及光合特性. 作物学报, 2008, 34(3): 447-455.
LÜ L H, TAO H B, XIA L K, ZHANG Y J, ZHAO M, ZHAO J R, WANG P. Canopy structure and photosynthesis traits of summer maize under different planting densities., 2008, 34(3): 447-455. (in Chinese)
[3] 薛吉全, 梁宗锁, 马国胜, 路海东, 任建宏. 玉米不同株型耐密性的群体生理指标研究. 应用生态学报, 2002, 13(1): 55-59.
XUE J Q, LIANG Z S, MA G S, LU H D, REN J H. Population physiological indices on density-tolerance of maize in different plant type., 2002, 13(1): 55-59. (in Chinese)
[4] BIRCH C J, ANDRIEU B, FOURNIER C, VOS J, ROOM P. Modelling kinetics of plant canopy architecture concepts and applications., 2003, 19(4): 519-533.
[5] CHELLE M, EVERS J B, COMBES D, VARLET-GRANCHER C, VOS J, ANDRIEU B. Simulation of the three-dimensional distribution of the red: Far-red ratio within crop canopies., 2007, 176(1): 223-234.
[6] SARLIKIOTI V, DE VISSER P H, MARCELIS L F. Exploring the spatial distribution of light interception and photosynthesis of canopies by means of a functional-structural plant model., 2011, 107(5): 875-883.
[7] 王谦, 陈景玲, 孙治强. 用LI-2000冠层分析仪确定作物群体外红多面高度. 农业工程学报, 2005, 21(8): 70-74.
WANG Q, CHEN J L, SUN Z Q. Determination of the height of active surface of the crop colonies using Li-2000 Canopy Analyzer., 2005, 21(8): 70-74. (in Chinese)
[8] DROUET J L. MODICA and MODANCA: Modelling the three- dimensional shoot structure of graminaceous crops from two methods of plant description., 2003, 83(2): 215-222.
[9] 孔娅, 劳彩莲, 曹素云. 利用3D 模型模拟天空与叶面散射对玉米冠层截光率的影响. 农业工程学报, 2011, 27(5): 248-252.
KONG Y, LAO C L, CAO S Y. Effect of sky radiation and leaf scattering on maize canopy light interception by 3D modeling., 2011, 27(5): 248-252. (in Chinese)
[10] DROUET J L, PAGES L. GRAAL: A model of GRowth, Architecture and carbon ALlocation during the vegetative phase of the whole maize plant : Model description and parameterisation., 2003, 165(2/3): 147-173.
[11] PALMER A F E, HEICHEL G H, MUSGRAVE R B. Patterns of translocation, respiratory loss, and redistribution of14C in maize labeled after flowering., 1973, 13(3): 371-376.
[12] CHELLE M, ANDRIEU B. The nested radiosity model for the distribution of light within plant canopies., 1998, 111(1): 75-91.
[13] MADDONNI G A, CHELLE M, DROUET J L, ANDRIEU B. Light interception of contrasting azimuth canopies under square and rectangular plant spatial distributions: Simulations and crop measurements., 2001, 70(1): 1-13.
[14] WILSON J W. Inclined point quadrats., 1960, 59(1): 1-7.
[15] WELLES J, NORMAN J. Instrument for indirect measurement of canopy architecture., 1991, 83(5): 818-825.
[16] 刘开昌, 董树亭, 赵海军, 王庆成, 李宗新, 刘霞, 张慧. 我国玉米自交系叶片保绿性及其与产量的关系. 作物学报, 2009, 35(9): 1662-1671.
LIU K C, DONG S T, ZHAO H J, WANG Q C, LI Z X, LIU X, ZHANG H. Leaf stay-green traits in Chinese maize inbred lines and their relationship with grain yield., 2009, 35(9): 1662-1671. (in Chinese)
[17] 张子山, 杨程, 高辉远, 李耕, 刘鹏. 保绿玉米与早衰玉米叶片衰老过程中叶绿素降解与光合作用光化学活性的关系. 中国农业科学, 2012, 45(23): 4794-4800.
ZHANG Z S, YANG C, GAO H Y, LI G, LIU P. Relationship between photochemistry activity and decrease in chlorophyll content during senescence in leaves of stay green and quick-leaf-senescence inbred line of maize., 2012, 45(23): 4794-4800. (in Chinese)
[18] TIAN F, BRADBURY P J, BROWN P J, HUNG H, SUN Q, FLINT-GARCIA S, ROCHEFORD T R, MCMULLEN M D, HOLLAND J B, BUCKLER E S. Genome-wide association study of leaf architecture in the maize nested association mapping population., 2011, 43(2): 159-162.
[19] 黄华国, 窦宝成, 胡妮. 雄穗对玉米冠层热辐射方向性的影响分析. 红外与毫米波学报, 2011, 30(2): 120-123.
HUANG H G, DOU B C, HU N. Tassel effect on the thermal emission directionality of corn canopies., 2011, 30(2): 120-123. (in Chinese)
[20] ANDRIEU B, IVANOV N, BOISSARD P. Simulation of light interception from a maize canopy model constructed by stereo plotting., 1995, 75(1): 103-119.
[21] CABRERA-BOSQUET L, FOURNIER C, BRICHET N, WELCKER C, SUARD B, TARDIEU F. High-throughput estimation of incident light, light interception and radiation-use efficiency of thousands of plants in a phenotyping platform., 2016, 212(1): 269-281.
[22] BORRAS L, MADDONNI G A, OTEGUI M E. Leaf senescence in maize hybrids: Plant population, row spacing and kernel set effects., 2003, 82(1): 13-26.
[23] Koetz B, Baret F, Poilvé H, Hill J. Use of coupled canopy structure dynamic and radiative transfer models to estimate biophysical canopy characteristics., 2005, 95(1): 115-124.
[24] 徐丽娜, 陶洪斌, 黄收兵, 明博, 王璞. 黑龙港流域夏玉米产量提升限制因素. 生态学报, 2013, 33(1): 222-228.
XU L N, TAO H B, HUANG S B, MING B, WANG P. Analysis on the limiting factors to further improve yield of summer maize in Heilonggang River Valley., 2013, 33(1): 222-228. (in Chinese)
[25] ZHOU B, YUE Y, SUN X, WANG X, WANG Z, MA W, ZHAO M. Maize grain yield and dry matter production responses to variations in weather conditions., 2016, 108(1): 196-204.
[26] 李向岭, 李从锋, 侯玉虹, 侯海鹏, 葛均筑, 赵明. 不同播期夏玉米产量性能动态指标及其生态效应. 中国农业科学, 2012, 45(6): 1074-1083.
Li X L, Li C F, Hou Y H, Hou H P, Ge J Z, Zhao M. Dynamic characteristics of summer maize yield performance in different planting dates and its effect of ecological factors., 2012, 45(6): 1074-1083. (in Chinese)
[27] 明博, 朱金城, 陶洪斌, 徐丽娜, 郭步庆, 王璞. 黑龙港流域玉米不同生育阶段气象因子对产量性状的影响. 作物学报, 2013, 39(5): 919-927.
MING B, ZHU J C, TAO H B, XU L N, GUO B Q, WANG P. Effects of meteorological factors at different growth stages on yield traits of maize (L) in Heilonggang Basin., 2013, 39(5): 919-927. (in Chinese)
[28] 付雪丽, 张惠, 贾继增, 杜立丰, 付金东, 赵明. 冬小麦-夏玉米“双晚”种植模式的产量形成及资源效率研究. 作物学报, 2009, 35(9): 1708-1714.
FU X L, ZHANG H, JIA J Z, DU L F, FU J D, ZHAO M. Yield performance and resources use efficiency of winter wheat and summer maize in double late-cropping system., 2009, 35(9): 1708-1714. (in Chinese)
[29] WANG J, WANG E, YANG X, ZHANG F, YIN H. Increased yield potential of wheat-maize cropping system in the North China Plain by climate change adaptation., 2012, 113(3/4): 825-840.
[30] 薛庆禹, 王靖, 曹秀萍, 马薇, 冯利平. 不同播期对华北平原夏玉米生长发育的影响. 中国农业大学学报, 2012, 17(5): 30-38.
XUE Q Y, WANG J, CAO X P, MA W, FENG L P. Effect of sowing date and variety on growth and population characteristics of summer maize in North China Plain., 2012, 17(5): 30-38. (in Chinese)
[31] 陈建忠, 肖荷霞. 黑龙港流域气象生态因子对夏玉米穗粒数的影响. 河北农业科学, 2000, 4(3): 23-27.
Chen J Z, Xiao H X. Effects of ecoclimatical factors on grain number of summer maize in Heilonggang Area., 2000, 4(3): 23-27. (in Chinese)
(责任编辑 杨鑫浩)
Dynamic Simulation of Relationship Between Light Interception and Growth of Maize Population and Its Application
CAO YiBing, Huang ShouBing, Wang YuanYuan, Xia YuQing, Meng QingFeng, Tao HongBin, Wang Pu
(College of Agronomy and Biotechnology, China Agricultural University, Beijing 100193)
【Objective】The population light interception (PLI) has a great impact on growth, development, and grain production of maize (L.) in interaction with canopy structure of maize population. It costs amounts of time and labor to measure the morphology index in the evaluation of maize canopy structure. And it is impossible to integrate solar radiation into the evaluation of canopy structure. Therefore, it is quite necessary to develop a simple but feasible model that combine canopy structure and solar radiation to improve the evaluation of maize canopy structure, and thus to guide farmers in crop management.【Method】Based on the canopy structure model and light distribution model, a simple and reliable light interception model of maize canopy was developed. In order to evaluate the prediction accuracy of this model, two field trials were conducted at Wuqiao Experimental Station of China Agricultural University in 2009-2011. Trial 1 was set up as a split-plot design with varieties (ZD 958 and DY 405) and plant densities (45 000, 60 000, and 75 000 plant/hm2) as main factors, and nitrogen management (No nitrogen; 180 kg·hm-2N applied at sowing﹕13-leaf stage﹕silking stage=1﹕4﹕1; 270 kg·hm-2N applied at sowing﹕13-leaf stage﹕silking stage =4﹕4﹕1) as second factor. Trial 2 was in a randomized complete block design, including one variety (ZD 958), one plant density (82 000 plant/hm2), and six sowing dates (April 20, May 5 and 20, June 4 and 18, and July 3, 2009). 【Result】The simulated PLI rates were significantly correlated with measured PLI rates at silking stage and mid kernel-filling stage, withvalues of 0.91 and 0.85, respectively. During the entire kernel-filling stage, the PLI values first increased and then decreased, and the reduction became more obvious on 50 days after silking. Moreover, the PLI values remained at the highest level when plant density was greater than 60 000 plant/hm2. Additionally, population structure under sowing date of June 4 achieved the highest PLI, demonstrating its great potential of structure-function for the yield improvement. PLI and yield were positively correlated, while their correlation coefficient declined over time during grain-filling period, indicating that the dry matter allocation might play more important roles on yield during the late grain-filling period. The decline of correlation coefficient also depended on variety; variety with erecter leaves could remain a higher coefficient value over time. 【Conclusion】The potential to further improve maize yield through increasing plant density (>75 000 plant/hm2) is limited at experimental site. Some strategies should be concerned, such as (1) applying varieties with erect leaves and long-lasting canopy structure persistence during late grain-filling period, and (2) adjusting sowing date according to climate situation to increase population photosynthesis at early grain-filling period.
maize (L.); canopy structure; model; light interception; plant density; sowing date; yield
2016-08-25;
2017-03-14
国家玉米产业技术体系(201203031)、“粮食丰产增效科技创新”重点专项(SQ2017YFNC050062)
黄收兵,E-mail:huangshoubing@cau.edu.cn
联系方式:曹亦兵,E-mail:784298412@qq.com。
