莱州湾强壮滨箭虫种群数量及个体大小季节分布
2017-09-12时永强
左 涛,王 俊,时永强
1 中国水产科学研究院黄海水产研究所,青岛 266071 2 青岛海洋科学与技术国家实验室海洋生态与环境科学功能实验室,青岛 266200 3 农业部海洋渔业可持续发展重点实验室,青岛 266071 4 山东省渔业资源与生态环境重点实验室,青岛 266071
莱州湾强壮滨箭虫种群数量及个体大小季节分布
左 涛1,2,3,*,王 俊1,2,4,时永强1,2,3
1 中国水产科学研究院黄海水产研究所,青岛 266071 2 青岛海洋科学与技术国家实验室海洋生态与环境科学功能实验室,青岛 266200 3 农业部海洋渔业可持续发展重点实验室,青岛 266071 4 山东省渔业资源与生态环境重点实验室,青岛 266071
基于2011年5月至2012年4月(12 月和翌年1—2月冰期除外)在渤海莱州湾逐月采集的数据资料,分析莱州湾强壮滨箭虫丰度空间分布和体长的季节变化,估算其生产力和摄食率,以掌握其种群动态特征,为评价该种在莱州湾生态系统结构功能中地位提供参数和依据。结果显示:强壮滨箭虫丰度和生物量年变化曲线为双峰型,最高峰出现于5月,次高峰出现于8—9月;4 mm和15 mm体长的个体分别是丰度和生物量的主要贡献者。生产力估值为0.25 mg C m-3d-1,摄食率估值为0.88 mg C m-3d-1。种群月均体长以3—4月最高(>10 mm)、5月和11月最低(<5 mm);月均体长与月表层水温、盐度呈负相关。春季5月、夏季7—8月和秋季10—11月中,小型个体丰度所占的比例较高,应为种群数量增长期。各月的体长频数分布可分辨出平均体长<5mm(Cohort I)、介于5—10 mm(Cohort II)和>10 mm(Cohort III)的3个同生群。Cohort III和Cohort I高丰度区的表层水温和盐度分别介于15—17℃和29—32。Cohort III基本不出现于水温大于24℃的站位,其丰度与水温呈显著负相关。Cohort I基本不出现于水温低于11℃的站位,其丰度与水温呈显著正相关。Cohort II高丰度区的表层水温和盐度为26—27℃和30—32。由上推测强壮滨箭虫发生明显体型更替的水温阈值应不超过15—17℃。
强壮滨箭虫;体长;丰度;生产力;季节分布;莱州湾
强壮滨箭虫(Aidanosagittacrassa(Tokioka,1938))是渤海的毛颚动物中唯一优势种。它在渤海周年出现[1],是渤海季节性浮游动物生物量的主要贡献者[2]。它既是上、中层鱼类和幼鱼的食物组分[3],又是众多浮游动物[4]以及幼鱼[5-6]的捕食者,在渤海生态系统结构和功能中占据着重要的地位。莱州湾是渤海的三大海湾之一,是中国北部传统的渔业生物产卵、孵育和索饵场。近年来,莱州湾内生物群落结构和格局已发生重大改变[7-8],原有的重要经济渔业生物如中国明对虾、三疣梭子蟹、海蜇等需通过增殖放流来实现资源部分修复。这些放流种类的幼苗多以小型浮游动物为食[9],客观上与强壮滨箭虫形成了食物竞争关系。因此,深入了解莱州湾强壮滨箭虫数量分布及其季节变化,掌握其种群结构动态,对于了解湾内生物环境状况和次级生产水平、评价和指导增殖放流有重要意义。
强壮滨箭虫存在着明显的季节生态变型和更替[10],各生态型在形态上、时空(出现季节和垂直分布)偏好差异大,生活周期不尽相同[11]。如日本濑户内海[12]、东京湾[13]和胶州湾外[14]不同季节出现的强壮滨箭虫生活世代周期从3个月至5个月不等。国内有关强壮滨箭虫的研究较多,包括数量分布[1,15-17]、体型变化[18]、年代际变化[19]及其生态指示作用[20-21],以及摄食[22]及其生理[23-24]、温盐耐受性[25-26]等实验生态研究。但渤海强壮滨箭虫的相关研究资料更多来源20世纪50年代[17,20]和90年代[1],近期研究报道[22]很少。本研究以莱州湾作为研究海域,利用近年采集的浮游生物周年调查资料,分析强壮滨箭虫种群丰度时空分布,着重开展了强壮滨箭虫个体大小及数量分布季节变动研究,估算莱州湾强壮滨箭虫的现存量和生产力水平。本研究可为量化莱州湾生态系统功能群结构,优化增殖放流预测提供有效的基础数据和参数。
1 材料与方法
1.1 样品采集与分析
2011年5—11月、2012年3—4月用渔船“鲁昌渔4193号”(额定功率350kW)在莱州湾(37.25°—38.75°N,119°—120.75°E)进行了9个航次逐月定点综合调查,站位布置可参见后文图5和图6。强壮滨箭虫样使用浮游生物网(网孔径为160 μm,网口面积为0.2 m2)从底至表垂直拖网采集获得,甲醛固定保存。该网孔径对于强壮滨箭虫幼体和成体都具有较高的捕获效率[27]。所有采样都是在白天操作完成。强壮滨箭虫样品在实验室Nikon SMZ1500体式显微镜下分样、镜检、计数和体长测量。体长(BL)为从头的前端至尾鳍末端,精确至0.1mm。
1.2 数据处理
个体计数结果转化为丰度(单位滤水量个数),用个m3表示。个体干重生物量W(μg)则基于体长-生物量的经验公式W=0.197BL3.01[28]换算得到。生产力(mg C m-3d-1)和摄食率(mg C m-3d-1)的估算过程参考文献[14,18],即先由干重生物量推得呼吸率[29]、再将呼吸率转换成相应的生产力和摄食率[13]。
体长组值取1mm间隔分组后的体长上限,各体长组的丰度相对频数为其丰度占所有体长组的总丰度的比值。强壮滨箭虫体长-相对频数分布利用混合分布法[30]拟合正态曲线,经方差分析(ANOVA)检验后优化和确定拟合曲线与原观测值的差异不显著(P>0.05),方接受拟合后曲线和各拟合分布组或同生群的分布。甲醛固定保存强壮滨箭虫的体长损耗最大为0.4mm[12]或小于10%[31],小于分组间隔1mm,因此本文不考虑固定保存所引起的强壮滨箭虫个体形态损耗。
平面图和线型图分别采用Mapviewer 7.0和Grapher 8.0(Golden software,Inc.)绘制。
2 结果
2.1 丰度、生物量和生产力、摄食率估算
强壮滨箭虫月均丰度为186 个/m3,生物量为12.6 mg/m3(或174 mg/m2),生产力估值为0.25 mg C m-3d-1(或3.57 mg C m-2d-1),摄食率月均值为0.88 mg C m-3d-1(或10.25 mg C m-2d-1)。它们的季节变化曲线都为双峰型,最高峰值位于5月,次峰值出现于8—9月(图1)。
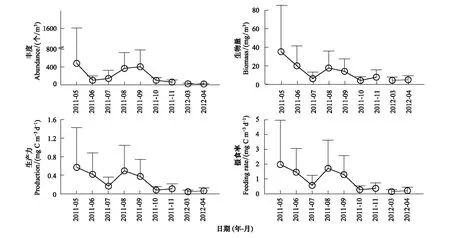
图1 莱州湾强壮滨箭虫丰度、生物量和生产力、摄食率的月变化Fig.1 Monthly mean values of abundance, biomass, and production, feeding rate of Aidanosagitta crassa in Laizhou Bay图中只显示正向标准偏差
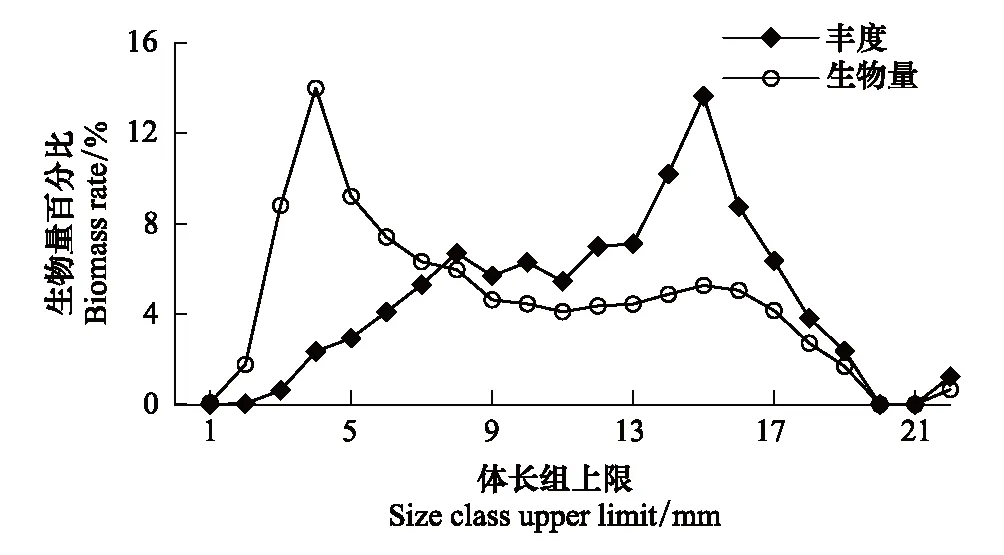
图2 莱州湾强壮滨箭虫各体长组的丰度和生物量百分比分布 Fig.2 Percentage contributions of size-classified individuals to the total Aidanosagitta crassa abundance and biomass in Laizhou Bay
各体长组中,对丰度贡献最多的是4mm体长组,其次为3mm和5mm体长组,6—18mm体长组的相对丰度介于4%—8%;对生物量贡献最大的是15mm体长组,其次为14mm体长组个体。<2mm以及>19mm体长组个体的丰度及生物量相对比例都小于2%(图2)。
2.2 体长分布
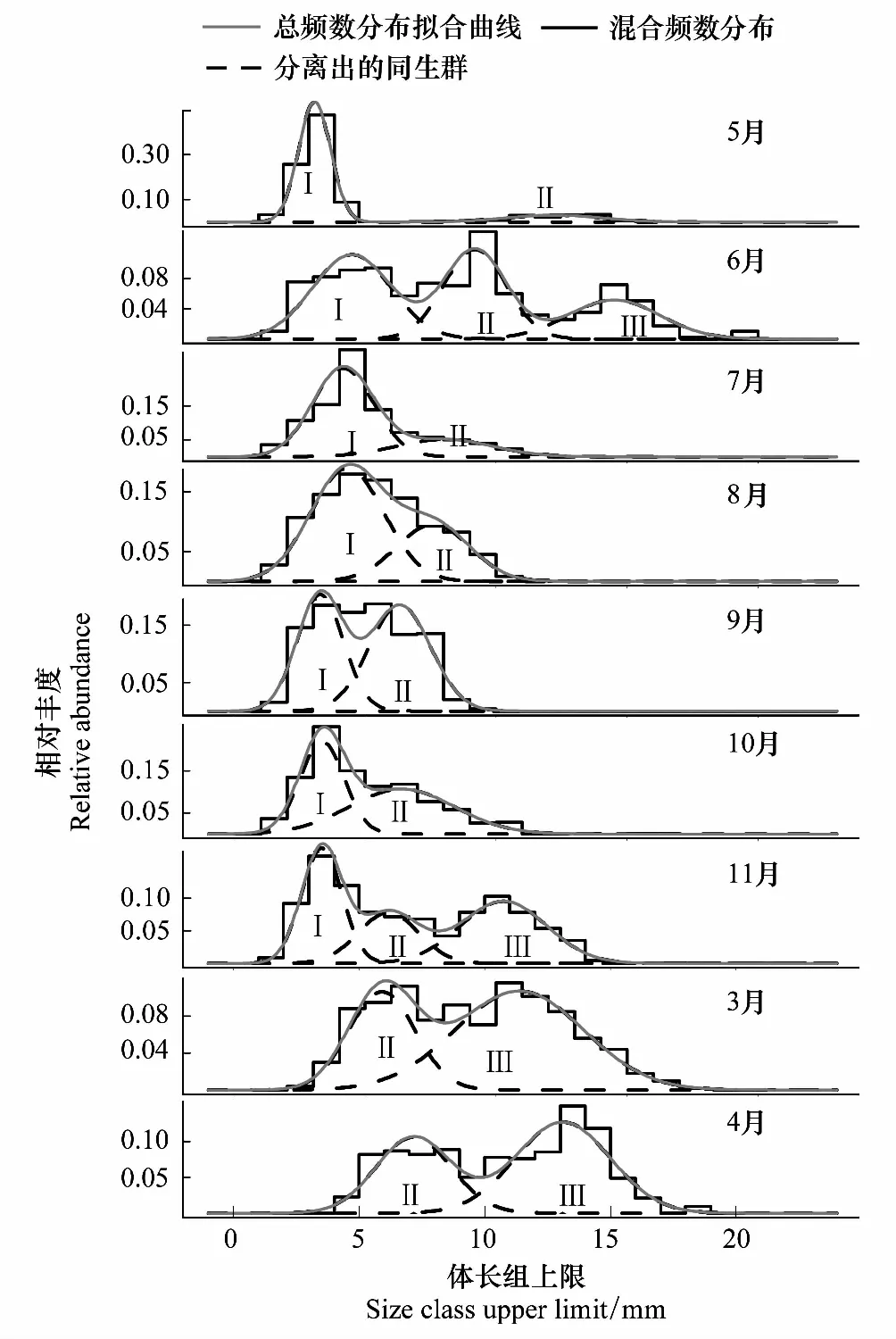
图3 莱州湾强壮滨箭虫的体长-频数相对分布 Fig.3 Mixed body length-abundance normal distributions of Aidanosagitta crassa in Laizhou Bay
强壮滨箭虫平均体长的月变化表现为,3—4月最高、分别为9.15 mm和10.80mm,其次为6月(8.34mm)和11月(7.19mm),5月和9月最低、小于5mm,其他月的平均体长介于5—5.5mm。
>10mm的个体多出现于11月至6月、极少或基本不出现于7—10月。7—10月,个体的体长分布上限阈值多在11mm以内。各月的体长频数分布可以分辨出3个平均体长分别为<5mm(I)、5—10mm(II)和>10mm(III)的3个同生群(Cohort)。6月和11月存在上述3个同生群,7—10月出现Cohort I和II,3月和4月出现Cohort II和III,5月出现Cohort I和III(图3)。
为方便分析3个同生群的平均体长和丰度的月变化,图4中将2012年3月和4月的位置调至2011年5月之前。3—6月间,3个同生群的体长呈增加趋势,之后开始下降,其中以Cohort II最为明显,9月各同生群的体长不再明显下降(图4)。Cohort I和Cohort III都以5月丰度最高,Cohort I在8月还有一个丰度次高峰;Cohort II以6月和9月丰度最高(图4)。
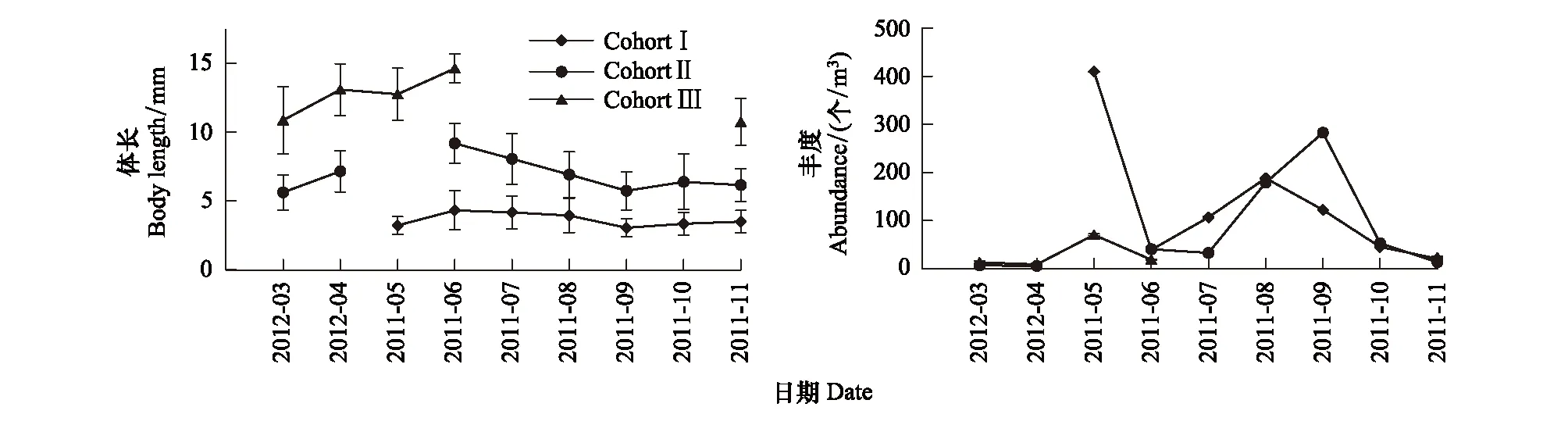
图4 强壮滨箭虫体长-相对频数混合分布的同生群所对应的月平均体长和丰度Fig.4 Monthly mean body length and abundance of size-classified Aidanosagitta crassa cohorts from mixture distributions results

图5 莱州湾强壮滨箭虫体长组丰度(个/m3)相对组成平面分布Fig.5 Spatial distributions and size-specific compositions of Aidanosagitta crassa abundance (ind/m3) in Laizhou Bay
2.3 丰度和生物量的平面分布
5月,强壮滨箭虫高丰度(>4000 个/m3)出现在莱州湾的湾顶、近黄河口侧,主要由<5mm个体贡献所致;从湾顶向湾口,丰度逐渐降低,>10mm个体是丰度贡献主体。此月,生物量贡献者主要是>10mm个体,在湾顶和湾口东侧还有一定比例的<5mm和5—10mm个体(图5和图6)。6月,湾内各站丰度和生物量明显降低于5月;丰度和生物量相对高值区在湾内沿岸分布;>10mm个体是生物量构成主体;湾口外东侧和湾内沿岸>10mm、5—10mm个体相对较多(图5和图6)。7月丰度和生物量分布中心位于湾西侧,<5mm和5—10mm个体分别是丰度和生物量的构成主体(图5和图6)。8月和9月湾内丰度主要由<5mm和5—10mm个体组成,5—10mm个体是多数站生物量的主要贡献者(图5和图6);<5mm个体在8月和9月分别是在湾中部(图5)和湾内站位(图6)有较高丰度。10月总丰度持续减少,湾内仍以<5mm和5—10mm个体为丰度构成主体,>10mm个体在湾西侧站位相对比例略高(图5和图6)。11月,分布退缩至湾内少数站,>10mm个体是生物量构成主体(图5和图6)。3—4月,分布范围扩至湾口,但丰度较低(<50个/m3),>10mm个体仍为生物量的构成主体(图5和图6)。
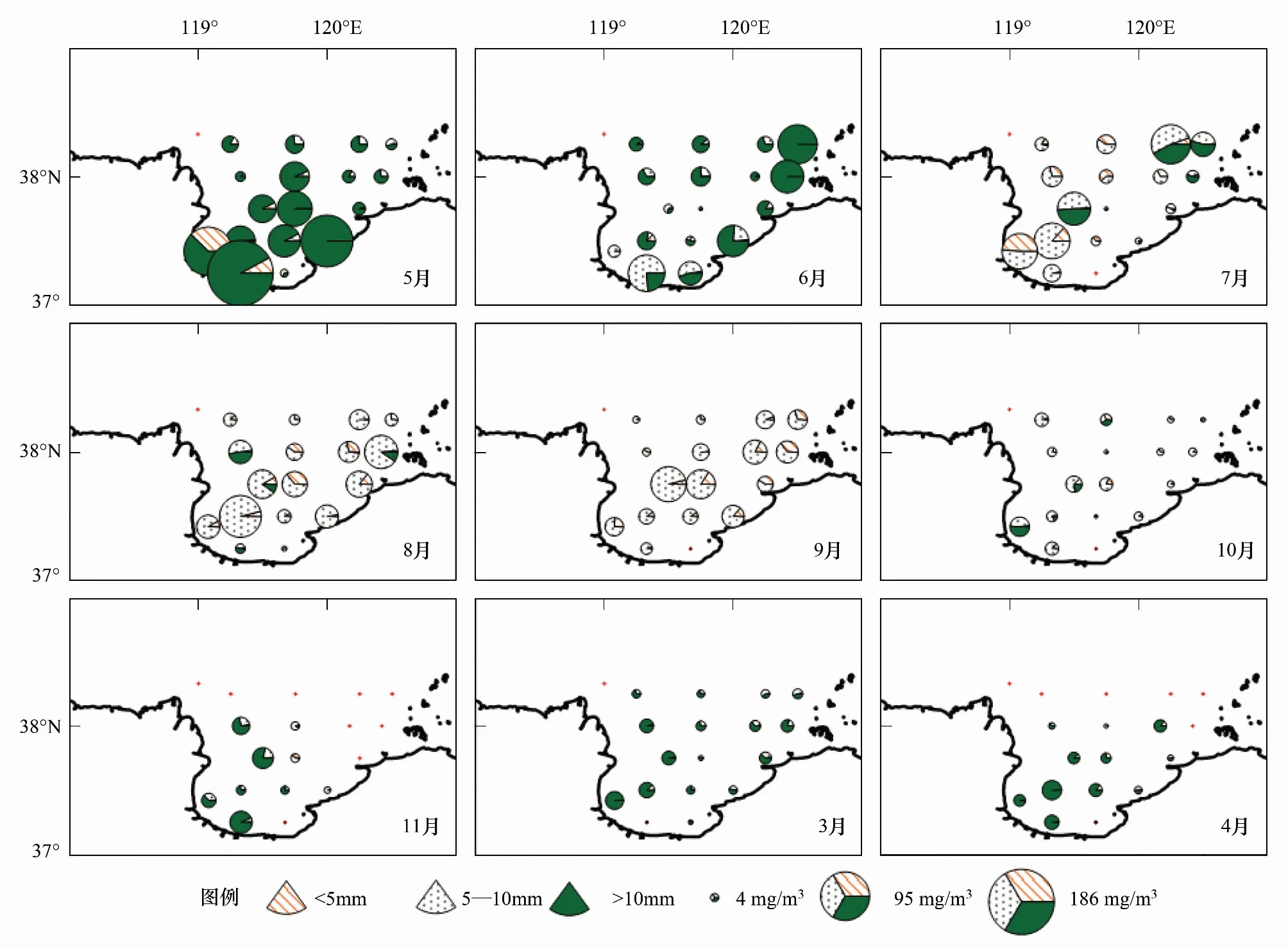
图6 莱州湾强壮滨箭虫体长组生物量(mg/m3)相对组成平面分布Fig.6 Spatial distributions and size-specific compositions of Aidanosagitta crassa biomass(mg/m3)in Laizhou Bay
体长>10mm和<5 mm的强壮滨箭虫高丰度区出现于5月水温介于15—17℃、盐度介于29—32的站位(图7a,7c);>10 mm的个体基本不出现(<1 个/m3)7—9月间水温大于24℃的站位(<1 个/m3)(图7c);<5 mm的个体基本不出现于3—4月以及11月内水温低于11℃的站位(图7a);5—10 mm个体在各调查月的绝大数站位都有分布,其高丰度区是8月水温介于26—27℃、盐度30—32的部分站位(图7b)。各调查站中,水温高于17℃时,强壮滨箭虫的平均体长介于4—6 mm;水温低于17℃时,平均体长介于8—12 mm;平均体长>10 mm的等值线仅出现于水温<20℃的区域(图7d)。

图7 莱州湾强壮滨箭虫不同体长组的丰度(个/m3)、各调查站平均体长(mm)与表层水温(℃)和盐度的相对分布Fig.7 Abundance and mean body length distributions of Aidanosagitta crassa in related to the surface temperature (℃) and salinity in Laizhou Baya.体长<5mm个体丰度;b. 体长介于5—10 mm个体丰度;c. 体长>10 mm个体丰度;d.各调查站所有个体的平均体长
强壮滨箭虫的丰度和体长与水温表现明显相关关系。表层水温与强壮滨箭虫中>10 mm个体的丰度和生物量呈显著负相关(P<0.05),与5—10mm个体丰度、总丰度及总生物量呈显著正相关(P<0.05);平均体长与表层水温、盐度都呈显著负相关(P<0.05)(表1)。
表1莱州湾强壮滨箭虫丰度、生物量和平均体长与表层水温、盐度相关性分析
Table1Correlationanalysisbetweentheabundance,biomassandmeanbodylengthofAidanosagittacrassaandsurfacewatertemperatureandsalinityinLaizhouBay
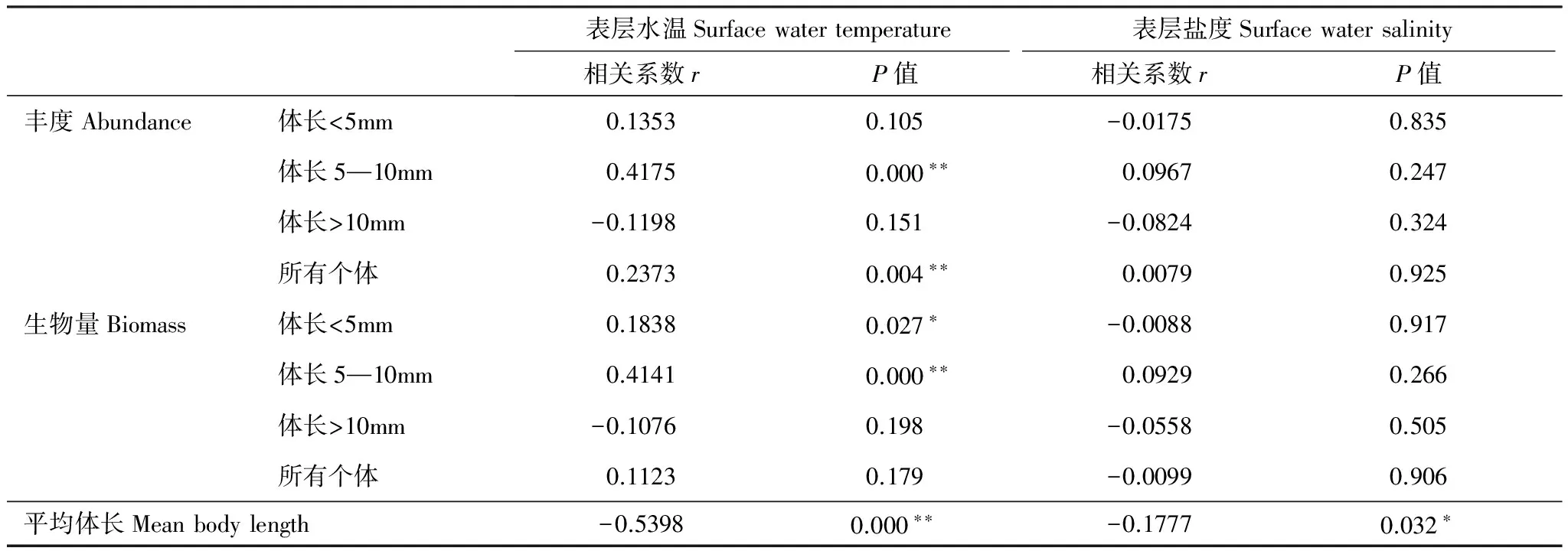
表层水温Surfacewatertemperature表层盐度Surfacewatersalinity相关系数rP值相关系数rP值丰度Abundance体长<5mm0.13530.105-0.01750.835体长5—10mm0.41750.000∗∗0.09670.247体长>10mm-0.11980.151-0.08240.324所有个体0.23730.004∗∗0.00790.925生物量Biomass体长<5mm0.18380.027∗-0.00880.917体长5—10mm0.41410.000∗∗0.09290.266体长>10mm-0.10760.198-0.05580.505所有个体0.11230.179-0.00990.906平均体长Meanbodylength-0.53980.000∗∗-0.17770.032∗
*P<0.05;**P<0.01
3 讨论
3.1 体长频数分布
强壮滨箭虫的体型季节更替明显。根据泡状组织在身体的分布延展程度,强壮滨箭虫大致可分为个体较大的冬季型、个体较小的内海型(囊开型或夏季型)以及介于前两者之间的中间型。内海型在春末-夏季孳生旺盛、成体体长7—10mm;冬季型适低温、多在冬春繁殖,成体体长13—20mm[10]。本研究虽未对强壮滨箭虫进行分型,但图7d和图3中显示,莱州湾强壮滨箭虫夏、秋季(7—10月)个体小、平均体长小于5mm;冬、春季(11—6月)个体大,绝大多数大于10 mm的强壮滨箭虫个体出现于此期。这与烟台外海[20]、胶州湾[14,18]、日本濑户内海[11- 12]等的强壮滨箭虫体型季节变化模式基本相符。
种群体长—频数分布是进行种群世代分析的基础。Nagasawa[13]和霍元子等[14]等曾根据体长频数粗略估计了日本东京湾和胶州湾外强壮滨箭虫的世代数。强壮滨箭虫存在多种生态型,单个生态型都能发育成熟、实现完整的生命周期;而且强壮滨箭虫可以自体或异体受精,一次受精后产卵可持续数周[28],卵孵化只需1—4d[12],也即是某个产卵群体可以在较长的时间内,支持其种群持续增长。因此,特定时期内强壮滨箭虫种群不同生态型、世代间的体长频数分布存在着高度重叠。要准确推算世代数,需要基于高频观察的体长参数、发育期(年龄)组成、性腺成熟度以及产卵等数据。Murakami曾结合每周观察到的发育期组成和体长分布,分离出日本濑户内海强壮滨箭虫4个世代[12]。本研究中逐月体长频数分布(图3),反映了强壮滨箭虫的种群结构季节变动基本特点,但由于采样时间间隔较大,单个同生群中可能包含多种生态型,因而并不能据之准确估测莱州湾强壮滨箭虫种群周年世代数。
无论是内海型或冬季型,强壮滨箭虫成体体长都大于5 mm[11-12]。本文中分离出的Cohort I的平均体长介于3—5 mm(图4),可视为未成熟个体或补充群体。该同生群亦是春季5月、夏季7—8月和秋季10月强壮滨箭虫体长频数混合分布的重心分布所在(图3)。此三季强壮滨箭虫的体长频数分布接近金字塔型年龄锥体[32],据之可认为上述3个季节为强壮滨箭虫种群增长和补充期。3—4月,体长频数分布重心移至Cohort II和III,偏向于壶形锥体[32],处于种群丰度下降期。6月,3个同生群相对组成均衡,体长频数分布偏向于钟形锥体[32],处于种群丰度相对稳定型。冬季冰期12月至2月未进行调查,期间湾内的水温低,不可能形成适宜强壮滨箭虫种群快速增长的外部条件。因而认为,莱州湾的强壮滨箭虫种群补充期应在5—10月间,晚于日本濑户内海(2—11月)[12]和黄海(3—10月)[14]。
3.2 水温和盐度对强壮滨箭虫体型和丰度分布的影响
强壮滨箭虫体长与水温呈明显的负相关[12,14],生长和体型受水温变化控制,中间型个体遇低温时可继续生长发展成为冬季型较大个体,而不进入产卵期[12]。在渤海冬季型强壮滨箭虫在4月占主导,中间型在9—10月以及1月占主导,内海型在5—8月以及11—12月占主导[10]。图3中,Cohort III平均体长大于10 mm,应主要包含强壮滨箭虫冬季型和过渡中间型个体,但该组群并不周年存在。10月至11月湾内平均水温由17℃降至10.6℃,Cohort III开始出现,5月至6月平均水温由12℃升至19℃,6月之后Cohort III消失,11月和6月是其开始出现和消失的转折期,此两季莱州湾月间水温差最大(与前月水温差>7℃)。在东京湾也曾发现3—4月水温8.3—15℃、12月水温12—13℃时,强壮滨箭虫体型呈现明显的夏、冬季型更替[13]。胶州湾外,强壮滨箭虫体长也是在水温由13℃升至17℃的5—6月间出现大幅变化[14]。已报道的培养实验发现,冬季型的强壮滨箭虫能存活至17℃[12]。本研究中显示,15—17℃可将平均体长>10mm和<10mm个体分布区分开(图7d)。综上推测,强壮滨箭体型变化或更替的水温阈值应不超过15—17℃,主要更替季节为6月和11月。
强壮滨箭虫被认为是近岸广盐种和较喜低温的广温种[23]。虽然本文结果中显示强壮滨箭虫盐度耐受范围广,但它表现一定的低盐偏好习性,其高丰度往往位于等盐线33以低一侧(图7a—c)。强壮滨箭虫的相对密集区也基本与莱州湾低盐近岸水季节分布动态相适。受季风、黄河冲淡水和外海水共同作用,春季(3—5月)莱州湾水温低,黄河冲淡水向南及东南方、湾底扩散,强壮滨箭虫数量激增,高密区位于湾南侧底部;夏季(6—8月)水温升高,黄河入海径流受莱州湾顺时针环流和东南季风影响、向东或东北向流出,外海水以补偿形式流入湾内,导致湾底特别是东侧湾底盐度反而较高,此季强壮滨箭虫的数量和生物量分布中心明显向湾中西侧偏移;冬季(11—2月)沿岸黄河冲淡水向莱州湾底输送积聚、形成冬季湾底清晰动力学低盐区,湾底水温较高、盐度较低,湾口处水温低、盐度高,强壮滨箭虫则退缩于湾内,基本不出现于湾口。
3.3 生物量和生产力估算
5月和10月莱州湾强壮滨箭虫的生产力远高于渤海湾南部相应值(0.27 mgC m-2d-1、0.14 mgC m-2d-1)[22]。3月胶州湾外强壮滨箭虫生物量峰值(411.10 mg/m2)高于莱州湾的5月峰值。如果将未调查的12—2月取11—4月间低值,莱州湾强壮滨箭虫的年生产力和摄食量估算值分别为1.058g C m-2a-1和2.8 g C m-2a-1,略高于胶州湾外强壮滨箭虫年生产力(0.67 g C m-2a-1)和摄食率(2.33 g C m-2a-1)[14]。若以莱州湾水域面积为7000 km2、海洋生物含C量约为干重的40%[33]、体内含水量86—89%[34]计,莱州湾内强壮滨箭虫全年需要摄食生物的湿重大约为350—490 t。
致谢:本文海上样品采集和处理、分析得到王秀霞、陈峻峰、吴强、孙坚强、陈瑞盛以及农业部黄渤海渔业资源环境重点野外科学观测试验站工作人员的帮助,特此致谢。
[1] 白雪娥, 庄志猛. 渤海浮游动物生物量及其主要种类数量变动的研究. 海洋水产研究, 1991, (12): 71- 92.
[2] 彭荣, 左涛, 万瑞景, 林群, 李显森, 王俊, 金显仕. 春末夏初莱州湾浮游动物生物量谱及潜在鱼类生物量的估算. 渔业科学进展, 2012, 33(1): 10- 16.
[3] 林景祺, 杨纪明. 烟台、威海和青岛沿岸当年生鲐鱼幼鱼的摄食习性. 海洋水产研究, 1980, (1): 1- 15.
[4] 杨纪明, 李军. 渤海强壮箭虫摄食的初步研究. 海洋科学, 1995, (6): 38- 42.
[5] Nomura H, Aihara K, Ishimaru T. Feeding of the chaetognathSagittacrassaTokioka in heavily eutrophicated Tokyo Bay, Japan. Plankton and Benthos Research, 2007, 2(3): 120- 127.
[6] Fukuhara O, Fukunaga T. Predation by Sagitta on larval fish in earthen pond. Bulletin of the Nansei Regional Fisheries Research Laboratory, 1984, 17: 151- 153.
[7] Jin X S, Shan X J, Li X S, Wang J, Cui Y, Zuo T. Long-term changes in the fishery ecosystem structure of Laizhou Bay, China. Science China Earth Sciences, 2013, 56(3): 366- 374.
[8] 杨尧尧, 李忠义, 吴强, 王俊. 莱州湾渔业资源群落结构和多样性的年际变化. 渔业科学进展, 2016, 37(1): 22- 29.
[9] 张波, 金显仕, 吴强, 谢周全. 莱州湾中国明对虾增殖放流策略研究. 中国水产科学, 2015, 22(3): 361- 370.
[10] 萧贻昌. 中国动物志 无脊椎动物 第三十八卷 毛颚动物门 箭虫纲. 北京: 科学出版社, 2004: 54- 56.
[11] Hirota R. On the morphological variation ofSagittacrassa. Journal of the Oceanographical Society of Japan, 1959, 15(4): 191- 202.
[12] Murakami A. Marine biological study on the planktonic chaetognaths in the Seto Inland Sea. Bulletin of Naikai Regional Fisheries Research Laboratory, 1959, 12: 1- 186.
[13] Nagasawa S. Vertical distribution, life cycle and production of the chaetognathSagittacrassain Tokyo Bay, Japan. Journal of Plankton Research, 1991, 13(6): 1325- 1338.
[14] 霍元子, 孙松, 杨波. 南黄海强壮箭虫(Sagittacrassa)的生活史特征. 海洋与湖沼, 2010, 41(2): 180- 185.
[15] 戴燕玉. 南黄海和东海毛颚类生态特征的研究I. 数量分布的特征. 海洋学报, 2006, 28(6): 106- 111.
[16] 盖建军, 倪金俤. 海洲湾强壮箭虫的生态学特征研究. 生物学杂志, 2012, 29(6): 59- 61.
[17] 毕洪生, 孙松, 高尚武, 张光涛, 张芳. 渤海浮游动物群落生态特点III. 部分浮游动物数量分布和季节变动. 生态学报, 2001, 21(4): 513- 521.
[18] 王倩, 孙松, 霍元子, 杨波. 胶州湾毛颚类生态学研究. 海洋与湖沼, 2010, 41(4): 639- 644.
[19] 陈峻峰, 左涛, 王秀霞. 南黄海浮游动物主要种类数量分布年间比较. 海洋学报, 2013, 35(6): 195- 203.
[20] 郑重, 郑执中, 王荣, 林雅蓉, 高尚武. 烟、威鲐鱼渔埸及邻近水域浮游动物生态的初步研究. 海洋与湖沼, 1965, 7(4): 329- 354.
[21] 李云, 徐兆礼, 高倩. 长江口强壮箭虫和肥胖箭虫的丰度变化对环境变暖的响应. 生态学报, 2009, 29(9): 4773- 4780.
[22] 张晓举, 温若冰, 于海洋. 渤海湾南部水域春秋季强壮滨箭虫摄食压力变化的初步研究. 海洋通报, 2013, 32(4): 434- 439.
[23] 刘青, 曲晗, 张硕, 魏杰. 强壮箭虫摄食生态的实验研究. 水产学报, 2006, 30(6): 767- 772.
[24] 刘青, 刘芳, 朱海燕, 胡浩. 光照、体长对强壮箭虫摄食强度和日粮的影响. 海洋湖沼通报, 2009, (1): 155- 161.
[25] 刘青, 曲晗, 张硕. 强壮箭虫对温度、盐度的耐受性研究. 海洋湖沼通报, 2007, (1): 111- 116.
[26] 刘青, 朱海燕, 刘芳, 丁子元. 温度、盐度对强壮箭虫耗氧率和窒息点的影响. 应用生态学报, 2011, 22(11): 3081- 3086.
[27] 于浩, 刘光兴, 陈洪举. 3种浮游生物网对强壮箭虫捕获效率的比较研究. 海洋学报, 2015, 37(10): 141- 145.
[28] Nagasawa S. Laboratory feeding and egg production in the chaetognathSagittacrassaTokioka. Journal of Experimental Marine Biology and Ecology, 1984, 76(1): 51- 65.
[29] Ikeda T. Metabolic rates of epipelagic marine zooplankton as a function of body mass and temperature. Marine Biology, 1985, 85(1): 1- 11.
[30] Macdonald P D M, Pitcher T J. Age-groups from size-frequency data: a versatile and efficient method of analyzing distribution mixtures. Journal of the Fisheries Research Board of Canada, 1979, 36(8): 987- 1001.
[31] 冯秋园, 孙晓霞, 任琳琳. 几种保存方法对中华哲水蚤和强壮箭虫体型的影响. 海洋科学, 2014, 38(4): 20- 26.
[32] 孙儒泳, 李庆芬, 牛翠娟, 娄安如. 基础生态学. 北京: 高等教育出版社, 2002: 64- 69.
[33] Uye S. Length-weight relationships of important zooplankton from the Inland Sea of Japan. Journal of the Oceanographical Society of Japan, 1982, 38(3): 149- 158.
[34] Omori M. Weight and chemical composition of some important oceanic zooplankton in the North Pacific Ocean. Marine Biology, 1969, 3(1): 4- 10.
SeasonaldistributionofbodylengthandabundanceofAidanosagittacrassa(Tokioka,1938)inLaizhouBay,BohaiSea
ZUO Tao1, 2,3,*, WANG Jun1, 2,4, SHI Yongqiang1, 2, 3
1YellowSeaFisheriesResearchInstitute,ChineseAcademyofFisherySciences,Qingdao266071,China2LaboratoryforMarineEcologyandEnvironmentalSciences,QingdaoNationalLaboratoryforMarineScienceandTechnology,Qingdao266200,China3KeyLaboratoryforSustainableDevelopmentofMarineFisheries,MinistryofAgriculture,Qingdao266071,China4ShandongProvincialKeyLaboratoryofFisheryResourcesandEcologicalEnvironment,Qingdao266071,China
Aidanosagittacrassa(Tokioka, 1938) is the only dominant chaetognatha species in Laizhou Bay, Bohai Sea. This gelatinous species plays an important role in the ecosystem of Laizhou Bay, acting as the major carnivorous predator of small invertebrate and fish larva. In addition, it has different morphs responding to variations in environmental factors. To determine the population structure in Laizhou Bay, the detailed body size and abundance ofA.crassawere analyzed monthly from 147 vertical net samples collected from May 2011 to April 2012 (excluding December 2011 and January and February 2012, because of the presence of sea ice). Results showed thatA.crassaoccurred continuously and was distributed extensively throughout the survey seasons and area in the bay. Both the abundance and biomass of the species presented two peaks, a considerable peak in May, and a second peak during August to September. Individuals with a body length of about 4 and 15 mm were the major contributors to the total abundance and biomass, respectively. The production of theA.crassapopulation was estimated to be 0.25 mg C m-3d-1, whereas the feeding rate was 0.88 mg C m-3d-1during the survey months. The body size ofA.crassafluctuated seasonally, with the largest sizes in March and April (mean body length > 10 mm), and the smallest in May and November (mean body length < 5 mm). The mean body length of the species showed an inverse correlation with both surface water temperature (SWT) and surface water salinity (SWS) of the corresponding period. Based on length-frequency distributions, different types of population pyramids occurred, namely, the expanding type in May to November except in June, stationary type in March to April, and constrictive type in June. The growing season for theA.crassapopulation in Laizhou Bay was presumed to be from May to August, because of the presence of abundant small individuals. Moreover, from the monthly length-frequency distributions, three common normalized cohorts (I, II, III) were detected with mean lengths of <5, 5—10, and >10 mm, respectively. Regarding their spatial distributions, individuals of cohorts III and I were the most abundant at sites with an SWT of 15—17℃ and SWS of 29—32 during spring. However, cohort III showed a considerable negative correlation between its abundance and water temperature, with a limited abundance at warm sites with an SWT > 24℃. Additionally, cohort I showed a positive correlation between its abundance and water temperature, and showed a limited abundance at cold sites with an SWT < 11℃. Cohort II was most abundant at sites with an SWT of 26—27℃ and SWS of 30—32 during autumn. From above the results, it was assumed that the water temperature threshold was lower than 15—17℃ for the transformation between the two typical size morphs, i.e., the large winter and small summer forms ofA.crassa.
Aidanosagittacrassa; body length; abundance; production; seasonal distribution;Laizhou Bay
国家基金委-山东省联合基金项目(U1406403);国家重点基础研究发展规划项目(2011CB403604);公益性行业(农业)科研专项(201303050-01);山东省自然科学基金(2009ZRB02317)
2016- 04- 26; < class="emphasis_bold">网络出版日期
日期:2017- 03- 22
10.5846/stxb201604260788
*通讯作者Corresponding author.E-mail: zuotao@ysfri.ac.cn
左涛,王俊,时永强.莱州湾强壮滨箭虫种群数量及个体大小季节分布.生态学报,2017,37(15):5189- 5197.
Zuo T, Wang J, Shi Y Q.Seasonal distribution of body length and abundance ofAidanosagittacrassa(Tokioka, 1938) in Laizhou Bay, Bohai Sea.Acta Ecologica Sinica,2017,37(15):5189- 5197.
