不同氮效率水稻根系形态和氮素吸收利用与产量的关系
2017-10-13李娜杨志远代邹孙永健徐徽何艳严田蓉蒋明金郭长春王春雨马均
李娜,杨志远,代邹,孙永健,徐徽,何艳,严田蓉,蒋明金,郭长春,王春雨,马均
不同氮效率水稻根系形态和氮素吸收利用与产量的关系
李娜,杨志远,代邹,孙永健,徐徽,何艳,严田蓉,蒋明金,郭长春,王春雨,马均
(四川农业大学水稻研究所/农业部西南作物生理生态与耕作重点实验室,成都 611130)
【目的】探究不同氮效率水稻根系形态和氮素吸收利用与产量关系的规律,明确水稻高产根系形态特征,提出相应的水氮优化管理措施。【方法】2013年以氮高效品种川农优498和氮低效品种川优6203为试验材料,进行不同穗肥运筹比例的裂区试验;2014年以相同试验材料进行穴苗数和促花肥、保花肥配比的裂裂区试验;2015年以氮高效品种德香4103和氮低效品种宜香3724为试验材料,进行水分管理方式和氮肥施用模式的裂裂区试验;分别以上述3个大田试验获得的产量、根系形态及氮素吸收利用相关指标为样本,通过计算方差膨胀因子诊断变量间存在的多重共线性关系。在多重共线性严重的情况下,运用岭回归分析研究根系形态、氮素吸收利用和产量三者之间的关系。基于这些关系在年度间的重演性,确定其中普遍存在的共性规律。【结果】(1)氮高效品种拔节期、抽穗期及成熟期的氮素积累量、氮素干物质生产效率和干物重与产量岭回归方程的决定系数范围分别为0.0219—0.3961、0.0452—0.1379和0.0914—0.6694,氮低效品种分别为0.0084—0.6190、0.1224—0.4341和0.0818—0.4881,产量与氮素吸收利用的关系年度间重演性较差,无明显共性规律;以根干重、不定根数量、长度、表面积、体积、粗分枝根长度、表面积、体积以及细分枝根长度、表面积、体积11项根系形态指标为自变量,氮素积累量、氮素干物质生产效率和干物重三者分别为因变量进行岭回归分析,氮高效品种的回归方程决定系数范围分别为0.0527—0.2728、0.0653—0.3139和0.0714—0.3158,氮低效品种分别为0.0607—0.5040、0.0555—0.4411和0.0724—0.5449,氮素吸收利用与根系形态的关系规律年度间存在较大差异;2013—2015年,氮高效和氮低效品种抽穗期根系形态与产量岭回归方程的决定系数均超过0.8,<0.001,表明抽穗期根系形态对产量具有显著影响是共性规律。(2)在以抽穗期根系形态指标为自变量,产量为因变量的岭回归方程中,氮高效品种的粗分枝根长度标准回归系数最高,对产量影响最大;对氮低效品种而言,细分枝根表面积的增加对产量提高最有利。(3)常规施氮量(150 kg·hm-2)下,优化施肥模式为穗肥占比40%,且保花肥的比例达到或超过50%,其结合交替灌溉,有利于优化根系形态;SPAD指导施肥模式下,施氮量较优化施肥减少(由150 kg·hm-2降为120 kg·hm-2),采用常规灌溉比交替灌溉更有利于根系形态优化。【结论】水稻抽穗期根系形态与产量关系极为密切,合理的水氮管理措施能够优化根系形态提高产量。常规灌溉结合SPAD指导施肥或交替灌溉结合优化施肥均有利于氮高效品种抽穗期粗分枝根长度的增长和氮低效品种细分枝根表面积的增大,最终提高产量。
氮效率;水稻;根系形态;产量;岭回归分析
0 引言
【研究意义】水稻根系是土壤养分的直接利用者和地上部光合生产主要的养分供应者,其形态和生理特征与地上部生长发育和产量形成有密切关系。氮素是水稻生长发育最重要的元素之一,水稻体内氮素主要由根系从土壤中吸收[1-2]。分别以根系和产量为出发点和落脚点,以氮素吸收利用为纽带,进行水稻根系性状和氮素吸收利用与产量关系的定量研究,有助于深入剖析根系在水稻产量形成过程中发挥作用的机理。【前人研究进展】围绕水稻根系与氮素吸收和利用的关系,及其对产量形成的影响,前人已经开展了比较系统的研究,从水稻根系对氮素吸收利用的基因型差异[3-7],到根系对氮肥调控的响应[8-10],再到农艺措施对根系的影响及其与地上部生长及产量形成的关系[11-12]等方面均有涉及,这些研究在根系形态特征、生理活性、基因表达等层面取得了丰硕的理论成果,同时针对性地提出了一系列栽培技术措施,促进了水稻氮肥利用效率和产量的同步提高。【本研究切入点】已有研究多以包含较少变量的环境控制性重复试验为样本,通过简单相关分析或普通最小二乘法回归分析,获得水稻根系形态和氮素吸收利用与产量的关系。这种研究策略针对性较强,得到的结果对特定条件下水稻生产的指导价值较大,但由于重复试验工作量大,故这类研究包含的控制变量普遍较少,导致最终的试验结果存在理论局限性较大,技术可移植性不高的问题。同时在统计分析中由于未对研究对象内部(自变量间)的共线性问题予以足够重视,使得试验结果中变量间关系的稳定性较差,加剧了不同研究结果的不一致性[6-7, 13]。针对上述研究不足,拟从不同试验组成的包含较多变量因子的样本中探求水稻根系形态、氮素吸收利用和产量三者之间的关系,与少变量重复试验样本相比,从多变量非重复试验样本中发掘三者关系的共性规律难度更大,但若三者的关系在排除共线性的干扰后仍能够呈现出共性规律,则该研究结果的生产指导价值将更加广泛。【拟解决的关键问题】本研究以课题组前期筛选出的在四川盆地稻区广泛种植的氮高效和氮低效品种为试验材料[14-16],通过3年大田试验,探究水稻根系形态、氮素吸收和利用、产量形成三者之间关系的规律,以及实现高产的根系特征及相应水氮调控措施,为水稻高产高效育种及栽培提供理论依据及技术参考。
1 材料与方法
1.1 试验地点
试验于2013—2015年在四川农业大学水稻研究所试验农场(30°43’N,103°47’E)进行。试验点地处成都平原,属亚热带湿润气候区。2006—2015年水稻生长季气象资料来自四川省气象局(图1)。试验地前茬为油菜,土壤质地为砂壤土,耕层土壤养分含量见表1。
1.2 供试材料
本研究以课题组前期筛选出的在农学利用率上有显著差异的品种为供试材料。2013年和2014年为氮高效品种川农优498和氮低效品种川优6203[16],2015年为氮高效品种德香4103和氮低效品种宜香3724[14-15]。4个品种均为中籼迟熟杂交稻,生育期150 d左右。

图1 2006—2015年水稻生长季平均气温和降雨量

表1 2013—2015年耕层土壤养分含量表
1.3 试验设计
2013年试验采用裂区设计,品种为主区,氮肥运筹为副区,设基蘖肥与穗肥配比为B1(90﹕10)、B2(75﹕25)、B3(60﹕40)、B4(45﹕55)、B5(30﹕70),共计10个处理,3次重复,施氮量为150 kg·hm-2。
2014年试验为裂裂区设计,品种为主区,每穴苗数为副区,设单苗(S)和双苗(D)2个水平,穗肥氮运筹为裂裂区,设促花肥与保花肥配比为P1(40﹕0)、P2(20﹕20)、P3(0﹕40),共计12个处理,3次重复,施氮量为150 kg·hm-2,其中穗肥占比40%。
2015年试验为裂裂区设计,品种为主区,水分管理为副区,设常规灌溉(W1)和交替灌溉(W2)2个水平[17],氮肥管理为裂裂区,设SPAD指导施肥(N1)、优化施肥(N2)及农民习惯施肥(N3)3种模式(表2),共计12个处理,3次重复。
2013—2015年试验中,基肥(包含75 kg·hm-2磷肥和150 kg·hm-2钾肥)、蘖肥、促花肥和保花肥分别于移栽前1 d、移栽后7 d、幼穗分化期(倒四叶)和抽穗前(倒二叶)施用。
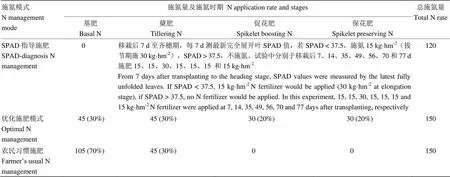
表2 2015年氮肥施用量及施肥时期
括号内百分数代表该日期氮肥施入量占总施氮量的比例 Values in the brackets indicate the percentage of N fertilizer applied to total N rate
1.4 测定项目与方法
1.4.1 根系形态指标测定 分别于拔节期、抽穗期和成熟期,每小区按平均茎蘖数标记5株具有代表性稻株,采用原状土柱法,用铁板取根器以稻株为中心掘取长等于行距(33.3 cm),宽等于株距(16.7 cm),深20 cm的土柱,装入孔径为0.4 mm的尼龙网袋中,浸泡6 h后洗去泥土杂质,获得单株完整根系。用Epson Expression 10000XL扫描后,采用WinRHIZO Prov.2009c软件分析不定根、粗分枝根、细分枝根各形态参数,而后置于80℃下烘干至恒重,测得根系总干重。不定根(0.3 mm<D≤1.65 mm)、粗分枝根(0.1 mm<D≤0.3 mm)、细分枝根(D≤0.1 mm)的界定参照顾东祥等[18]的方法。
1.4.2 氮素积累测定 将根系形态指标测定剩余的地上部,分茎、叶、穗各器官烘干,称质量,再粉碎、过筛,用凯氏定氮仪(FOSS—8400)测定各器官的全氮含量。氮素干物质生产效率(nitrogen dry matter production efficiency,NDMPE)用单位氮素生产的干物质量表示。
1.4.3 考种与计产 成熟期各小区单独收割,按实收株数计产。
1.5 数据处理
方差膨胀因子(variance inflation factor,VIF)用于诊断变量间存在的共线性关系,当某组变量的VIF平均值大于1且最大值大于10时,认为该组变量共线性严重。本研究用SPSS 20计算自变量的方差膨胀因子,结果显示自变量间共线性严重(表3),符合进行岭回归分析的条件。岭回归分析(ridge regression)是一种用于共线性数据分析的有偏估计回归,通过在自变量信息矩阵的主对角线元素上加入一个非负因子(岭回归参数k),使回归系数的估计稍有偏差,但估计的稳定性明显提高。本研究中当k=0.2时各自变量的岭迹都基本稳定,因此岭参数都取k=0.2。本研究用DPS 7.05进行岭回归分析和多重比较,用Excel 2003进行图表制作。
2 结果
2.1 根系形态与氮素吸收利用及物质积累的岭回归分析
氮高效品种的产量、氮积累量、氮素干物质生产效率及成熟期干物重均显著高于氮低效品种(表4)。二者根系形态与氮素吸收利用及物质积累的岭回归分析显示,54组样本中仅有2组决定系数超过0.5。2015年氮低效品种在拔节期的根系形态对氮素积累量和干物重变化的影响程度分别达到0.5040和0.5449,但在2013年和2014年对应样本的方程决定系数均不超过0.2(表5)。这表明水稻根系形态与氮素积累量、氮素干物质生产效率及干物重关系的规律性较差。

表3 自变量方差膨胀因子
HN:氮高效;LN:氮低效;JS:拔节期;HS:抽穗期;MS:成熟期;NA:氮素积累;NDMPE:氮素干物质生产效率;DW:干物质量。下同
HN: High N use efficiency; LN: Low N use efficiency; JS: Jointing stage; HS: Heading stage; MS: Maturity stage; NA: N accumulation; NDMPE: N dry matter production efficiency; DW: Dry matter weight. The same as below

表4 氮高效和氮低效品种产量和氮素吸收利用比较
同列数据后不同字母表示在5 %水平差异显著。下同
Values within a column followed by different letters are significantly different at<0.05. The same as below

表5 根系形态与氮素吸收利用及物质积累的岭回归分析
*, **, ***分别表示在0.05, 0.01, 0.001水平差异显著;NS表示二者差异未达显著水平。下同
*, ** and *** indicate significance at 0.05, 0.01 and 0.001 levels, respectively; NS means that the difference was not significant. The same as below
2.2 各生育时期氮素吸收利用及物质积累与产量的岭回归分析
表6表明,氮高效和氮低效水稻在拔节期、抽穗期和成熟期的氮素积累量、氮素干物质生产效率及干物重与产量岭回归分析方程的决定系数大多低于0.5,仅有的决定系数超过0.6的3组(共计18组)样本在年份间或品种间的重演性均较差。2013年和2014年氮高效品种成熟期的氮素积累量、氮素干物质生产效率及干物重对产量的影响程度分别为0.6694和0.6596,其中成熟期干物重对产量影响最大,氮素干物质生产效率次之,氮素积累量最小,但2015年岭回归方程的决定系数仅为0.0914,且干物重对产量的影响远小于氮素干物质生产效率和氮素积累量。2015年氮低效品种在拔节期的氮素积累量、氮素干物质生产效率及干物重对产量的影响程度达到0.6190,但2013年和2014年岭回归方程的决定系数仅为0.0084和0.0680。这表明水稻拔节期、抽穗期及成熟期的氮素吸收、利用及生物量对产量的影响可能具有较强的品种特异性或受气象条件影响较大,因而相互关系的规律重演性较差。
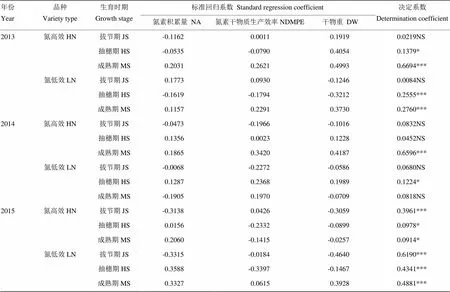
表6 产量与各生育时期氮素吸收利用及物质积累的岭回归分析
2.3 抽穗期根系形态与产量的岭回归分析
以水稻拔节期、抽穗期及成熟期的根系形态为自变量与产量作岭回归分析,结果显示不同氮效率水稻均表现为抽穗期根系形态结构特征与产量关系的密切程度远大于拔节期、成熟期,且该结果在3年大田试验中重演性较好(数据未列出),本文仅对抽穗期根系形态与产量的关系进行分析(表7)。6组样本的方程决定系数均超过0.8,达到极显著水平,2013年和2014年氮高效品种抽穗期根系形态结构对产量的解释程度均高于氮低效品种,2015年二者岭回归方程的决定系数相近。从各根系形态指标分析,氮高效品种的粗分枝根长度与产量关系最密切,氮低效品种的细分枝根表面积对产量的影响居所有指标之首。

表7 抽穗期根系形态与产量的岭回归分析
TRW:根干重。下同 TRW: Total root dry weight. The same as below
2.4 水肥管理对抽穗期根系形态的影响
2.4.1 氮肥运筹对抽穗期根系形态的影响 由表8可见,氮高效水稻抽穗期根系形态指标较氮低效水稻平均高21.81%—23.08%。随着穗肥占比升高,根系生物量呈逐渐增大趋势,穗肥占比超过40%,根系干重变化较小。穗肥占比提高促进了不定根数量、长度、表面积及体积的增长,穗肥占比超过40%,氮高效品种不定根数量及表面积等指标呈小幅下降趋势,氮低效品种基本保持稳定。不定根上分出的粗分枝根以及后者分出的细分枝根也延续了不定根对氮肥运筹的响应规律,大都在B3处理时达到最大值。
2.4.2 穗肥氮运筹及每穴苗数对抽穗期根系形态的影响 氮高效品种抽穗期根系指标较氮低效品种平均高21.76%—22.18%,单苗或双苗栽插对根系形态影响较小(表9)。施用保花肥能显著提高根系生物量,但不同保花肥施用比例间差异较小。不定根的数量、长度、表面积和体积及其上分出的粗分枝根和细分枝根的各项指标也基本符合根系干重对穗肥运筹的响应规律。施用保花肥使氮高效品种粗分枝根长度和氮低效品种细分枝根表面积分别增加5.21%—20.78%和12.19 %—16.74 %。
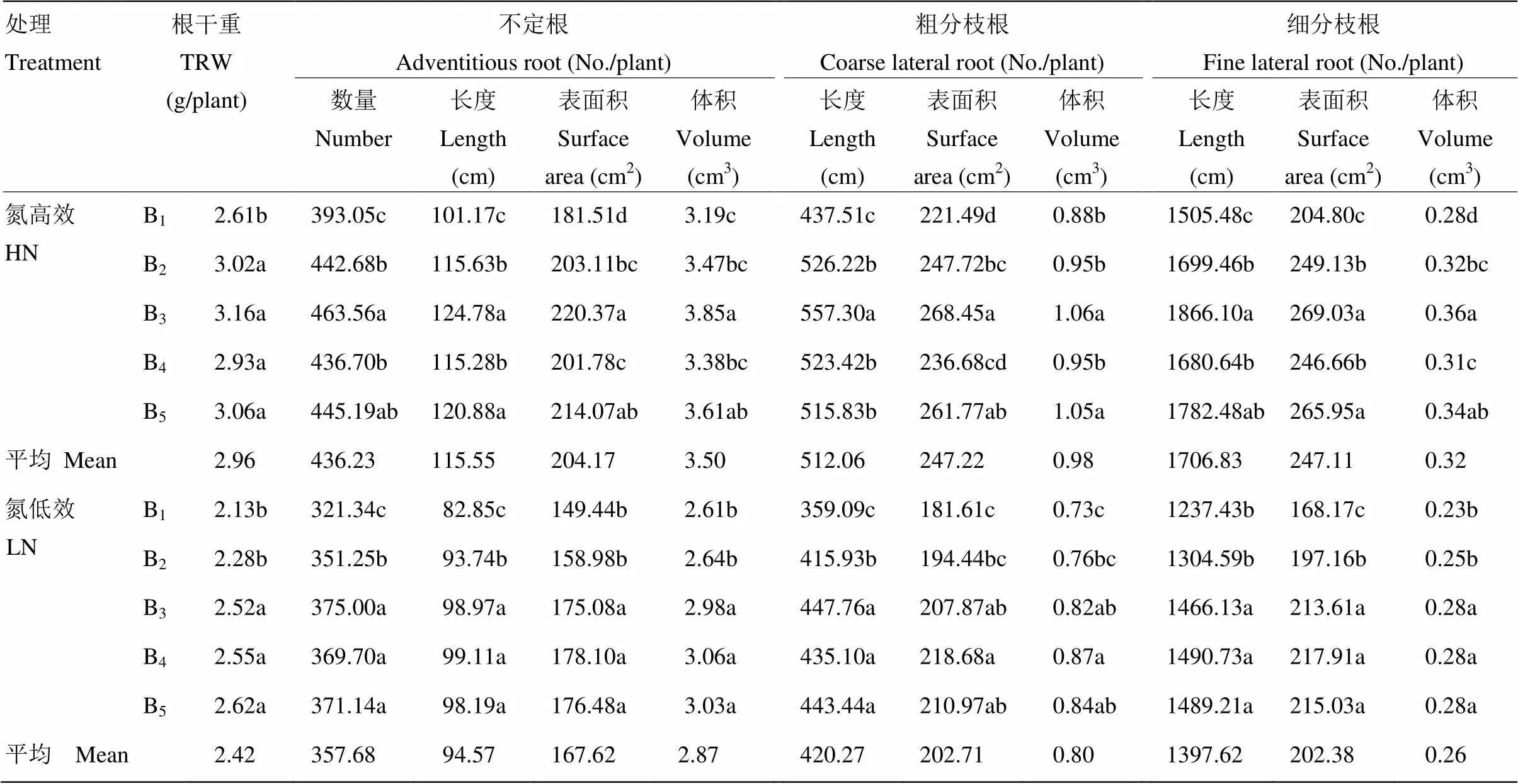
表8 氮肥运筹对抽穗期根系形态的影响(2013年)
B1:基蘖肥﹕穗肥=90 : 10;B2:基蘖肥﹕穗肥=75 : 25;B3:基蘖肥﹕穗肥=60 : 40;B4:基蘖肥﹕穗肥=45 : 55;B5:基蘖肥﹕穗肥=30 : 70
B1: Basal - tillering N : panicle N = 90 : 10; B2: Basal - tillering N : panicle N = 75 : 25; B3: Basal - tillering N : panicle N = 60 : 40; B4: Basal - tillering N : panicle N =45 : 55; B5: Basal - tillering N : panicle N = 30 : 70

表9 穗肥氮运筹及每穴苗数对抽穗期根系形态的影响(2014年)
S:单苗;D:双苗;P1:促花肥﹕保花肥=40 : 0;P2:促花肥﹕保花肥=20 : 20;P3:促花肥﹕保花肥=0 : 40
S: Single seedling; D: Double seedlings; P1: The ratio of nitrogen dressing at 4thand 2ndleaf from top stretching was 40 : 0; P2: The ratio of nitrogen dressing at 4thand 2ndleaf from top stretching was 20 : 20; P3: The ratio of nitrogen dressing at 4thand 2ndleaf from top stretching was 0 : 40
2.4.3 水氮管理对抽穗期根系形态的影响 表10表明,无论在常规灌溉还是交替灌溉条件下,均以农民习惯施肥(N3)抽穗期根系形态指标最小。常规灌溉条件下SPAD指导施肥氮高效品种根系形态指标均显著高于优化施肥,采用SPAD指导施肥或优化施肥对氮低效品种根系形态指标影响差异较小。交替灌溉条件下,氮高效品种优化施肥比SPAD指导施肥在根系形态指标上更具优势,而氮低效品种采用SPAD指导施肥更有利根系生长发育。对氮高效品种而言,常规灌溉下采用SPAD指导施肥和交替灌溉下采用优化施肥对粗分枝根长度增长更有利,比农民习惯施肥分别提高35.87%和36.26%;对氮低效品种而言,常规灌溉下SPAD指导施肥或交替灌溉下优化施肥均能有效促进细分枝根表面积增加,较农民习惯施肥分别提高29.91%和28.98%。

表10 水氮管理对抽穗期根系形态的影响(2015年)
W1:常规灌溉,W2:交替灌溉;N1、N2、N3分别代表SPAD指导施肥、优化施肥模式、农民习惯施肥
W1: Conventional irrigation, W2: Controlled alternate irrigation; N1: SPAD-diagnosis N management; N2: Optimal N management; N3: Farmer’s usual N management
3 讨论
3.1 根系形态与氮素吸收利用的关系
Cassman等[6]研究表明氮高效水稻在根系生物量、体积、总吸收表面积和活性吸收面积方面均具有较大优势。戢林等[3]研究认为水稻粗分枝根的发育情况会直接影响氮素的吸收,进而影响产量和氮素利用效率。Samejima等[7, 19]和石庆华等[13]研究认为部分在根系形态、生理活性方面较优的品种,在氮素吸收利用率方面并未表现明显优势。本研究水稻根系形态与氮素吸收利用岭回归方程的决定系数波动范围较大(0.0527≤2≤0.5449),且相同指标在不同试验中表现也差异较大,这反映了水稻根系形态与氮素吸收利用关系的复杂性。首先,水稻根系吸收养分的过程中,氮、磷、钾等元素存在显著地协同或拮抗作用,而磷等对根系生长发育具有较强的调节作用[20-22];其次,根系吸收养分的同时会以根泌的方式释放出不同类型化合物,反馈调节根系生长发育及对氮、磷、钾等的吸收利用[23];因此土壤性状、肥水管理等都可能对根系形态与氮素吸收利用的关系产生重要影响。本研究中,不同试验间的气象条件、土壤性状及水氮管理等均存在一定差异,这可能是根系性状与氮素吸收利用之间关系在3个试验中无共性规律的主要原因。
3.2 氮素吸收利用及物质积累与产量的关系
氮素干物质生产效率是衡量水稻氮素利用效率的重要指标之一[24],过量施氮或穗肥占比过高易导致植株氮积累尤其是花后氮积累偏高,削弱其碳同化能力,使氮素干物质生产效率降低[25]。Chen等[26]研究认为不同氮效率水稻的氮素籽粒生产效率及氮素干物质生产效率差异并不显著,氮高效品种发挥产量优势的关键在于吸收更多的氮素。Qiao等[27]研究认为在高地力稻田,施氮虽能增加植株氮积累量,但较多氮素会滞留在秸秆内,增产效果较差。本研究显示在决定系数较高的岭回归方程里,氮素吸收和利用对产量的影响较小,普遍低于生物量,与前人研究存在一定差异,其原因可能在两方面:首先,四川盆地稻田土壤基础肥力高,对产量贡献大,属地力贡献率高值区,施肥发挥的作用较小[28-29]。其次,四川盆地昼夜温差小、光照时数少,水稻产量潜力小。本研究中绝大多数处理产量已经超过该地区产量潜力估值,此时氮素可能已经不是产量的决定因素[30-31]。
2013年和2014年试验氮高效品种干物重对产量的影响远大于2015年试验,除了品种间的基因型差异外,气象条件的差异亦可能是重要原因。2015年试验水稻灌浆期降水频繁,日照时数较近十年平均值减少27%,较2013年和2014年分别减少35%和29%,光照不足限制了水稻群体花后光合生产能力,增强了花前物质输出对产量的影响,导致生物量对产量的影响减弱[32]。
3.3 抽穗期根系形态与产量的关系
目前研究普遍认为抽穗期水稻根系已经完成生长且对产量影响最大[33-34]。本研究显示抽穗期根系形态对产量的影响在所有样本中均较大,表明该时期根系形态与产量的密切关系具有普遍性。就常见形态指标来说,不定根数量、根系吸收表面积及根直径等对产量影响较大,采用根系分析系统细化到分枝根层面,则是粗分枝根长度、表面积及细分枝根表面积等与产量关系密切[3, 35]。本研究获得相近的结果,即粗分枝根长度和细分枝根表面积分别对氮高效和氮低效品种产量影响较大。不同氮效率品种间的差异可能与二者对养分的需求差异有关,氮高效品种更高产量的实现需要较多的养分支撑,粗分枝根数量增加有利于扩展根系吸收空间,增加土壤养分供应量[3]。
根系对土壤环境的适应具有可塑性,合理的水氮管理能够通过与土壤互作优化根系形态提高产量,在此过程中分枝根的表现尤为突出[10,18,36-38]。本研究中,合理施氮(基蘖肥﹕促花肥﹕保花肥= 60﹕20﹕20)和水氮耦合管理(SPAD指导施肥采用常规灌溉、优化施肥结合交替灌溉)能够优化抽穗期氮高效品种粗分枝根长度和氮低效品种细分枝根表面积,实现产量增长。
4 结论
水稻根系形态对产量具有重要影响,尤其是抽穗期根系形态与产量关系极为密切。合理施氮与控水可优化各类型根的生长与分布特征,但需考虑不同品种之间的差异,促进抽穗期粗分枝根长度及细分枝根表面积增长分别是实现氮高效和氮低效品种根系形态优化的首要目标。常规施氮量(150 kg·hm-2)下,穗肥比例为40%,且其中保花肥的占比达到或超过50%,同时结合交替灌溉,有利于优化根系形态;在SPAD指导施肥模式(施氮量为120 kg·hm-2)下,采用常规灌溉更有利于根系形态优化。
References
[1] PENG S B, HUANG J L, ZHONG X H, YANG J C, WANG G H, ZOU Y B, ZHANG F S, ZHU Q S, ROLAND B, CHRISTIAN W.Challenge and opportunity in improving fertilizer-nitrogen use efficiency of irrigated rice in China., 2002, 1(7): 776-785.
[2] 蔡昆争, 骆世明, 段舜山. 水稻根系在根袋处理条件下对氮养分的反应. 生态学报, 2003, 23(6): 1109-1116.
CAI K Z, LUO S M, DUAN S S. The response of the rice root system to nitrogen conditions under root confinement., 2003, 23(6): 1109-1116. (in Chinese)
[3] 戢林, 李廷轩, 张锡洲, 余海英. 氮高效利用基因型水稻根系形态和活力特征. 中国农业科学, 2012, 45(23): 4770-4781.
JI L, LI T X, ZHANG X Z, YU H Y. Root morphologyical and activity characteristics of rice genotype with high nitrogen utilization efficiency.2012, 45(23): 4770-4781. (in Chinese)
[4] 魏海燕, 张洪程, 张胜飞, 杭杰, 戴其根, 霍中洋, 许轲, 马群, 张庆, 刘艳阳. 不同氮利用效率水稻基因型的根系形态与生理指标的研究. 作物学报, 2008, 34(3): 429-436.
WEI H Y, ZHANG H C, ZHANG S F, HANG J, DAI Q G, HUO Z Y, XU K, MA Q, ZHANG Q, LIU Y Y.Root morphological and physiological characteristics in rice genotypes with different N use efficiencies.2008, 34(3): 429-436. (in Chinese)
[5] 李敏, 张洪程, 杨雄, 葛梦婕, 马群, 魏海燕, 戴其根, 霍中洋, 许轲, 曹利强, 吴浩. 水稻高产氮高效型品种的根系形态生理特征. 作物学报, 2012, 38(4): 648-656.
LI M, ZHANG H C, YANG X, GE M J, MA Q, WEI H Y, DAI Q G, HUO Z Y, XU K, CAO L Q, WU H. Root morphological and physiological characteristics of rice cultivars with high yield and high nitrogen use efficiency.2012, 38(4): 648-656. (in Chinese)
[6] CASSMAN K G, DOBERMANN A, WALTERS D T. Agroecosystems, nitrogen-use efficiency, and nitrogen management.,2002, 31(2): 132-140.
[7] SAMEJIMA H, KONDO M, ITO O, NOZOE T, SHINANO T, OSAKI M. Characterization of root systems with respect to morphological traits and nitrogen-absorbing ability in the new plant type of tropical rice lines.2005, 28(5): 835-850.
[8] 樊小林, 史正军, 吴平. 水肥(氮)对水稻根构型参数的影响及其基因型差异. 西北农林科技大学学报(自然科学版), 2002, 30(2): 1-5.
FAN X L, SHI Z J, WU P.Effects of nitrogen fertilizer on parameters of rice (L.) root architecture and their genotypic difference., 2002, 30(2): 1-5. (in Chinese)
[9] 孙虎威, 王文亮, 刘尚俊, 候蒙蒙, 谢天宁, 粱志浩, 樊亚男, 张亚丽. 低氮胁迫下水稻根系的发生及生长素的响应. 土壤学报, 2014, 51(5): 1096-1102.
SUN H W, WANG W L, LIU S J, HOU M M, XIE T N, LIANG Z H, FAN Y N, ZHANG Y L. Formation of rice root regulated by nitrogen dificiency.2014, 51(5): 1096-1102. (in Chinese)
[10] WALCH-LIU P, IVANOV I, FILLEUR S Y, GAN Y, REMANS T, FORDE B G. Nitrogen regulation of root branching.2006, 97(5): 875-881.
[11] 杨志远, 孙永健, 徐徽, 秦俭, 贾现文, 马均. 栽培方式与免耕对杂交稻Ⅱ优498灌浆期根系衰老和籽粒灌浆的影响. 中国农业科学, 2013, 46(7): 1347-1358.
YANG Z Y, SUN Y J, XU H, QIN J, JIA X W, MA J. Influence of cultivation methods and no-tillage on root senescence at filling stage and grain-filling properties of Eryou 498., 2013, 46(7): 1347-1358. (in Chinese)
[12] 张洪程, 朱聪聪, 霍中洋, 许轲, 蒋晓鸿, 陈厚存, 高尚勤, 李德剑, 赵成美, 戴其根, 魏海燕, 郭保卫. 钵苗机插水稻产量形成优势及主要生理生态特点. 农业工程学报, 2013, 29(21): 50-59.
ZHANG H C, ZHU C C, HUO Z Y, XU K, JIANG X H, CHEN H C, GAO S Q, LI D J, ZHAO C M, DAI Q G, WEI H Y, GUO B W.Advantages of yield formation and main characteristics of physiological and ecological in rice with nutrition bowl mechanical transplanting.2013, 29(21): 50-59. (in Chinese)
[13] 石庆华, 李木英, 涂起红. 杂交水稻根系N素营养效率及其生理因素研究. 杂交水稻, 2002, 17(4): 49-52.
SHI Q H, LI M Y, TU Q H. Studies on efficiency of N nutrition and physiological factors in roots of hybrid rice.2002, 17(4): 49-52. (in Chinese)
[14] 秦俭, 杨志远, 孙永健, 马均. 不同穗型杂交籼稻物质积累、氮素吸收利用和产量的差异比较. 中国水稻科学, 2014, 28(5): 514-522.
QIN J, YANG Z Y, SUN Y J, MA J. Differential comparison of assimilation products accumulation, nitrogen uptake and utilization and grain yield of hybridrice combinations with different panicle types.,2014, 28(5): 514-522. (in Chinese)
[15] 朱从桦, 孙永健, 严奉君, 蒋明金, 徐徽, 赵建红, 马均. 晒田强度和氮素穗肥运筹对不同氮效率杂交稻产量及氮素利用的影响. 中国水稻科学, 2014, 28(3): 258-266.
ZHU C H, SUN Y J, YAN F J, JIANG M J, XU H, ZHAO J H, MA J. Effects of different paddy field drainage degrees and panicle nitrogen fertilizer managements on yield and nitrogen utilization of different nitrogen efficiency hybrid rice.2014, 28(3): 258-266. (in Chinese)
[16] 杨世民, 杨志远, 孙永健, 马均. 氮肥运筹对2个不同穗重型杂交稻产量及氮肥利用的影响. 浙江大学学报(农业与生命科学版), 2015, 41(6): 685-694.
YANG S M, YANG Z Y, SUN Y J, MA J. Effects of nitrogen on grain yield and nitrogen use efficiency of two hybrid rice varieties with different panicle masses.2015, 41(6): 685-694. (in Chinese)
[17] 孙永健, 马均, 孙园园, 徐徽, 严奉君, 代邹, 蒋明金, 李玥. 水氮管理模式对杂交籼稻冈优527群体质量和产量的影响. 中国农业科学, 2014, 47(10): 2047-2061.
SUN Y J, MA J, SUN Y Y, XU H, YAN F J, DAI Z, JIANG M J, LI Y. Effects of water and nitrogen management patterns on population quality and yield of hybrid rice Gangyou 527.,2014, 47(10): 2047-2061. (in Chinese)
[18] 顾东祥, 汤亮, 徐其军, 雷晓俊, 曹卫星, 朱艳. 水氮处理下不同品种水稻根系生长分布特征. 植物生态学报, 2011, 35(5): 558-566.
GU D X, TANG L, XU Q J, LEI X J, CAO W X, ZHU Y. Root growth and distribution in rice cultivars as affected by nitrogen and water supply.2011, 35(5): 558-566. (in Chinese)
[19] SAMEJIMA H, KONDO M, ITO O, NOZOE T, SHINANO T. Root-shoot interaction as a limiting factor of biomass productivity in new tropical rice lines.2004, 50(4): 545-554.
[20] 许飞云, 张茂星, 曾后清, 朱毅勇. 水稻根系细胞膜质子泵在氮磷钾养分吸收中的作用. 中国水稻科学, 2016, 30(1): 106-110.
XU F Y, ZHANG M X, ZENG H Q, ZHU Y Y. Involvement of plasma membrane H+-ATPase in uptake of nitrogen, phosphorus and potassium by rice root.2016, 30(1): 106-110. (in Chinese)
[21] DINGKUHN M, LUQUET D, KIM H K, TAMBOUR L, CLEMENTVIDAL A. EcoMeristem, a model of morphogenesis and competition among sinks in rice. 2. Simulating genotype responses to phosphorus deficiency.,2006, 33(4): 325-337.
[22] INSALUD N, BELL R W, COLMER T D, RERKASEM B. Morphological and physiological responses of rice () to limited phosphorus supply in aerated and stagnant solution culture.2006, 98(5): 995-1004.
[23] LI M, ZHANG H C, YANG X, GE M J, MA Q, WEI H Y, DAI Q G, HUO Z Y, XU K, LUO D Q. Accumulation and utilization of nitrogen, phosphorus and potassium of irrigated rice cultivars with high productivities and high N use efficiencies.2014, 161: 55-63.
[24] CASSMAN K G, Peng S, OLK D C, LADHA J K, REICHARDT W, DOBERMANN A, SINGH U. Opportunities for increased nitrogen- use efficiency from improved resource management in irrigated rice systems.1998, 56(1): 7-39.
[25] JIANG L G, DAI T B, JIANG D, CAO W X, GAN X Q, WEI S Q. Characterizing physiological N-use efficiency as influenced by nitrogen management in three rice cultivars.2004, 88(2/3): 239-250.
[26] CHEN G, CHEN Y, ZHAO G H, CHENG W D, GUO S W, ZHANG H L, SHI W M. Do high nitrogen use efficiency rice cultivars reduce nitrogen losses from paddy fields?2015, 209: 26-33.
[27] QIAO J, YANG L Z, YAN T M, XUE F, ZHAO D. Nitrogen fertilizer reduction in rice production for two consecutive years in the Taihu Lake area.2012, 146(1): 103-112.
[28] 汤勇华, 黄耀. 中国大陆主要粮食作物地力贡献率和基础产量的空间分布特征. 农业环境科学学报. 2009, 28(5): 1070-1078.
TANG Y H, HUANG Y. Spatial distribution characteristics of the percentage of soil fertility contribution and its associated basic crop yield in Mainland China., 2009, 28(5): 1070-1078. (in Chinese)
[29] 梁涛, 陈轩敬, 赵亚南, 黄兴成, 李鸿, 石孝均, 张跃强. 四川盆地水稻产量对基础地力与施肥的响应. 中国农业科学, 2015, 48(23): 4759-4768.
LIANG T, CHEN X J, ZHAO Y N, HUANG X C, LI H, SHI X J, ZHANG Y Q. Response of rice yield to inherent soil productivity of paddies and fertilization in Sichuan basin, 2015, 48(23): 4759-4768. (in Chinese)
[30] VAN WART J, KERSEBAUM K C, Peng S B, MILNER M, CASSMAN K G. Estimating crop yield potential at regional to national scales.2013, 143(1): 34-43.
[31] DOBERMANN A. Fertilizer best management practices: General principles, strategy for their adoption and voluntary initiatives vs regulations//Proceedings of the IFA International Workshop on Fertilizer Best Management Practices. Paris: International Fertilizer Industry Association, 2007: 1-28.
[32] 杨志远, 胡蓉, 孙永健, 徐徽, 许远明, 马均. 三角形强化栽培模式下氮肥运筹对II优498产量及氮肥利用的影响. 作物学报, 2012, 38(6): 1097-1106.
YANG Z Y, HU R, SUN Y J, XU H,XU Y M, MA J. Effects of nitrogen fertilizer management on yield and nitrogen use efficiency of Eryou 498 in triangle-planted system of rice intensification., 2012, 38(6): 1097-1106. (in Chinese)
[33] 刘桃菊, 戚昌瀚, 唐建军. 水稻根系建成与产量及其构成关系的研究. 中国农业科学, 2002, 35(11): 1416-1419.
LIU T J, QI C H, TANG J J. Studies on relationship between the character parameters of root and yield formation in rice.2002, 35(11): 1416-1419. (in Chinese)
[34] 樊剑波, 沈其荣, 谭炯壮, 叶利庭, 宋文静, 张亚丽. 不同氮效率品种根系生理生态指标的差异. 生态学报, 2009, 29(6): 3052-3058.
FAN J B, SHEN Q R, TAN J Z, YE L T, SONG W J, ZHANG Y L. Difference of root physiological and ecological indicies in rice cultivars with different N use efficiency.2009, 29(6): 3052-3058. (in Chinese)
[35] 李杰, 张洪程, 常勇, 龚金龙, 胡雅杰, 龙厚元, 戴其根, 霍中洋, 许轲, 魏海燕, 高辉. 高产栽培条件下种植方式对超级稻根系形态生理特征的影响. 作物学报, 2011, 37(12): 2208-2220.
LI J, ZHANG H C, CHANG Y, GONG J L, HU Y J, LONG H Y, DAI Q G, HUO Z Y, XU K, WEI H Y, GAO H. Influence of planting methods on root system morphological and physiological characteristics of super rice under high-yielding cultivation condition., 2011, 37(12): 2208-2220. (in Chinese)
[36] DE DORLODOT S, FORSTER B, PAGÈS L, PRICE A, TUBEROSA R, DRAYE X. Root system architecture: Opportunities and constraints for genetic improvement of crops.2007, 12(10): 474-481.
[37] DAVIES W J. Root growth response and functioning as an adaptation in water limiting soils//JENKS M A, HASEGAWA P M, JAIN S M.. Dordrecht: Springer Netherlands, 2007: 55-72.
[38] 徐国伟, 吕强, 陆大克, 王贺正, 陈明灿. 干湿交替灌溉耦合施氮对水稻根系性状及籽粒库活性的影响. 作物学报, 2016, 42(10): 1495-1505.
XU G W, Lü Q, LU D K, WANG H Z, CHEN M C. Effect of wetting and drying alternative irrigation coupling with nitrogen application on root characteristic and grain-sink activity.2016, 42(10): 1495-1505. (in Chinese)
(责任编辑 杨鑫浩)
The Relationships Between Root Morphology, N Absorption and Utilization and Grain Yield in Rice with Different N Use Efficiencies
LI Na, YANG ZhiYuan, DAI Zou, SUN YongJian, XU Hui, HE Yan, YAN TianRong, JIANG MingJin, GUO ChangChun, WANG ChunYu, MA Jun
(Rice Research Institute, Sichuan Agricultural University/Key Laboratory of Crop Physiology, Ecology, and Cultivation in Southwest, Ministry of Agriculture, Chengdu 611130)
【Objective】The objective of this research is to study the relationships between root morphology, nitrogen (N) absorption and utilization and grain yield in rice with different N use efficiencies, clarify the root morphology characters of high yielding, and put forward the corresponding optimized water-N management practice. 【Method】In 2013, high N use efficiency (HN) variety Chuannongyou 498 and low N use efficiency (LN) variety Chuanyou 6203 were used as trial materials to conduct a split-plot experiment with different panicle fertilizer ratio. In 2014, a split-split plot experiment was performed with the same varieties, where seedling number per hole was assigned to the split plot, and the proportion of spikelet preserving fertilizer and spikelet promoting fertilizer was assigned to the split-split plot. In 2015, a split-split plot experiment was carried out with water management practice as split plot and N-management practice as split-split plot, where HN variety Dexiang 4103 and LN variety Yixiang 3724 were used as trial materials. Grain yield, root morphology characters, and N absorption and utilization characters, obtained from the above 3 field experiments, were performed multiple collinearity diagnostics through calculating the variance inflation factor. After confirming that there were serious multiple collinearity problems, ridge regression analysis was used as the main data analysis method to investigate the relationships between root morphology characters, N absorption and utilization and grain yield. Based on the recurrence of these relationships from 2013 to 2015, common regularities hidden among these relationships were located. 【Result】The results were as follows: (1) For HN varieties, the determination coefficients of ridge regression equation between grain yield and N accumulation (NA), N dry matter production efficiency (NDMPE), and dry matter weight (DW) at elongation stage, heading stage, and maturity stage, were in the range of 0.0219-0.3961, 0.0452-0.1379, and 0.0914-0.6694, respectively. For LN varieties, the determination coefficients were in the range of 0.0084-0.6190, 0.1224-0.4341, and 0.0818-0.4881, respectively. The relationship between grain yield and N uptake and utilization was less repeatable during the year, and there was no obvious common regularity. Ridge regression analysis was performed with 11 root morphology characters of total root dry weight, adventitious root number, length, surface area, volume, coarse lateral root length, surface area, volume, and fine lateral root length, surface area, volume as independent variables and NA, NDMPE, and DW as dependent variables, respectively. For HN varieties, the determination coefficients were in the range of 0.0527-0.2728, 0.0653-0.3139, and 0.0714-0.3158, respectively. For LN varieties, the determination coefficients were in the range of 0.0607-0.5040, 0.0555-0.4411, and 0.0724-0.5449, respectively. There were significant differences in the relationships between N absorption and utilization and root morphology among years. From 2013 to 2015, the determination coefficients of ridge regression equation between grain yield and root morphology characters of both HN varieties and LN varieties all exceeded 0.8 (<0.001), which showed an common regularity that root morphology at heading stage had a significant effect on grain yield. (2) In the ridge regression equation with root morphology characters as independent variables, and grain yield as dependent variable, the coarse lateral root length of HN varieties obtained the largest standard coefficient, and had the greatest impact on grain yield. However, for the LN varieties, the fine lateral root surface area played the most important role in increasing grain yield. (3) With N rate of 150 kg·hm-2, the optimal N management practice (panicle fertilizer ratio was 40% and the ratio of spikelet preserving fertilizer to panicle fertilizer reached or exceeded 50%) combined with dry-wet alternate irrigation was beneficial to optimize the root morphology. For the SPAD-diagnosis N management practice, N rate reduced from 150 kg·hm-2to 120 kg·hm-2, conventional irrigation mode was more favorable to root morphology optimization than dry-wet alternate irrigation.【Conclusion】Rice root morphology at heading stage had close relationship with grain yield, and appropriate water-N management practice could optimize rice morphology to increase grain yield. SPAD-diagnosis N management practice coupled with conventional irrigation or optimal N management combined with dry-wet alternate irrigation mode could extend the coarse lateral root length of HN varieties as well as the fine lateral root surface area of LN varieties at the heading stage to increase their grain yields, respectively.
N use efficiency; rice; root morphology; grain yield; ridge regression analysis
2016-11-09;接受日期:2017-03-07
国家重点研发计划(2016YFD0300506)、国家粮食丰产科技工程(2013BAD07B13)
李娜,E-mail:lina1409bs@163.com。通信作者马均,E-mail:majunp2002@163.com
