葡萄Actin基因家族的鉴定及进化和表达分析
2017-10-12崔力文张克坤上官凌飞房经贵
崔力文, 郑 婷, 张克坤, 张 川, 上官凌飞, 房经贵
(南京农业大学园艺学院, 江苏 南京 210095)
葡萄Actin基因家族的鉴定及进化和表达分析
崔力文, 郑 婷, 张克坤, 张 川, 上官凌飞, 房经贵①
(南京农业大学园艺学院, 江苏 南京 210095)
采用生物信息学方法从葡萄(VitisviniferaLinn.)全基因组中鉴定Actin基因家族,并对各基因的染色体定位和结构特征,编码蛋白质的理化性质、亚细胞定位、二级结构、三级结构和系统进化,以及不同组织的基因表达进行研究。结果表明:葡萄Actin基因家族16个基因分布在12条染色体上。16个基因的结构特征及其编码蛋白质的理化性质差异较大。16个基因的长度及其内含子总长度的变化范围较大,编码序列(CDS)和外显子总长度的变化范围较小。除登录号GSVIVG01008254001和GSVIVG01014035001的基因外,其他14个基因的GC含量均低于其CDS的GC含量。除登录号GSVIVG01008254001的基因外,其他15个基因编码的蛋白质的理论相对分子质量为12 534.54~82 612.33,理论等电点为pI 4.92~pI 9.13。16个基因编码蛋白质的消光系数为14 105~73 645,脂肪族氨基酸指数为65.54~92.06,其中9个为稳定蛋白,7个为不稳定蛋白。除登录号GSVIVG01014035001的基因外,其他15个基因编码的蛋白质均为亲水性蛋白。登录号GSVIVG01016517001的基因编码的蛋白质定位于细胞质和细胞核,其他15个基因编码的蛋白质定位于细胞质。二级结构和三级结构显示:葡萄Actin基因家族16个基因编码的蛋白质均由α螺旋、无规则卷曲和延伸链构成,且总体以无规则卷曲为主。系统进化分析和不同组织的基因表达分析结果显示:与拟南芥〔Arabidopsisthaliana(Linn.) Heynh.〕相似,葡萄Actin基因家族16个基因编码的蛋白质分为3个亚家族,ClassⅡ亚家族(营养型)包括登录号GSVIVG01003099001和GSVIVG01026580001的基因编码的蛋白质,这2个基因在所有组织中的表达均较高;ClassⅢ亚家族(生殖型)包括登录号GSVIVG01033494001、GSVIVG01024980001和GSVIVG01016550001的基因编码的蛋白质,这3个基因在花粉、雄蕊和花中的表达均较高;ClassⅠ亚家族包括其他11个基因编码的蛋白质,这11个基因在各组织中的表达总体上较低。研究结果显示:葡萄Actin基因家族的表达具有组织特异性。
葡萄;Actin基因家族; 系统进化; 基因表达
Abstract:Actingene family was identified from the whole genome ofVitisviniferaLinn. by bioinformatics method, and chromosome localization and structural characteristics of each gene, physicochemical property, subcellular localization, secondary structure, tertiary structure and phylogeny of proteins encoded, and gene expression in different tissues were researched. The results show that sixteen genes inActingene family ofV.viniferaare distributed on twelve chromosomes. There are great differences in structural characteristics of sixteen genes and physicochemical property of their proteins encoded. Variation ranges of length of sixteen genes and their total length of introns are large, and those of total length of coding sequences (CDSs) and exons are small. Except genes with accession number of GSVIVG01008254001 and GSVIVG01014035001, GC contents in other fourteen genes are lower than those in their CDSs. Except gene with accession number of GSVIVG01008254001, theoretical relative molecular mass of proteins encoded by other fifteen genes is 12 534.54-82 612.33, and their theoretical isoelectric point is pI 4.92-pI 9.13. Extinction coefficient of proteins encoded by sixteen genes is 14 105-73 645, their aliphatic amino acid index is 65.54-92.06, in which, there are nine stable proteins and seven instable proteins. Except gene with accession number of GSVIVG01014035001, proteins encoded by other fifteen genes are hydrophilic proteins. Protein encoded by gene with accession number of GSVIVG01016517001 is located in cytoplasm and nucleus, and those encoded by other fifteen genes are located in cytoplasm. The secondary and tertiary structures show that proteins encoded by sixteen genes inActingene family ofV.viniferaare composed withαhelix, random coil and extended strand, and in general, random coil is the main component. Analysis results of phylogeny and gene expression in different tissues show that being similar toArabidopsisthaliana(Linn.) Heynh., proteins encoded by sixteen genes inActingene family ofV.viniferacan be divided into three subfamilies. Class Ⅱ subfamily (vegetative type) includes proteins encoded by genes with accession number of GSVIVG01003099001 and GSVIVG01026580001, and the expression of these two genes in all tissues is higher. Class Ⅲ subfamily (reproductive type) includes proteins encoded by genes with accession number of GSVIVG01033494001, GSVIVG01024980001 and GSVIVG01016550001, and the expression of these three genes in pollen, stamen and flower is higher. Class Ⅰ subfamily includes proteins encoded by the rest of eleven genes, and the expression of these eleven genes in all tissues is generally low. The results of gene expression analysis show that there is tissue specificity in expression ofActingene family ofV.vinifera.
Keywords:VitisviniferaLinn.;Actingene family; phylogeny; gene expression
自从在脊椎动物骨骼细胞中发现肌动蛋白(actin)以来,陆续在真菌、高等植物[1]以及藻类中发现了肌动蛋白的存在。肌动蛋白广泛存在于植物界,是微丝的主要组分。肌动蛋白参与许多对植物发育至关重要的生理活动,如细胞形状的维持,胞质流动以及细胞的分裂分化、重力感应和信号传导等[2];在尖端生长的花粉管中,膜泡和生殖细胞基于肌动蛋白的运动而移动[3]。有关多核糖体与微丝共定位的研究认为,微丝骨架影响多核糖蛋白在细胞内的分布[4]。植物肌动蛋白丰富的功能和作用由Actin基因家族的多样性决定。多细胞植物具有10个及以上Actin基因,如拟南芥〔Arabidopsisthaliana(Linn.) Heynh.〕[5]有10个Actin基因,碧冬茄(PetuniahybridaVilm.)[6]的基因组中有100多条Actin基因序列,从豌豆(PisumsativumLinn.)[7]卷须中克隆出18个Actin基因。高等植物的Actin基因家族成员具有多样性和表达差异。McDowell等[8]研究了拟南芥Actin基因家族的结构及进化关系,认为拟南芥Actin基因家族至少包含2个古代基因类型,在陆地植物进化早期分化产生生殖和营养2种类型,然后进一步分化产生6种肌动蛋白亚类,其中5种肌动蛋白亚类具有不同的组织特异性表达模式。拟南芥肌动蛋白的8个异型体中有大量的非保守氨基酸替换,所有氨基酸替换都发生在分子表面,除1个Actin基因外,在其余Actin基因的核苷酸序列中的同一位置均含有3个内含子,且每个Actin基因都含有1个180~220 nt的3′-非翻译区(untranslated region,UTR)[9-10]。豌豆肌动蛋白异型体可分为3种类型[11],这3类肌动蛋白异型体基因在根、茎、叶、卷须、花粉和幼嫩果实中均可表达,但在发育时间以及表达强度上存在明显差异,而且具有组织特异性[12-13]。对豌豆Ⅱ类肌动蛋白异型体3个基因的全序列测定结果显示:3个基因的核苷酸序列完全相同,差别仅在于3′-UTR的长度不同。此外,豌豆卷须中3类肌动蛋白异型体基因的核苷酸序列同源性为80%,其氨基酸序列同源性为94%[11]。
葡萄(VitisviniferaLinn.)是世界最古老的果树树种之一,也是世界性的重要水果。2007年,葡萄基因组草图测序完成[14],为后续通过生物信息学方法挖掘、鉴定和分析葡萄基因组奠定了基础。本研究采用生物信息学方法对葡萄全基因组中的Actin基因家族进行鉴定,对Actin基因家族中各基因的结构特征,编码蛋白质的理化性质、亚细胞定位、二级结构、三级结构和系统进化,以及各基因的组织表达模式进行分析,以期为进一步研究葡萄Actin基因的功能提供基础资料。
1 材料和方法
1.1 材料
葡萄全基因组信息由Phytozome v12.0(https:∥phytozome.jgi.doe.gov/pz/portal.html)数据库提供。葡萄芯片表达数据来源于NCBI的GEO数据库,编号GSE36128,包括葡萄54个不同组织样品,覆盖了葡萄不同生长发育阶段的大部分器官[15]。
1.2 方法
1.2.1Actin基因家族数据集的建立和染色体定位分析 首先利用葡萄全基因组序列,构建本地BLAST数据库,以拟南芥Actin基因家族序列为靶序列执行本地BLAST搜索;同时,利用Pfam数据库工具建立葡萄全基因组蛋白质结构域模型,利用HMMER程序筛选含有Actin典型结构域(PF00022)的蛋白质序列[16]。采用E值对筛选结果进行可靠性评价,E值越低越可靠。本研究选用E值小于等于10-5的序列作为候选蛋白质序列,删除重复序列。
从Phytozome v12.0数据库下载的葡萄全基因组信息中提取Actin基因家族各基因的登录号和染色体定位信息。
1.2.2 基因结构特征分析 根据Phytozome v12.0数据库提供的信息,分析基因长度以及编码序列(coding sequence,CDS)、外显子、内含子、3′-非翻译区(UTR)和5′-UTR的数量和总长度,采用DNASTAR软件计算GC含量。
1.2.3 编码蛋白质的理化性质分析 利用ProtParam tool(http:∥web.expasy.org/protparam/)分析Actin基因家族中各基因编码蛋白质的基本理化性质,包括氨基酸残基数量、理论相对分子质量、理论等电点、平均亲水系数、消光系数、不稳定系数及脂肪族氨基酸指数[17]。1.2.4 编码蛋白质的亚细胞定位、二级结构及三级结构分析 参照董晨等[18]的方法利用Plant-mPLoc sever程序进行Actin基因家族各基因编码蛋白质的亚细胞定位;利用SOPMA程序分析蛋白质二级结构;利用Phyre2(http:∥www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index)在线绘制蛋白质三级结构图。
长期以来,我国危险货物国际运输和国内运输在船舶技术条件、货物适运条件等方面是差异化要求,国际运输适用相关国际公约和规则,国内运输则适用国内法规和技术规范。但由于国际和国内运输的货物适运条件不一致,导致危险货物国内运输转国际运输时需按照国际公约规定重新包装、积载、隔离,给货主与船东造成很大不便,也增加安全监管与执法难度。同时,《危险化学品安全管理条例》对危险化学品的适运要求统一适用于国际和国内运输,未有所区分。在实践中,长期以来国内运输包装危险货物已在参照国际公约管理。
1.2.5 系统发育树的构建 通过Clustal Omega程序对拟南芥和葡萄Actin基因家族中各基因编码的蛋白质进行多序列联配比对分析并构建系统发育树。1.2.6Actin基因家族的组织表达分析 在葡萄芯片表达数据库中选取Actin基因家族各基因的表达值,将原值以2为底取对数后采用MeV软件绘制热图。
2 结果和分析
2.1 葡萄Actin基因家族的全基因组鉴定
葡萄Actin基因家族的信息见表1。由表1可见:葡萄Actin基因家族共包括16个基因,且各基因在染色体上的分布比较分散,分别分布在01、04、06、08、11、12、13、14、16、17、18和19号共12条染色体上,其中,13号染色体分布的基因数量最多(3个),11和17号染色体均分布2个基因,其他染色体均分布1个基因。
表1葡萄Actin基因家族的信息
Table1InformationofActingenefamilyofVitisviniferaLinn.

编号No.登录号Accessionnumber染色体定位Chromosomelocalization1GSVIVG01000587001ch01:6782564-68104572GSVIVG01003099001ch12:8678866-86811353GSVIVG01008254001ch17:4031362-40614774GSVIVG01008255001ch17:4013614-40309335GSVIVG01014035001ch16:9532118-95324446GSVIVG01014840001ch19:10434342-104440227GSVIVG01015049001ch11:713295-7148608GSVIVG01015556001ch11:4862914-48670369GSVIVG01016517001ch13:3197303-320762310GSVIVG01016550001ch13:2880916-288302211GSVIVG01024980001ch06:5548568-555219112GSVIVG01026580001ch04:21427667-2143096813GSVIVG01032120001ch13:22480789-2249677614GSVIVG01033049001ch14:25325845-2533437115GSVIVG01033494001ch08:20249818-2025260016GSVIVG01036362001ch18:4225053-4229977
2.2 葡萄Actin基因家族的结构特征分析
葡萄Actin基因家族的结构特征见表2。由表2可见:葡萄Actin基因家族16个基因含1~16个编码序列(CDS)和外显子以及0~15个内含子,登录号GSVIVG01008254001的基因中外显子和内含子数量最多,登录号GSVIVG01014035001的基因中外显子和内含子数量最少。葡萄Actin基因家族16个基因长度的变化范围较大,为327~30 116 bp。除登录号GSVIVG01014035001的基因不含内含子外,其他15个基因中内含子总长度的变化范围较大,为314~28 412 bp。葡萄Actin基因家族16个基因中CDS和外显子总长度的变化范围均较小,分别为327~2 178和326~2 544 bp。
由表2还可见:葡萄Actin基因家族16个基因中GC含量的变化范围为30.48%~43.68%,CDS中GC含量的变化范围为34.09%~50.23%。除登录号GSVIVG01008254001和GSVIVG01014035001的基因外,其他14个基因的GC含量均低于其CDS的GC含量。
由表2还可见:葡萄Actin基因家族16个基因中登录号GSVIVG01008255001、GSVIVG01014035001和GSVIVG01015556001的基因不含3′-非翻译区(UTR),其他13个基因均含1个3′-UTR,且这13个基因中3′-UTR的总长度为120~1 171 bp。登录号分别为GSVIVG01008254001、GSVIVG01014035001、GSVIVG01015049001、 GSVIVG01015556001、GSVIVG01016550001和GSVIVG01036362001的基因不含5′-UTR,登录号GSVIVG01024980001和GSVIVG01033494001的基因含2个5′-UTR,其他8个基因均含1个5′-UTR,且这10个基因中5′-UTR总长度的变化范围较小,为10~373 bp。
表2葡萄Actin基因家族的结构特征1)
Table2StructuralcharacteristicsofActingenefamilyofVitisviniferaLinn.1)

登录号Accessionnumber基因长度/bpGenelengthGC含量/%GCcontentCDS外显子 Exon数量Number总长度/bpTotallengthGC含量/%GCcontent数量Number总长度/bpTotallengthGSVIVG010005870012789438.2311114945.87112110GSVIVG01003099001227041.98586748.1051315GSVIVG010082540013011634.3916132934.09161655GSVIVG010082550011732037.88853141.818644GSVIVG0101403500132741.28132741.281326GSVIVG01014840001968130.4812142245.08121772GSVIVG01015049001156643.684113146.774952GSVIVG01015556001412339.197130250.2371302GSVIVG010165170011032135.9412217844.21122544GSVIVG01016550001210742.764113447.1841440GSVIVG01024980001362438.274113147.6651611GSVIVG01026580001330239.926122746.2162118GSVIVG010321200011598837.407108644.1171464GSVIVG01033049001852734.5815117043.50151680GSVIVG01033494001278340.32587048.5161327GSVIVG01036362001492543.29374145.8831913登录号Accessionnumber内含子 Intron3'-UTR5'-UTR数量Number总长度/bpTotallength数量Number总长度/bpTotallength数量Number总长度/bpTotallengthGSVIVG01000587001102580215861373GSVIVG0100309900149551436110GSVIVG0100825400115284121325——GSVIVG01008255001716746——1112GSVIVG010140350010—————GSVIVG010148400011179091273175GSVIVG0101504900133141120——GSVIVG0101555600162821————GSVIVG010165170011177771320144GSVIVG0101655000136671305——GSVIVG010249800013109313562121GSVIVG010265800015118516751213GSVIVG0103212000161434012551121GSVIVG0103304900114683713381180GSVIVG0103349400148311376278GSVIVG010363620012301311171——
1)CDS: 编码序列Coding sequence; UTR: 非翻译区Untranslated region. —: 无数据No datum.
2.3葡萄Actin基因家族16个基因编码蛋白质的理化性质分析
葡萄Actin基因家族16个基因编码蛋白质的理化性质见表3。结果显示:葡萄Actin基因家族16个基因编码蛋白质的氨基酸残基数量为108~725,登录号GSVIVG01016517001的基因编码蛋白质的氨基酸残基数量最多,登录号GSVIVG01014035001的基因编码蛋白质的氨基酸残基数量最少。除登录号GSVIVG01008254001的基因外,葡萄Actin基因家族中其他15个基因编码蛋白质的理论相对分子质量为12 534.54~82 612.33,理论等电点为pI 4.92~pI 9.13,其中,12个基因编码蛋白质的理论等电点小于pI 6.5,显酸性,3个基因编码蛋白质的理论等电点大于pI 8.0,显碱性。葡萄Actin基因家族16个基因编码蛋白质的消光系数为14 105~73 645,脂肪族氨基酸指数为65.54~92.06。葡萄Actin基因家族16个基因中,9个基因编码蛋白质的不稳定指数小于40,为稳定蛋白;其他7个基因编码蛋白质的不稳定指数大于40,为不稳定蛋白。除登录号GSVIVG01014035001的基因编码的蛋白质为疏水性蛋白外,葡萄Actin基因家族中其他15个基因编码的蛋白质为亲水性蛋白。
表3葡萄Actin基因家族16个基因编码蛋白质的理化性质1)
Table3PhysicochemicalpropertyofproteinsencodedbysixteengenesinActingenefamilyofVitisviniferaLinn.1)
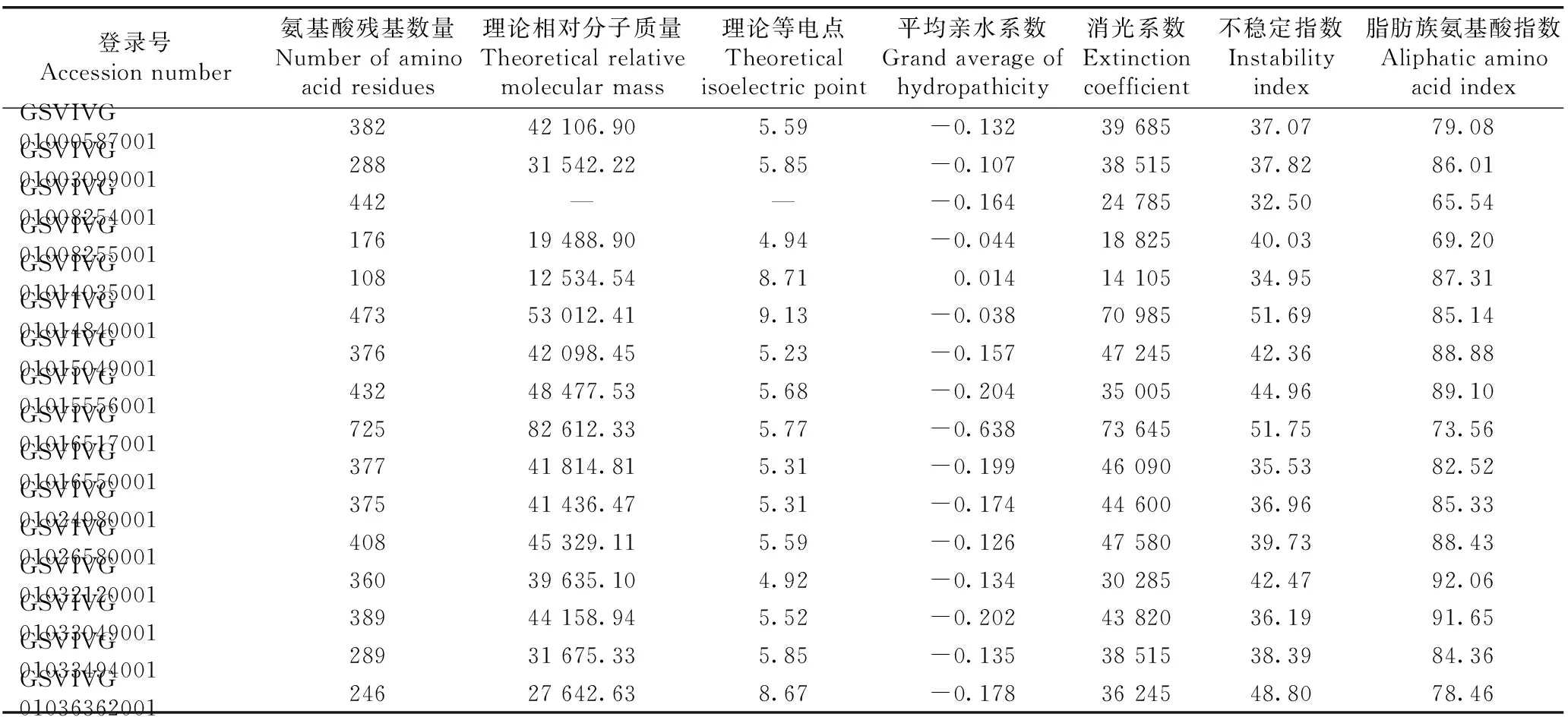
登录号Accessionnumber氨基酸残基数量Numberofaminoacidresidues理论相对分子质量Theoreticalrelativemolecularmass理论等电点Theoreticalisoelectricpoint平均亲水系数Grandaverageofhydropathicity消光系数Extinctioncoefficient不稳定指数Instabilityindex脂肪族氨基酸指数AliphaticaminoacidindexGSVIVG0100058700138242106.905.59-0.1323968537.0779.08GSVIVG0100309900128831542.225.85-0.1073851537.8286.01GSVIVG01008254001442——-0.1642478532.5065.54GSVIVG0100825500117619488.904.94-0.0441882540.0369.20GSVIVG0101403500110812534.548.710.0141410534.9587.31GSVIVG0101484000147353012.419.13-0.0387098551.6985.14GSVIVG0101504900137642098.455.23-0.1574724542.3688.88GSVIVG0101555600143248477.535.68-0.2043500544.9689.10GSVIVG0101651700172582612.335.77-0.6387364551.7573.56GSVIVG0101655000137741814.815.31-0.1994609035.5382.52GSVIVG0102498000137541436.475.31-0.1744460036.9685.33GSVIVG0102658000140845329.115.59-0.1264758039.7388.43GSVIVG0103212000136039635.104.92-0.1343028542.4792.06GSVIVG0103304900138944158.945.52-0.2024382036.1991.65GSVIVG0103349400128931675.335.85-0.1353851538.3984.36GSVIVG0103636200124627642.638.67-0.1783624548.8078.46
1)—: 不明确Undefined.
2.4葡萄Actin基因家族16个基因编码蛋白质的亚细胞定位、二级结构及三级结构分析
葡萄Actin基因家族16个基因编码蛋白质的亚细胞定位和二级结构见表4,16个基因编码蛋白质的三级结构见图1。
2.4.1 亚细胞定位 由表4可见:葡萄Actin基因家族中仅登录号GSVIVG01016517001的基因编码的蛋白质定位于细胞质和细胞核,其他15个基因编码的蛋白质均定位于细胞质。
2.4.2 二级结构 由表4还可见:葡萄Actin基因家族16个基因编码蛋白质的二级结构均由α螺旋、无规则卷曲和延伸链组成,且总体以无规则卷曲为主。登录号GSVIVG01000587001、GSVIVG01003099001、GSVIVG01008255001、 GSVIVG01014035001、GSVIVG01033494001和GSVIVG01036362001的基因编码蛋白质的二级结构中,各构成元件所占比例由高到低依次为无规则卷曲、延伸链、α螺旋;登录号GSVIVG01008254001的基因编码蛋白质的二级结构中,各构成元件所占比例由高到低依次为延伸链、无规则卷曲、α螺旋;其余9个基因编码蛋白质的二级结构中,各构成元件所占比例由高到低依次为无规则卷曲、α螺旋、延伸链。
表4葡萄Actin基因家族16个基因编码蛋白质的亚细胞定位及二级结构
Table4SubcellularlocalizationandsecondarystructureofproteinsencodedbysixteengenesinActingenefamilyofVitisviniferaLinn.

登录号Accessionnumber亚细胞定位1)Subcellularlocalization1)比例/%2) Percentage2)HRCES登录号Accessionnumber亚细胞定位1)Subcellularlocalization1)比例/%2) Percentage2)HRCESGSVIVG01000587001C20.4257.8521.73GSVIVG01016517001C,N36.4645.5817.96GSVIVG01003099001C22.5750.3527.08GSVIVG01016550001C 29.7148.2822.02GSVIVG01008254001C23.9835.9740.05GSVIVG01024980001C 34.3145.4820.21GSVIVG01008255001C5.1165.9128.98GSVIVG01026580001C 27.2146.8125.98GSVIVG01014035001C12.0462.0425.93GSVIVG01032120001C 37.9546.8115.24GSVIVG01014840001C33.1942.7124.10GSVIVG01033049001C 33.9347.8118.25GSVIVG01015049001C37.2342.8219.95GSVIVG01033494001C 22.4950.1727.34GSVIVG01015556001C26.6452.1922.17GSVIVG01036362001C 13.4157.7228.86
1)C: 细胞质Cytoplasm; N: 细胞核Nucleus.
2)H:α螺旋αhelix; RC: 无规则卷曲Random coil; ES: 延伸链Extended strand.
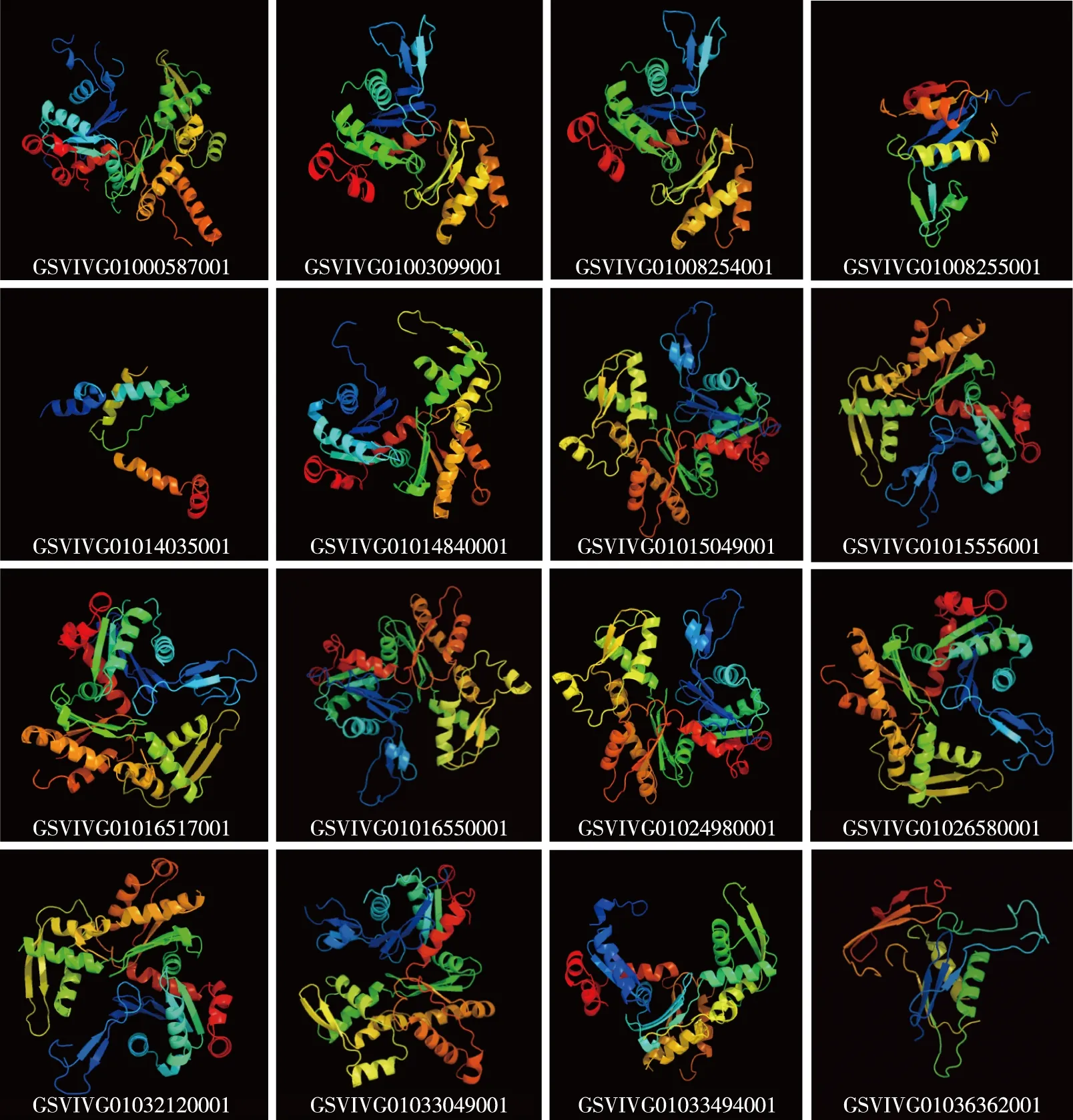
图中编号为基因登录号Numbers in figures are accession numbers of genes.图1 葡萄Actin基因家族16个基因编码蛋白质的三级结构Fig. 1 Tertiary structure of proteins encoded by sixteen genes in Actin gene family of Vitis vinifera Linn.
2.4.3 三级结构 由图1可见:葡萄Actin基因家族中,由α螺旋数量来看,登录号GSVIVG01016517001的基因编码的蛋白质中α螺旋数量最多,为26个;登录号GSVIVG01008254001和GSVIVG01024980001的基因编码的蛋白质均包含14个α螺旋;登录号分别为GSVIVG01000587001、 GSVIVG01003099001和GSVIVG01003099001的基因编码的蛋白质均包含8个α螺旋;登录号GSVIVG01008255001的基因编码的蛋白质中α螺旋数量最少,仅2个。
由延伸链数量来看,登录号GSVIVG01016517001的基因编码的蛋白质中延伸链数量最多,为31个;登录号GSVIVG01000587001、GSVIVG01008254001、GSVIVG01033049001和GSVIVG0100309900的基因编码的蛋白质均包含17个延伸链;登录号分别为GSVIVG01036362001、 GSVIVG01024980001和GSVIVG01033494001的基因编码的蛋白质均包含16个延伸链;登录号GSVIVG01014035001的基因编码的蛋白质中延伸链最少,仅6个。
此外,虽然登录号GSVIVG01000587001和GSVIVG01003099001的基因编码的蛋白质含有相同的α螺旋和延伸链数量,但二者的三级结构存在一定差异,推测这些差异导致二者在功能上有所区别。
2.5葡萄Actin基因家族16个基因编码蛋白质的系统进化分析
基于葡萄Actin基因家族16个基因编码蛋白质的全长序列构建的系统发育树见图2。参照拟南芥Actin基因家族的分类方法[18],葡萄Actin基因家族16个基因编码蛋白质分为ClassⅠ、ClassⅡ和ClassⅢ 3个亚家族,其中,ClassⅡ亚家族属于营养型,包括登录号GSVIVG01003099001和GSVIVG01026580001的基因编码的蛋白质;ClassⅢ亚家族属于生殖型,包括登录号GSVIVG01033494001、GSVIVG01024980001和GSVIVG01016550001的基因编码的蛋白质;ClassⅠ亚家族包括其他11个基因编码的蛋白质。

*: 来自拟南芥From Arabidopsis thaliana (Linn.) Heynh.图2 葡萄与拟南芥Actin基因家族中各基因编码蛋白质的系统发育树Fig. 2 Phylogenetic tree of proteins encoded by each gene in Actin gene family of Vitis vinifera Linn. and Arabidopsis thaliana (Linn.) Heynh.
2.6 葡萄Actin基因家族的组织表达分析
通过葡萄基因芯片平台,对葡萄Actin基因家族(登录号GSVIVG01036362001的基因在GSE36128数据集未出现)进行组织表达分析(图3)。结果显示:ClassⅡ亚家族中登录号GSVIVG01003099001和GSVIVG01026580001的基因在所有组织中的表达均较高;ClassⅢ亚家族中登录号GSVIVG01033494001、GSVIVG01024980001和GSVIVG01016550001的基因在花粉、雄蕊和花中的表达均较高,其中,登录号GSVIVG01033494001的基因在果皮中的表达也较高;ClassⅠ亚家族中除登录号GSVIVG01016517001、GSVIVG01000587001和GSVIVG01032120001的基因在各组织中均有一定表达之外,ClassⅠ亚家族中其他基因在各组织中的表达均较低,尤其是登录号GSVIVG01014035001的基因在所有组织中几乎不表达。

1-15: 基因登录号Accession number of genes. 1: GSVIVG01014840001; 2: GSVIVG01016517001; 3: GSVIVG01015556001;4: GSVIVG01008255001; 5: GSVIVG01014035001; 6: GSVIVG01000587001; 7: GSVIVG01008254001; 8: GSVIVG01032120001;9: GSVIVG01033049001; 10: GSVIVG01015049001; 11: GSVIVG01003099001; 12: GSVIVG01026580001; 13: GSVIVG01033494001; 14: GSVIVG01024980001; 15: GSVIVG01016550001.图3 葡萄Actin基因家族的组织表达模式Fig. 3 Tissue expression pattern of Actin gene family of Vitis vinifera Linn.
3 讨论和结论
肌动蛋白是存在于真核生物细胞质中最丰富的蛋白质,其氨基酸序列也是保守性最高的蛋白质氨基酸序列之一,在整个进化过程中几乎没有改变。肌动蛋白的氨基酸序列在不同种类藻类和人类中的差异不超过20%[19]。拟南芥中10个Actin基因显示出不同的表达模式,并涉及一系列不同细胞功能[20]。基于拟南芥中10个Actin基因表达类型,其中2个Actin基因为假基因,其他8个Actin基因可以分为营养型和生殖型2类,表现出不同的表达模式,并且同一类中各基因间紧密相关[21]。目前,已有关于拟南芥、大豆和芜青(BrassicarapaLinn.)Actin基因家族的全基因组分析[19],并根据其功能分为不同的类型,为进一步研究葡萄Actin基因家族提供了重要的参考价值。
本研究通过对葡萄Actin基因家族进行全基因组生物信息学分析,共鉴定出16个基因,这16个基因编码蛋白质的亚家族分类与拟南芥和芜青[19]相似,说明葡萄Actin基因家族是植物进化中非常保守的家族。虽然Actin基因家族在葡萄全基因组中只有16个基因,但这16个基因的结构特征及其编码蛋白质的理化性质差异较大。其中,基因长度和内含子总长度的差异较大,分别相差29 789和28 098 bp;内含子数量变化范围也较大,在0~15之间。内含子不同程度的插入和缺失构成了基因结构的多样性,在一定程度上,内含子的进化历程反映了基因结构的进化[22]。除登录号GSVIVG01014035001的基因外,葡萄Actin基因家族其他15个基因编码的蛋白质为亲水性蛋白,其理化性质存在较大差异,体现在氨基酸残基数量相差549,理论相对分子质量相差63 123.43,理论等电点相差pI 4.21。
本研究中,葡萄Actin基因家族中Class Ⅱ和Class Ⅲ亚家族分别属于营养型和生殖型,这2个亚家族中的基因在葡萄生长发育过程中具有重要作用[23]。ClassⅡ和ClassⅢ亚家族Actin基因中,基因和编码序列(CDS)的GC含量以及CDS、外显子、内含子、3′-非翻译区(UTR)和5′-UTR的数量接近,其中,Class Ⅱ和Class Ⅲ亚家族基因的GC含量分别相差2.06%和4.49%,CDS的GC含量相差更少,分别仅1.89%和1.33%。Class Ⅱ和Class Ⅲ亚家族基因编码蛋白质的理论等电点均小于pI 6.5,显酸性;并且,不稳定指数均小于40,为稳定蛋白,在一定程度上反映出其功能的稳定性。
与其他植物相比,拟南芥的芯片数据更加全面,并已得到广泛验证。本研究通过聚类分析比较了葡萄和拟南芥Actin基因家族中各基因编码蛋白质间的进化关系,认为这2种植物间的进化关系具有保守性,同一亚家族中的Actin基因可能具有相似的基因功能。葡萄Actin基因家族中,Class Ⅱ亚家族基因在所有组织中的表达均较高,Class Ⅲ亚家族基因在花粉、雄蕊和花中的表达均较高,ClassⅠ亚家族中多数基因在各组织中的表达均较低,这与拟南芥Actin基因家族的表达情况相似,因此,可根据拟南芥中已有功能注释的Actin基因,对葡萄Actin基因进行功能预测。拟南芥中ACT7基因(登录号AT5G09810.1)不仅在愈伤组织形成过程中发挥着重要作用[5],还可能是对外界刺激反应最强烈的Actin基因[24]。葡萄Actin基因家族中登录号GSVIVG01026580001和GSVIVG01003099001的基因与拟南芥中ACT7基因共同聚在Class Ⅱ亚家族中,推测这2个基因可能具有相似的功能。拟南芥中ACT12(登录号AT3G46520.1)和ACT4(登录号AT5G59370.1)基因主要在花粉管伸长过程中表达[23],推测与二者共同聚在Class Ⅲ亚家族中登录号GSVIVG01016550001的基因也可能在花粉管伸长过程中起重要作用。说明葡萄Actin基因家族的表达在调控葡萄生长发育过程中发挥着重要作用。3个亚家族中基因的表达具有组织特异性,暗示Actin基因家族中不同基因具有特异性功能,但每个亚家族中的基因对葡萄生长发育进程的具体调控机制尚有待进一步研究。
本研究采用生物信息学方法,对葡萄Actin基因家族进行全基因组分析,为验证其基因功能提供了重要参考信息,但基因的具体功能还需进一步研究。
[1] 阎龙飞, 石德权. 高等植物中的收缩蛋白[J]. 生物化学与生物物理学报, 1963, 3(4): 490-496.
[2] 刘 曦, 张少斌, 汪 澈. 植物肌动蛋白功能的研究进展[J]. 生物技术通报, 2010(3): 13-16.
[3] MASCARENHAS J P. Molecular mechanisms of pollen tube growth and differentiation[J]. The Plant Cell, 1993, 5: 1303-1314.
[4] ITO Y, ABE S, DAVIES E. Co-localization of cytoskeleton proteins and polysomes with a membrane fraction from peas[J]. Journal of Experimental Botany, 1994, 45: 253-259.
[5] KANDASAMY M K, MCKINNEY E C, MEAGHER R B. Functional nonequivalency of actin isovariants inArabidopsis[J]. Molecular Biology of the Cell, 2002, 13: 251-261.
[6] BAIRD W V, MEAGHER R B. A complex gene superfamily encodes actin in petunia[J]. The EMBO Journal, 1987, 6: 3223-3231.
[7] 曹晓风, 王荣臣, 阎隆飞, 等. 豌豆卷须cDNA文库构建及肌动蛋白基因序列分析[J]. 科学通报, 1993, 38(19): 1804-1808.
[8] MCDOWELL J M, HUANG S, MCKINNEY E C, et al. Structure and evolution of the actin gene family inArabidopsisthaliana[J]. Genetics, 1996, 142: 587-602.
[9] AN Y Q, HUANG S, MCDOWELL J M, et al. Conserved expression of the ArabidopsisACT1 andACT3 actin subclass in organ primordia and mature pollen[J]. The Plant Cell, 1996, 8: 15-30.
[10] HUANG S, AN Y Q, MCDOWELL J M, et al. TheArabidopsisthalianaACT4/ACT12 actin gene subclass is strongly expressed throughout pollen development[J]. The Plant Journal, 1996, 10: 189-202.
[11] 胡松年, 阎隆飞. 豌豆卷须肌动蛋白Ⅱ类异型体cDNA克隆的序列分析[J]. 中国生物化学与分子生物学报, 1999, 15(6): 857-860.
[12] 凌 毅, 赵武玲. 豌豆肌动蛋白异型体基因的特异性表达[J]. 植物学通报, 2001, 18(1): 76-80.
[13] JIANG Y Q, ZHAO W L. Expression and phylogenetic analysis of pea actin isoforms[J]. 植物学报(英文版), 2002, 44(12): 1456-1461.
[14] JAILLON O, AURY J M, NOEL B, et al. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla[J]. Nature, 2007, 449: 463-468.
[15] FASOLI M, SANTO S D, ZENONI S, et al. The grapevine expression atlas reveals a deep transcriptome shift driving the entire plant into a maturation program[J]. The Plant Cell, 2012, 24: 3489-3505.
[16] 郭景康, 陈青云, 戢 茜, 等. 拟南芥、水稻和杨树ACTIN家族全基因组分析[J]. 上海大学学报(自然科学版), 2009, 15(4): 426-431.
[17] ARTIMO P, JONNALAGEDDA M, ARNOLD K, et al. ExPASy: SIB bioinformatics resource portal[J]. Nucleic Acids Research, 2012, 40: W597-W603.
[18] 董 晨, 贾利强, 决登伟, 等. 香蕉α-淀粉酶基因家族的系统进化分析[J]. 中国南方果树, 2016, 45(5): 12-17, 22.
[19] GUNNING P W, GHOSHDASTIDER U, WHITAKER S, et al. The evolution of compositionally and functionally distinct actin filaments[J]. Journal of Cell Science, 2015, 128: 2009-2019.
[21] MEAGHER R B, MCKINNEY E C, KANDASAMY M K. Isovariant dynamics expand and buffer the responses of complex systems: the diverse plantactingene family[J]. The Plant Cell, 1999, 11: 995-1005.
[22] 魏 潇, 刘威生, 刘 宁, 等. 果实软化相关PG基因的进化分析和基因组定位[J]. 园艺学报, 2011, 38(9): 1791-1799.
[23] 张少斌, 刘国琴. 植物肌动蛋白异型体研究进展[J]. 植物学通报, 2006, 23(3): 242-248.
[24] MCDOWELL J M, AN Y Q, HUANG S, et al. The ArabidopsisACT7 actin gene is expressed in rapidly developing tissues and responds to several external stimuli[J]. Plant Physiology, 1996, 111: 699-711.
(责任编辑: 张明霞)
Identification,evolutionandexpressionanalysesofActingenefamilyofVitisvinifera
CUI Liwen, ZHENG Ting, ZHANG Kekun, ZHANG Chuan, SHANGGUAN Lingfei, FANG Jinggui①
(College of Horticulture, Nanjing Agricultural University, Nanjing 210095, China),J.PlantResour. &Environ., 2017,26(3): 1-10
Q943.2; S663.1
A
1674-7895(2017)03-0001-10
10.3969/j.issn.1674-7895.2017.03.01
2017-03-22
国家自然科学基金资助项目(31672131)
崔力文(1991—),女,山东莒县人,硕士研究生,主要从事果树遗传育种与分子生物学方面的研究。
①通信作者E-mail: fanggg@njau.edu.cn
