庙岛植物多样性差异与原因探讨
2017-09-22何俊徐陆煜李加庆张坚强邓传远
何俊++徐陆煜++李加庆++张坚强++邓传远
摘要:选择6个20m × 20m的具有代表性的样地群落,调查与分析了样方群落内α多样性,群落间β多样性,以及比较了附近山体群落多样性的差异。结果表明:庙岛主要植被类型为刺槐林,黑松林以及6个典型群落。样地群落内共有38种植物,草本种类较多,反映出冬季需经历较长时间。群落间的乔木层与灌木层的Jaccard指数,Bray-Curtis指数均较低,异质性较高,对于外来物种需谨慎使用。灌木层的Margalef丰富度,Shannon-Wiener指数,Simpson指数,Pielou 均匀度相对于乔木层较大,这与乔木层物种遮光能力有关。廟岛植物多样性较陆地山体植物群落多样性低,可能与演替阶段或生境差异有关。
关键词:庙岛;植被;植物群落;多样性;相似性
中图分类号:S602
文献标识码:A文章编号:16749944(2017)16010705
1引言
海岛由于与陆地隔离,植物种群相对较为单一[1]。植物群落具有一定的外貌和结构,能反映植物生境的特点,而多样性的研究能更多反映植物物种的丰富度和群落的稳定性[2]。现今对陆地山体植物多样性研究较为成熟并有相应调查技术规范[3],而关于海岛植物多样性的调查与研究则不多见[4,5]。由于海陆生境差异与自然灾害等原因造成海岛植物多样性与陆地山体具有一定的差异性,这使得海岛植物群落调查具有研究意义。庙岛群岛位于山东烟台市长山县,由于人类长期历史活动,天然植被受到影响。关于庙岛群岛的研究主要集中在植物区系[6~9],α多样性指数[9,10],以及野生观赏植物资源的应用探讨[11],而对于庙岛主岛的植物群落多样性研究不够深入。本文从α多样性,β多样性指数以及与附近山体群落多样性的比较,3个方面进行探讨山东庙岛植物群落内多样性,植物群落间的相似程度,同时反映与陆地山体植物多样性的差异性,为庙岛植物多样性保护与生态修复提供基础数据和借鉴参考的意义。
2研究样地概况与研究方法
2.1研究样地概况
庙岛位于山东烟台市长岛县,37°56′18″N,120°40′48″E,海岛面积约为5.43 km2,海岸线长度为6.7 km,岛上海拔最高点为98.3 m[6]。庙岛属暖温带季风性气候,地带性植被为落叶阔叶林,以栎林为主要类型,主要树种有麻栎,槲栎等。庙岛年平均气温为12.012 ℃,气温年差26.1 ℃。年均降水量为549.5 mm,集中在6~9月,岛上淡水资源较为缺乏。岛上土壤主要为棕壤土类和褐土土类。岛上植物种类约有378种[6],由于长期的人为活动,岛上植被受较大影响,主要以黑松林与刺槐林为主。
2.2样方设置与调查
2015年9月对山东庙岛调查,参考朱春玲等[4]的海岛植物调查的方法,选择具有代表性的植物群落作为样地,共计6个20 m×20 m的样方,记录样方内的乔木(胸径≥5 cm 或树高≥3 m)树种名称,胸径,冠幅等。在每个20 m×20 m的样方内划定4个5m×5m 小样方,记录样方内的灌木名称,棵数,冠幅和株高。在5 m×5 m 小样方内划定5个1 m ×1 m的小样方记录样方内的草本植物名称,棵/丛数,株高,盖度等信息。同时,对样方群落的经纬度、海拔、群落郁闭度等信息进行记录。
2.3群落多样性计算
本文选用Berger-Parker 优势度指数,Margalef 丰富度指数,Simpson 多样性指数,Shannon 多样性指数,Pielou 均匀度指数用来衡量群落内的α多样性指数,选用Jaccard 指数,Bray-Curtis 相似性指数来衡量群落间的β多样性指数[12]。
Berger-Parker 优势度指数:
d=1/nmaxN(1)
Margalef丰富度指数:
dMa=S-1lnN(2)
Simpson多样性指数D:
D=1-∑Si=1P2i;P2i=ni(ni-1)N(N-1)(3)
Shannon-Wiener多样性指数H′e:
H′e=-∑Si=1PilnPi;Pi=niN(4)
Pielou均匀度指数Je:
Je=H′eH′max;H′max=lnS(5)
Jaccard指数Cj:
Cj=ja+b-j(6)
Bray-Curtis相似性指数:
CN=2NjNa+Nb(7)
式中:nmax为个体数量最多物种的个体数量,S为样方内物种数,N为所有物种个体的总数,Ni为第i种的个体数,i=1,2,3,…,S。J为共同种 Nj 指样地a和b 共同种中个体数较少者。Na,Nb 分别指样地A和B的物种数。
3研究结果
3.1植被概况
根据吴征镒《中国植被》[13]将庙岛植被类型划分为落叶阔叶林、常绿针叶林,2个植被类型;有合欢-一叶荻-茜草群落、刺槐-荆条-天门冬群落、构树-一叶荻-迎春群落、黑松-扁担木-鸭葱群落、黑松-一叶荻-茜草群落、黑松-柘-天门冬群落,6个群落,简述如下。
3.1.1落叶阔叶林
(1)刺槐-荆条-天门冬群落(Robinia pseudoacacia - Vitex negundovar.heterophylla- Asparagus cochinchinensis Association),群落外貌整齐,刺槐林约为6 m,胸径为8.1~21.2 cm,平均冠幅为3.8 m,林下植物种类较多。
(2)构树-一叶荻-迎春群落(Broussonetia papyrifera-Flueggea suffruticosa-Jasminum nudiflorum Association),群落生长构树35株,桃树3株。群落高度约为3 m左右,胸径为2.5~4 cm,平均冠幅为3.1 m,灌木层以一叶荻占优势,草本层以迎春占绝对优势。endprint
(3)合欢-一叶荻-茜草群落(Albizia julibrissin - Flueggea suffruticosa - Rubia cordifolia Association),群落外貌整齐,高度为0.8~5 m,胸径为5~10.2 cm,平均冠幅为2.3 m,林下植物草本物种多,盖度约为30%~60%。群落的伴生植物有酸枣,沙参等。
3.1.2常绿针叶林
(1)黑松-扁担木-蒙古鸭葱群落(Pinus thunbergii - Grewia biloba -Scorzonera mongolica Association),群落高度約为4.5~12 m左右,胸径为3.8~27.8 cm,平均冠幅为3.3 m,乔木层黑松重要值较大,刺槐也占有一定优势。灌木层以扁担木占优势,同时灌木层还有麻栎幼苗,及酸枣占大多数。草本层物种丰富,优势并不明显。
(2)黑松-一叶荻-茜草群落(Pinus thunbergii - Flueggea suffruticosa - Rubia cordifolia Association),群落结构整齐,高度约为3~13 m左右,胸径为3.8~20.4 cm,平均冠幅为2.7 m,灌木层仅有一叶荻,草本层以茜草为优势种,林下散生菊花,萱草,萝藦等。
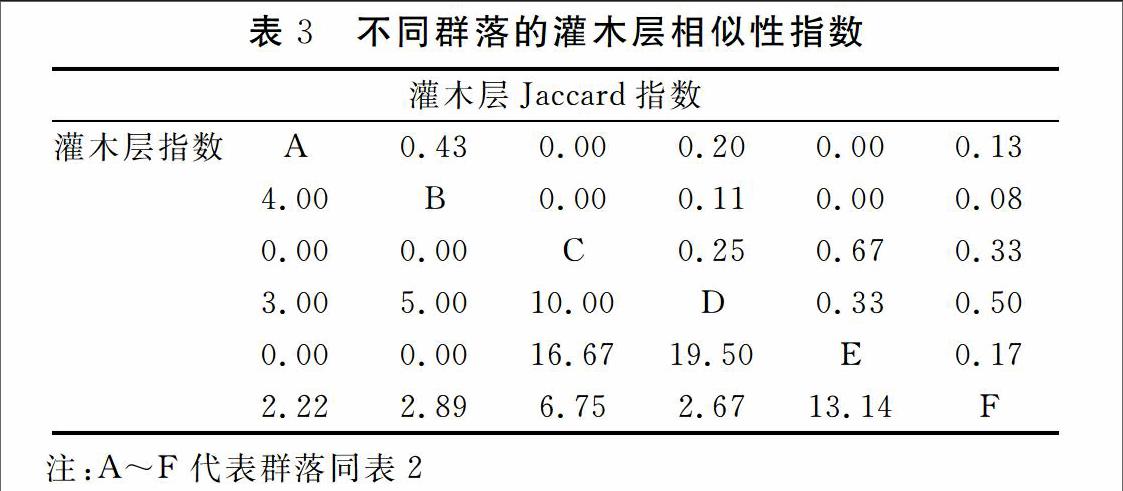
3.3植物群落相似性
Jaccard指数能反映群落之间的物种组成相似程度,而Bray-Curtis指数能体现种的多度在群落物种相似程度。不同群落的乔木层与灌木层相似性指数分别见表2、3。在乔木层中,群落A与E的Jaccard指数最大为1.0,Bray-Curtis指数为9.0。群落A与B的Jaccard指数为0.5,Bray-Curtis指数为3.67。群落B与E之间Jaccard指数0.50,Bray-Curtis指数分别为3.33。群落D与F的Bray-Curtis指数最大为12.25,而Jaccard指数为0.14,群落C与E的Bray-Curtis指数为9.0,而Jaccard指数仅为0.33。群落B与C Bray-Curtis指数 3.67,Jaccard指数为0.2。群落D与其他群落的异质性较强,Jaccard指数和Bray-Curtis指数均为0。
注:A:刺槐-荆条-天门冬群落;B:黑松-扁担木-蒙古鸦葱 群落; C:合欢-一叶荻- 茜草群落; D:构树-一叶荻-迎春 群落; E:黑松-一叶荻-茜草 群落; F:赤松-柘-天门冬群落
在灌木层中,群落C与E的Jaccard指数最大为0.67,Bray-Curtis指数为16.67。群落D与F的Jaccard指数为0.5,Bray-Curtis指数为2.67。群落A与B之间Jaccard指数 0.43,Bray-Curtis指数为4.0。群落D与E的Bray-Curtis指数最大为19.50,Jaccard指数为0.33,群落E与F 的Bray-Curtis指数为13.14,Jaccard指数仅为0.17。群落C,D分别对群落A,B之间的异质性较强,Jaccard指数和Bray-Curtis指数均为0。
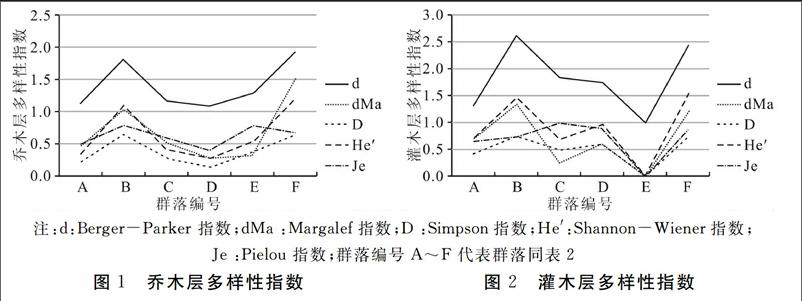
3.4植物群落多样性
庙岛植物群落多样性见图1、2。在6个群落中,灌
4讨论与结论
4.1植物组成
样方群落内共有38种植物,其中乔木层的主要的群落优势种为黑松、刺槐、构、麻栎、槲栎等。灌木优势种主要为荆条、紫穗槐、酸枣、柘、扁担木等。群落内重要值较大的植物均为落叶植物,常绿植物较少仅有黑松,较为符合庙岛暖温带气的植被类型落叶阔叶林的特点。6个海岛植物群落由38种植物组成,乔木9种,灌木12种,草本植物15种,比例为3∶4∶5。草本植物占比例较大,灌木其次。该比例与马成亮[6]提出的地面芽植物所占比例最大,高位芽植物其次,1年生和隐芽植物比例较大的结论较为一致。从群落的植物生活型构成比例即可反映植物生境的特点,暖温带植物生活型相对于亚热带植物生活型,其地面芽植物在群落内的比例增大,高位芽植物比例降低[17]。即庙岛植物冬季需经历长时间的严寒季节,而由于沿海气候的影响,植物在生长季节的生活环境为温热多湿。
4.2海岛植物群落相似性与生态干扰
Jaccard指数 表示物种种类相似度,而Bray-Curtis指数更强调种的个体数量,本文通过研究两个群落的相似性指数进行比较群落间的物种组成及其共同种个体数量的相似程度。在乔木层中,群落A与E的物种组成相同,因此Jaccard指数最大为1.0,Bray-Curtis指数为9.0。群落A与B,群落B与E的物种组成相似度大,个体数量较为相似,因此Jaccard指数为0.50,Bray-Curtis指数较为接近。群落D与F,群落C与E,群落B与C的Bray-Curtis指数最大达12.25,但Jaccard指数较小仅为0.14,表明两群落间物种组成相似程度较低,共同种种类较少,但个体数量较大。群落D与其他群落的相似性指数为0,异质性较强。在灌木层中,群落C与E的Jaccard指数最大为0.67,Bray-Curtis指数为16.67。群落D与E,群落E与F的Jaccard指数分别为19.50和13.14,而Bray-Curtis指数却仅为0.33,0.17。其余样方的群落间相似性很小或为0。整体上看,群落之间的乔木层与灌木层的相似性指数较小,表明山东庙岛植被景观破碎度较大,植物群落间异质性较大。
适生能力强的外来物种,易在异质性较大的群落间对抗干扰程度较低的乡土植物造成威胁[18],外来植物可能对植物群落产生干扰与威胁。庙岛植物群落异质性较大,而群落生长的黑松,刺槐,紫穗槐等原产地并非中国,因人为造林活动被引种栽培于此,并作为建群种和优势种,在岛上形成相应的林分。有研究表明,引进种能构建新的稳定的群落[19],然而引进种易导致本地物种濒临灭绝[20]。因此对引进植物进行绿化栽植或生态修复选用时应慎重考虑是否会对本地种造成威胁,需要进一步探讨。endprint
4.3海岛植物群落多样性
在6个样方群落中,乔木层中以黑松为建群种的群落,黑松优势度突出,且群落内的Margalef指数,Simpson指数,Shannon-Wiener指数,Pielou 指数相对于其他群落的乔木层较大,表明黑松群落在庙岛上适生强度大,且对群落的多样性与均匀度具有一定的提高。该结论与高远等[21]一致,外来种对群落多样性具有维持和提高的作用。灌木层中,植物群落演替后期,由于竞争趋势稳定,优势种突出,而劣种被淘汰,因此Simpson指数,Shannon-Wiener指数降低,Pielou 指数较大。灌木层中一叶荻为优势种的群落较为符合该演替规律。总的来说,灌木层的Margalef丰富度指数,Simpson多样性指数,Shannon-Wiener 多样性指数,Pielou 指数相对于乔木层较大。主要原因在于庙岛地处暖温带,乔木层较多为落叶或针叶植物,对阳光的遮挡能力较小,林下层植物获取阳光的能力增加,局部生境温度与光照升高,林下层植物生存与竞争的能力增加,物种丰富度变大。
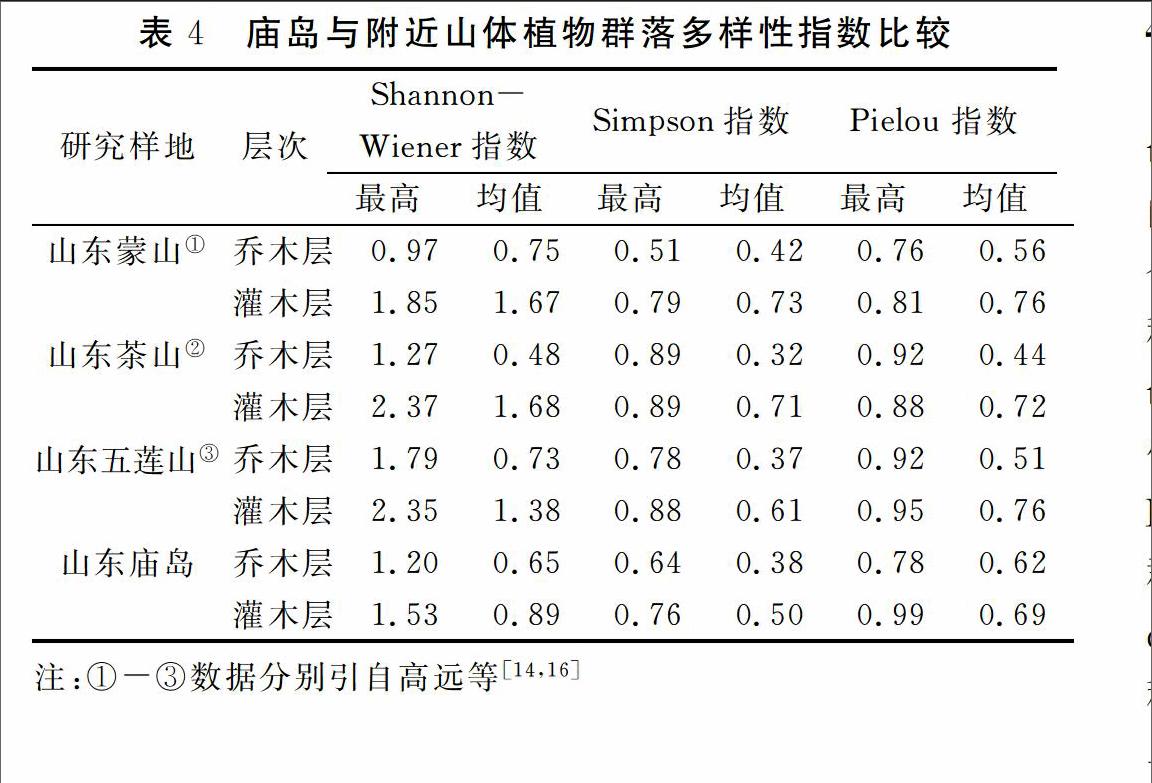
相对于山东蒙山、茶山和五莲山的植物群落Shannon-Wiener多样性指数,Simpson多样性,庙岛的乔木层与灌木层的较小,表明海岛植物群落的多样性相对山体植物群落较小,因此,在对庙岛进行植被生态修复工作时应对其物种多样性加以保护与利用。庙岛最好的群落Pielou 指数相对山体植物群落的高,庙岛的乔木层的Pielou 指数相对较高,灌木层的Pielou 指数略低,表明庙岛乔木层的相对山体植物群落较为均匀。
庙岛植被与陆地山体植被的Shannon-Wiener指数,Simpson指数,Pielou 指数的差距可能由于演替阶段不同或生境差异造成的。①可能庙岛植被演替阶段属于后期阶段,因此其Pielou 指数相对较大,群落结构相对稳定。物种间的竞争下降,Shannon-Wiener多样性指数,Simpson多样性相对较低,这需要进一步证明。②由于庙岛生境较陆地生境恶劣,海拔低,土壤盐分高,淡水少,风速较大等原因造成适生植物种类较少,而陆地植物群落由于山体地形因素形成相对适宜植物生长的小气候,植物物种丰富度大,物种多样性也相对较大。而山东大多为次生林[22],人为活动干扰较大,导致山体的群落多样性较庙岛植物差异性不大。
参考文献:
[1] 邬建国.岛屿生物地理学理论:模型与应用[J].生态学杂志,1989,8(6):34~39.
[2] Ricklefs R E.Community diversity:relative roles of local and regional processes[J].Science,1987(235):167~171.
[3] 方精云,王襄平,沈泽昊,等.植物群落清查的主要内容、方法和技术规范[J].生物多样性,2009,17(6):533~548.
[4] 朱春玲,韓玉洁,谢锦忠,等.上海大金山岛森林群落调查与特征分析[J].林业科技开发,2008,22(6):57~59.
[5] 朱弘,库伟鹏,戎建涛,等.浙江南麂岛陆生维管束植物多样性及区系特征[J].植物分类与资源学报,2015,37(6):713~720.
[6] 马成亮,程贯召,姜岩.庙岛种子植物区系研究[J].河南农业科学,2014,43(12):121~124.
[7] 马成亮,宋桂全.庙岛群岛种子植物区系的研究[J].安徽农业科学,2012,40(18):9577~9579,9581.
[8] 马成亮.庙岛群岛药用植物区系及多样性研究[J].安徽农业科学,2011,39(16):9607~9608.
[9] 马成亮.山东长岛列岛植物区系及群落结构研究[D].南京:南京林业大学,2007.
[10] 张骞.庙岛群岛植物群落结构研究[D].济南:山东师范大学,2008.
[11] 马成亮.庙岛群岛野生观赏植物资源与利用[J].安徽农业科学,2011,39(14):8550~8552,8575.
[12] 张金屯.数量生态学(第二版)[M].北京:科学出版社,2011:1~372.
[13] 吴征镒.中国植被[M].北京:科学出版社,1980:144~156.
[14] 高远,慈海鑫,邱振鲁,等.山东蒙山植物多样性及其海拔梯度格局[J].生态学报,2009,29(12):6377~6384.
[15] 高远,朱秀林,邱振鲁.山东茶山植物群落结构及物种多样性[J].安徽农业科学,2007,35(16):4841~4843.
[16] 高远,姚亮,邱振鲁,等.山东五莲山植物群落结构及物种多样性[J].植物研究,2008,28(3):359~363.
[17] 冷平生.园林生态学[M].北京:中国农业出版社,2003:196.
[18] 覃凤飞,安树青,卓元午,等.景观破碎化对植物种群的影响[J].生态学杂志,2003,22(3):43~48.
[19] Richburg,Julie A,Patterson,William A.Historical Description of the Vegetation of the Boston Harbor Islands:1600-2000[J].Northeastern Naturalist,2005,12(3):13~30.
[20] Losos JB.Ecological and evolutionary determinants of the species-area relation in Caribbean anoline lizards.Philosophical Transactions[J].Biological Sciences,1996(351):847~854.
[21] 高远,朱孔山,郝加琛,等.山东蒙山6种造林树种40余年成林效果评价[J].植物生态学报,2013,37(8):728~738.
[22] 刘冰.中国常见植物野外识别手册山东册[M].北京:高等教育出版社,2009:1~379.endprint
