贡湖湾退圩还湖区水位高程下植被分布格局与土壤特征
2017-09-01张森霖卢少勇陈方鑫毕润成
张森霖,卢少勇,陈方鑫,毕润成
1 山西师范大学生命科学学院, 临汾 0410002 中国环境科学研究院,环境基准与风险评估国家重点实验室,国家环境保护湖泊污染控制重点实验室,湖泊环境研究中心,国家环境保护洞庭湖科学观测研究站,湖泊工程技术中心,北京 1000123 中国地质大学(北京)水资源与环境学院,北京 1000834 山西师范大学生命科学学院,临汾 041000
贡湖湾退圩还湖区水位高程下植被分布格局与土壤特征
张森霖1,卢少勇2,*,陈方鑫3,毕润成4
1 山西师范大学生命科学学院, 临汾 0410002 中国环境科学研究院,环境基准与风险评估国家重点实验室,国家环境保护湖泊污染控制重点实验室,湖泊环境研究中心,国家环境保护洞庭湖科学观测研究站,湖泊工程技术中心,北京 1000123 中国地质大学(北京)水资源与环境学院,北京 1000834 山西师范大学生命科学学院,临汾 041000
为揭示水陆交错带水位高程下植被分布格局、多样性指数和土壤特征的变化,于2015年10月对贡湖湾湿地北部生态修复区不同水位高程下植物群落组成和土壤特征进行调查,结果表明:整个研究区有165种植物,隶属65科、142属,多为多年生植物且受人为影响严重;随水位高程增加,植物群落R、H、D、J呈现逐渐递增趋势,具体表现为常年水淹区G1<季节性水淹区G2<水淹区G3;土壤容重及机械组成中粉、黏粒含量随水位高程升高而增加;随土壤营养物渐增其植物群落多样性增加,多样性和优势度指数与土壤全氮(TN和有机质(OM)负相关。OM含量呈“V”型先减后增分布;TN呈逐渐增多的趋势;速效钾(EP)含量无明显波动,约为13.50 mg/kg;有效磷(AP)含量先减少后基本不变;4个多样性指数间正相关;TN与OM显著正相关(R2=0.533);OM与EP负相关(R2=-0.144;TN与丰富度、多样性、优势度正相关;AP与4个多样性指数均正相关;pH值与4个多样性指数负相关;回归分析可见,TN和OM是影响植物群落多样性的关键因子。科学的植被管理、全面调控土壤营养盐含量有助于水陆交错带植被恢复。
水陆交错带;水位高程;区系组成;生物多样性;土壤
水陆交错带(land/water ecotones, LWEs),是指内陆水生与陆地生态系统间的功能界面区,是阻止污染物入湖的最后屏障,水陆交错带植物群落和土壤作为其中两个重要组成部分。土壤是植物群落的主要环境因子之一,土壤理化性质和种子库的特征影响植被发展和演替进程,也受植被演变影响,土壤性质与植物群落组成及生物多样性密切相关[1-2],摸清二者间的关系尤为重要,也是生态学研究热点之一。许多学者已开展大量野外调查和模拟研究,不同放牧退化程度下植被群落和土壤养分盐之间相关性显著[3- 5];对森林、山地植被群落和土壤理化性质的研究表明,不同植被森林植被类型和土壤理化性质相互依存、相互制约[6- 8];沙地、荒漠表明沙漠化过程中土壤-植被关系依赖于空间尺度,不同土层深度及土壤类型对土壤理化性质都有较大的影响和关联[9- 11];过渡施肥对植物多样性和生态系统功能有负面影响,植物群落的功能组成和养分输入土壤过程之间有着显著的相互作用[12-14]。目前,多数研究集中在自然地貌中森林、山地、草原和荒漠等植被与土壤间的相互作用,鲜见对湖泊退圩还湖区基底与生态重建后交错带植被和土壤的定量研究。
贡湖是无锡市和苏州市的重要水源地,随着近年来太湖水质恶化,水环境现状堪忧,水华威胁太湖周边城市生产、生活和生存[15]。2011年对贡湖湾北部进行退圩还湖生态修复,主要利用地表径流拦截、入湖河口污染负荷消减、岸堤改造和水动力改善、水陆交错带植被重建与基底改造、沉水植物群落恢复等技术共同搭建生态修复区[16-21]。本文以典型退圩还湖后人工生态修复的水陆交错带为例,分析在水文过程下交错带植物群落多样性和土壤特征,为同类型交错带重建提供参考。
1 材料与方法
1.1 研究区概况
研究区位于贡湖湾北部退圩还湖生态修复区(图1a),总占地2.32 km2,西起许仙港东至小溪港,北临通锡高速,南与太湖相隔,介于31°26′53.19″—31°27′56.89″N, 120°19′40.08″—120°20′48.31″ E,区内交错带全长约10 km。年均气温16.2℃,年均无霜期超过240 d,积温5418℃,降水量1121.7 mm,年日照百分率46%。
研究区修复前为鱼塘,生态修复措施包括圩湖成垸(太湖引水,圩堤隔断,闸门放水但目前不与太湖连通);水陆交错带植被重建;基底重建、堤岸构建(淤泥干化构建基底、人工岛、岸堤);地表径流拦截污染拦截与净化利用;沉水植物群落恢复等方法。根据近4年来监测的水位变化趋势,1—5月基本保持在3.3 m(吴淞水位,下同)以下,6—10月保持高水位3.5—4.3 m,11—12月逐步回落到3.3 m以下。

图1 贡湖湾水陆交错带地理位置、采样点分布图Fig.1 Geographical position and sampling points in the land/inland water ecotone of Gonghu Lake
1.2 样品采集与处理
1.2.1 植物调查与样品采集
据生态学植物群落调查规范[22]及常年检测水位变化,将生态修复区内新、老大堤间的交错带按水位高程和植被类型分为3个梯度带(图2):常年水淹区G1(2.8—3.3 m)、季节性水淹区G2(3.3—4.3 m)、微水淹区G3(>4.3 m);各带内设4块样地,各样地内设3个10 m×10 m乔木样方;各乔木样方内设灌木/草本样方2个5 m×5 m/4个1 m×1 m,采样点分布图见图1b,其中G1红色、G2蓝色、G3绿色,黑色实心原点代表采样点位置。
2015年10月完成植被调查与土壤取样。统计各样方内的:乔木(种名、株高、胸径、冠幅)、灌木(种名、丛高、盖度、高度)和草本(种名、数量、平均高度、盖度)[23]。用彼得森抓泥斗和铁铲联合采土,装袋标号,带回实验室待测。用环刀和铝盒取土,重复3个,带回实验室测量土壤容重和机械组成及营养盐含量。

图2 贡湖湾水陆交错带不同水位高程结构示意图Fig.2 Structure diagram of different water level in the land/inland water ecotone of Gonghu LakeG1:草带 Grass zone,G2:灌草带 Shrub and grass zone,G3:乔灌草带 Tree shrub and grass zone
1.2.2 测定方法
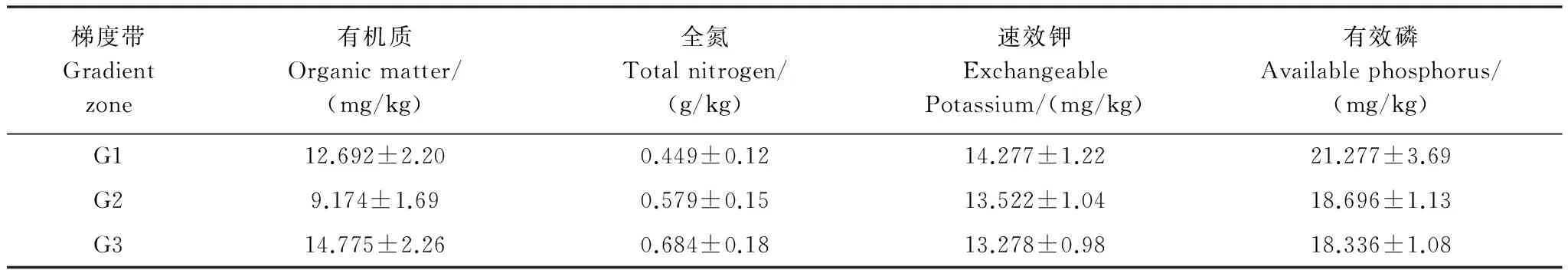
容重用环刀比重测定法、机械组成用比重计法、总氮(TN)用凯氏定氮法、总磷(TP)用酸溶-钼锑抗比色法、pH值用酸度计法、有机质(OM)用低温外加热重铬酸钾氧化-比色法、速效钾(EP)用中性乙酸铵提取火焰光度计法测定[24]。
1.3 数据处理
1.3.1 重要值
乔木重要值=(相对频度+相对多度+相对优势度)/3
灌、草本重要值=(相对盖度+相对高度)/2
1.3.2 物种多样性计算
用R语言中vegan包算生物多样性指数,其中R为物种数、H为Shannon-wiener指数、D为Simpson指数、J为Pielou均匀度指数。多样性、相关分析、回归分析等数据用Excel软件处理,用R软件计算生物多样性指数和相关性分析、回归分析。
1.3.3 群落多样性测度
因各层植物群落对生态环境贡献不同,按群落垂直结构特点对其不同生长型多样性指数加权[25],其公式如下[26-27]:
A=W1A1+W2A2+W3A3[27]
式中,A为群落总体多样性指数;A1,A2,A3依次为乔、灌、草3层多样性指数;W1,W2,W3依次为乔、灌、草3层多样性指数加权参数。算得W1=0.4483,W2=0.1988,W3=0.3529。统一多样性测度后的加权得到植物群落生物多样性指数。
2 结果与分析
2.1 交错带内植物区系组成及其多样性指数
2.1.1 交错带内植物群落组成
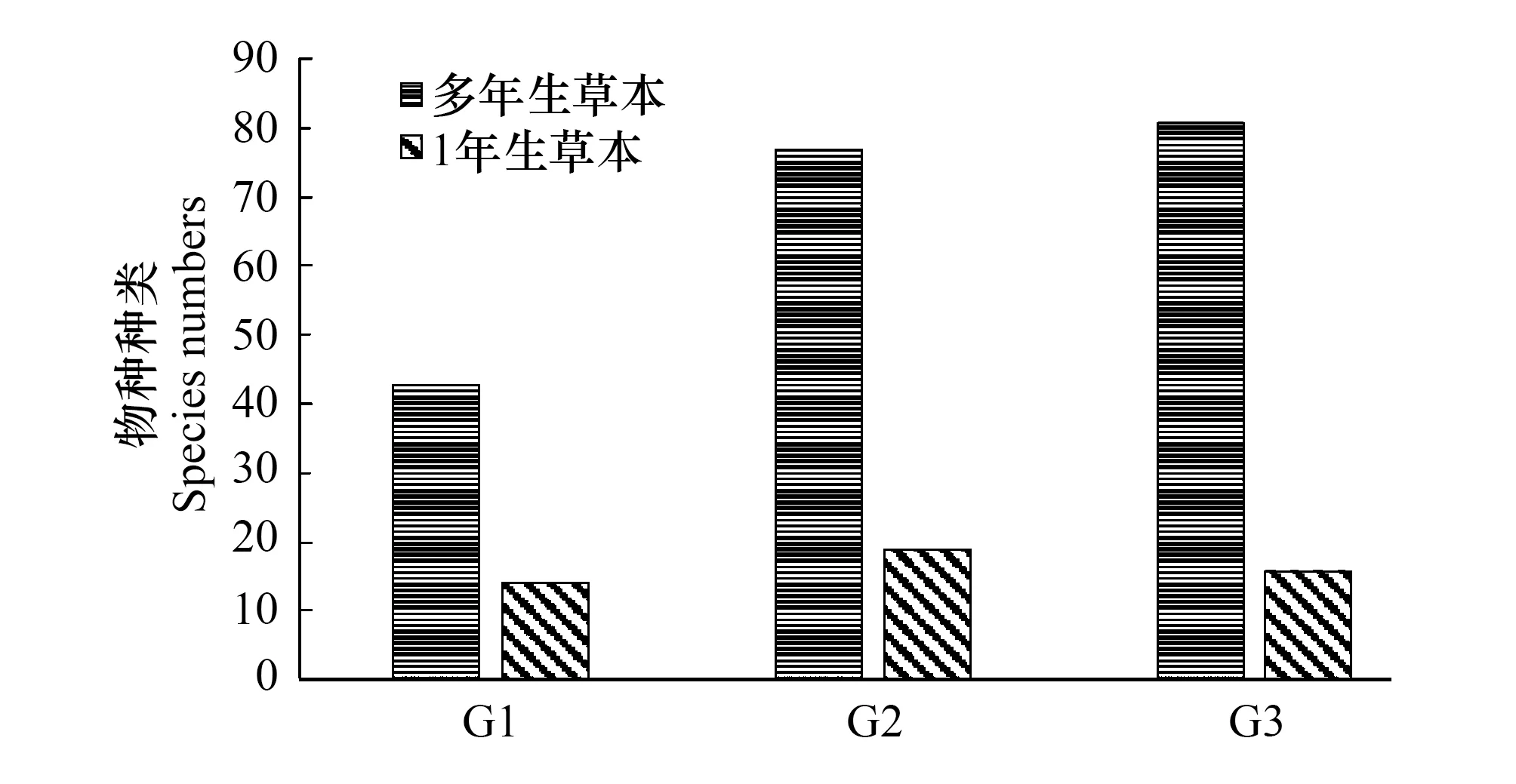
图3 不同水位高程植物组成分布Fig.3 Species distribution at different water level G1,草带 Grass zone,G2,灌草带 Shrub and grass zone,G3,乔灌草带 Tree shrub and grass zone
本调查中共出现165种植物,隶属65科、142属(附表1)。由图3可见,3个梯度带中多年生植物均多于1年生植物,是因研究区大部分植物人工配置,可快速提高植物多样性组成及满足造景功能,但与自然演替结果有别[28],也与国内外相关研究结论有别[29- 31]。调查表明仅有11中自然演替出的新物种(附表1中斜体)。1年生草本生长周期短,繁殖力强,在水陆交错带水文作用影响下也能较短时间完成其生命周期,依靠土壤种子库或其他种子源重新开始新的生长循环。而多年生草本则需较长时间完成生命循环,更依赖目前人为管理。可知本区域的植物群落组成还未稳定化,加之研究区本身是开放式公园景区,随着游人渐多,影响植物群落的发展和演替[32- 34],若人为干预撤销,生物多样性大幅降低。
2.1.2 水陆交错带内植物多样性指数变化
在研究区调查植物中,乔木出现15科、19属、21种;灌木出现14科、23属、26种;草本出现44科、92属、114种。可见草本层对植物群落多样性贡献最大,灌木层第二,乔木层最少。除5科12属21种草本植物外,其余均为人工种植。其中主要乔木有墨西哥落羽杉(Taxodiummucronatum)、池杉(Taxodiumdistichumvar.imbricatum)、樟(Cinnamomumcamphora);主要灌木有石楠(Photiniaserratifolia)、日本晚樱(Cerasusserrulatavar.lannesiana)、杨梅(Myricarubra);主要草本有铜钱草(Hydrocotylevulgaris)、芦竹(Arundodonax)、金鸡菊(Coreopsisdrummondii)、大滨菊(Leucanthemummaximum)等。高程增至最高水位后,植被类型从单一草本群落变为出现了乔、灌木群落。
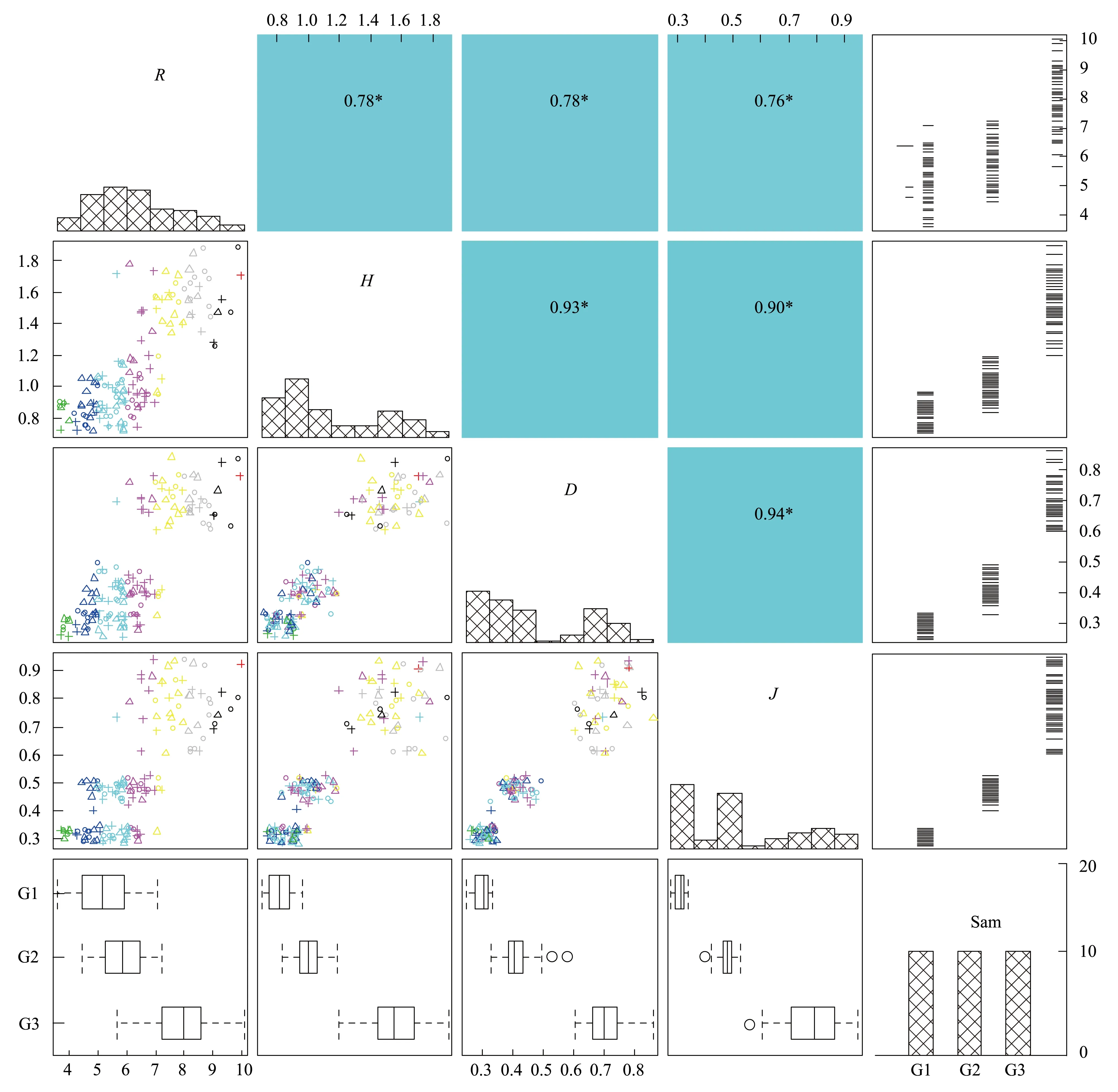
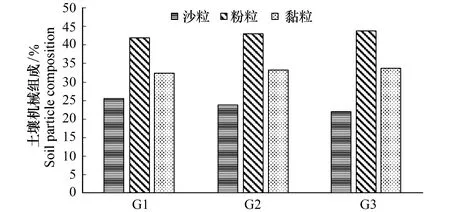
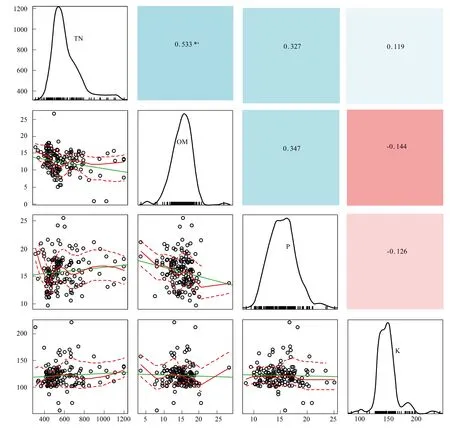
生物多样性反映着一片群落的结构、复杂度、功能特性和物质能量循环途径,是生态系统稳定的充要条件。由各梯度带多样性指数变化(图4)可见,随水位高程增加,群落物种丰富度、多样性指数、优势度和均匀度都呈明显升势。H表现为G1(0.824) 图4 不同水位高程下的植物多样性指数Fig.4 Vegetation diversity indexes at various water levelsR:物种数 Species;H:Shannon-winner指数 Shannon-winner index;D: Simpson指数Simpson index;J: 均匀度指数 Pielou index 2.1.3 物种多样性指数相关矩阵分析 R,H,D,J各指数间的相关矩阵图(图5)中,下半矩阵为各指数散点图,主对角线分别指示4指数变化趋势。上半矩阵分别显示了各相关分析结果。最后一行是各指数盒须图,最右一列为数据分布图。可见4指数间两两呈正相关(*P<0.05)。多样性指数和优势度指数及优势度指数和均匀度指数间极显著。即其中一个指数升高,其余3个指数升高。 结合盒须图和数据分布图可见,G1,G2,G3带的4个多样性指数区别较明显。其中,各梯度的4个多样性指数中,G3分布最广,为R(5.65—10.07),H(1.20—1.90),D(0.60—0.86),J(0.61—0.95);G1,G2,G3的4个指数均呈正偏态分布;G3中位数(R:7.95,H:1.55,λ:0.70,J:0.79)最大,其次是G2,最后是G1。各梯度落在各指数的50%值,R:G1(4.45—5.89),G2(5.25—6.48),G3(7.21—8.60);H:G1(0.75—0.88),G2(0.39—0.44),G3(1.45—1.68);λ:G1(0.28—0.32),G2(0.39—0.44),G3(0.66—0.74),J:G1(0.29—0.32),G2(0.46—0.49),G3(0.71—0.86)。 图5 物种多样性指数相关性矩阵图 (Pearson)Fig.5 Correlative Matrix plot of diversity indexes (Pearson) 蓝色为正相关,红色为负相关(本分析中未出现负相关)颜色越深相关度越大,反之越小;箱须图两条须子分别表示最小值和最大值,Sam表示样方;*P<0.05 2.2 不同梯度带土壤理化性质 2.2.1 不同梯度带土壤物理性质差异 土壤容重是反映土壤透水性、孔隙度、含水量及植物根系生长阻力等特性的综合指标[20]。表1所示为研究区G1,G2,G3带土壤容重和植被盖度变化。土壤容重呈随水位高程增加而增大的趋势,植被盖度也上升,结合多样性指数,可见植物群落组成趋于更复杂和高等。 土壤机械组成对土壤物理、化学、生物特性具决定作用,不同土壤机械组成有别,矿物组成差异大,其化学和其它性质均不同,以氮、磷、钾对植物生长发育影响为最[35]。比较G1,G2,G3带土壤机械组成(图6)可知,土壤颗粒以粉粒为主,质量分数分别为G1(41.72%),G2(42.00%),G3(44.82%)。粉、黏粒含量分别为沙粒的2.76,2.93,3.37倍。 按我国标准[36]土壤机械组成中沙、粉、黏粒划分标准依次为:1—0.05 mm,0.05—0.005 mm<0.005 mm。总体上,沙粒含量随高程增而降,而黏、粉粒含量均升。是因随水位高程增加,土壤受水流冲刷减弱。反之,土壤黏、粉粒含量下降,而含沙量增高。除水流侵蚀影响外,植物对土壤机械组成的影响不可忽视,土壤粉、黏粒颗粒易吸附OM和营养盐,随水位高程增加,粉、黏粒含量增加,植物多样性增高,植物群落趋于复杂,开始出现大型乔、灌木,继而改良土壤机械成分,使土壤颗粒分布均匀,改变土壤透气性和持水性,与有关研究一致[37-38]。 表1 不同高程梯度带土壤类型和植被特征 G1,草带 Grass zone,G2,灌草带 Shrub and grass zone,G3,乔灌草带 Tree shrub and grass zone 图6 不同水位高程的土壤机械组成Fig.6 Soil particle composition at different water levels G1:草带 Grass zone,G2:灌草带 Shrub and grass zone,G3:乔灌草带 Tree shrub and grass zone 2.2.2 不同梯度带土壤化学性质的差异 土壤OM是湿地植物最主要营养源,其含量直接影响生产力,而研究区内土壤特征受来自湖水水位不同频率和波动的影响,故使OM成为气候变化的敏感指示物,能指示湿地对气候变化的响应[39-40]。由表2可见OM随水位高程增加呈“V”型,G2带含量最低,因土壤中OM含量动态变化,其输入(有机残体腐化)和输出(生物和非生物因素)会影响其含量,G1带水位较低,与湖水交换最频,来自湖水中动、植物残体在岸边腐化因素明显高于G2和G3带,G3带中来自水文的因素大减,但随大量乔木群落出现,枯枝落叶层出现,增加了其OM输入量,且其生物多样性指数较高,植物群落组成更复杂,增强了其对OM的持留,故OM含量较高。 自然土壤氮主要来自动植物残体和生物固氮,少量来自大气降水。土壤氮输出主要是OM分解,大部分被植物吸收利用,部分有机氮经矿化(氨化)、硝化和反硝化及氨挥发而重返大气[41]。土壤氮含量将直接影响植物群落多样性、植物生产力等。表2中TN含量随水位高程增加呈增势,G3带TN含量约是G1的1.5倍,综合其植物多样性指标可知,更丰富多样物种组成可有效提高固氮微生物反应速率,调节各形态氮转换效率。 植物所需钾主要来自土壤,但除EP外绝大部分钾不能被植物直接吸收[42]。与氮不同,土壤钾含量受母质影响较大,本研究中G1,G2,G3带中EP含量无明显差异,均约13mg/kg因交错带在生态修复改造时坡岸所用堆土来源一致,成土母质性质相近。 表2 不同梯度带有机质、全氮、速效钾、有效磷、pH值 G1:草带 Grass zone,G2:灌草带 Shrub and grass zone,G3:乔灌草带 Tree shrub and grass zone 植物生长除需氮、钾外,磷也必不可少,AP是可被植物利用的形态之一。过高磷含量不仅不能被植物有效利用,反而入湖促进富营养化。G1,G2,G3带AP含量分别为G1(21.277 mg/kg)>G2(18.696 g/kg)≈G3(18.336 mg/kg),可见水位较高的G2、G3带AP含量较接近,水位较低的G1带含量略高。因G1带在靠近水体边界的交错带,陆源输入在此得到富集和积累,并与水体交换最频,一年中被淹时间最长,且调查区岸边有大量湿生植物,本次调查接近秋末,已有植物残体浮于岸边,此乃G1带OM、EP、AP含量较高的原因之一。 3.2.3 土壤特征指标相关性分析 由图7可见OM和TN在0.01水平上显著正相关,对其回归分析得到方程:Y(TN,g/kg)=28730X2+250024(R2=0.884,表3),说明该交错带中有机氮对OM含量贡献很大。而OM和EP及AP和EP的相关性较差,R2分别为-0.027和-0.023。 图7 土壤特征指标相关性矩阵图(Pearson)Fig.7 Correlative Matrix plot of characteristic indexes of the soil(Pearson)TN:全氮、OM:有机质、P:有效磷、K:速效钾;下半矩阵为两两指标的散点图并添加拟合曲线及趋势线;上半矩阵为两两矩阵的相关关系及系数,蓝色为正相关,红色为负相关,颜色越深相关性越大;主对角线为核密度估计曲线;**表示在P<0.01(双尾)时显著相关 2.3 水陆交错带植物多样性指数和土壤理化因子相关关系 研究结果表明,植物多样性与土壤理化性质间明显相关。其中,TN与4个植物多样性指数均正相关,其中丰富度、多样性、优势度与TN在0.01水平上(双尾)显著正相关,均匀度显著性不高,这与许多研究一致[43-45];AP在0.05水平上(双尾)与丰富度、均匀度显著正相关;pH值与4个植物多样性指数均负相关,仅与丰富度在0.05水平上显著负相关;AP与4个植物多样性指数均正相关,其中与丰富度和均匀度显著相关;OM与各植物多样性指标相关性不明显,此现象可能因G1,G2,G3带生境多样性(光照、土壤机械组成、人为干扰程度等)不同,影响了群落结构组成和多样性。有效钾与植物多样性指数正相关,但相关性不显著。 表3 植物多样性指数和土壤理化因子相关关系(Pearson) R: 物种数 Species;H: Shannon-winner指数 Shannon-winner index;D: Simpson指数Simpson index;J: 均匀度指数 Pielou index 植物生长状况综合了多种生物和非生物因素因素,各因素相互制约,现通过对丰富度、多样性、优势度、均匀度与表中土壤各理化性质多元回归分析,先用多元方差分析,删除对Y无影响的变量,再逐步回归(表4)。可知TN和OM均出现在4个方程中,说明TN和OM对提高植物群落多样性影响显著;植物群落多样性提高后可有效提高土壤养分含量。另土壤pH值出现了3次,说明pH值对植物群落多样性有一定影响。 表4 群落植物多样性指数与土壤理化因子多元线性回归方程 R:物种数 Species;H:Shannon-winner指数 Shannon-winner index;D: Simpson指数Simpson index;J: 均匀度指数 Pielou index 贡湖湾湿地北部生态修复区自2011年开工建设,本研究首次探讨其植物群落多样性和土壤分异特征,经分析植物群落生物多样性和土壤特征,及所获结果相关性,主要结论如下: (1)贡湖湾湿地生态修复工程开工至今,从改造前植物多样性极低(均为杂草,无灌、乔木)到现在植物多样性大幅提高(R:7.914,H:1.549,D:0.703,J:0.780),与所用植物配置和维护管理有关;本次植物群落调查中,发现多年生草本数量是1年生草本的4.33倍,且自然演替的物种不足10%,且未发现明显指示种,因研究区植物群落长期处于人为干扰状态,日常植物管理严重干扰了植物群落自然状态下的生长和发育,自我调节和修复能力不足,一旦终止人为植物管理,极可能多样性明显下降。 (2)本研究按水位将研究区划分成3个梯度带:常年水淹区G1(2.8—3.3 m)、季节性水淹区G2(3.3—4.3 m)、微水淹区G3(4.3 m以上);随水位高程水位升高:容重增加G1(1.614 g/cm3) [1] 杨小波, 张桃林, 吴庆书. 海南琼北地区不同植被类型物种多样性与土壤肥力的关系. 生态学报, 2002, 22(2): 190- 196. [2] 安树青, 王峥峰, 朱学雷, 刘志礼, 洪必恭, 赵儒林. 土壤因子对次生森林群落演替的影响. 生态学报, 1997, 17(1): 45- 50. [3] 萨茹拉, 侯向阳, 李金祥, 丁勇, 吴新宏, 运向军. 不同放牧退化程度典型草原植被——土壤系统的有机碳储量. 草业学报, 2013, 22(5): 18- 26. [4] 王向涛, 张世虎, 陈懂懂, 谈嫣蓉, 孙大帅, 杜国祯. 不同放牧强度下高寒草甸植被特征和土壤养分变化研究. 草地学报, 2010, 18(4): 510- 516. [5] 谭红妍, 闫瑞瑞, 闫玉春, 陈宝瑞, 辛晓平. 不同放牧强度下温性草甸草原土壤微生物群落结构PLFAs分析. 草业学报, 2015, 24(3): 115- 121. [6] 陈亮中, 谢宝元, 肖文发, 黄志霖. 三峡库区主要森林植被类型土壤有机碳贮量研究. 长江流域资源与环境, 2007, 16(5): 640- 643. [7] 陈晨, 高明, 郑杰炳, 陈凌静, 李东. 缙云山不同森林植被下土壤理化性状研究. 西南大学学报: 自然科学版, 2010, 32(3): 88- 92. [8] 欧芷阳, 苏志尧, 袁铁象, 彭玉华, 何琴飞, 黄小荣. 土壤肥力及地形因子对桂西南喀斯特山地木本植物群落的影响. 生态学报, 2014, 34(13): 3672- 3681. [9] 杨玉海, 陈亚宁, 李卫红, 陈亚鹏. 伊犁河谷不同植被带下土壤有机碳分布. 地理学报, 2010, 65(5): 605- 612. [10] 孙永宁, 王进昌, 韩庆杰, 等. 青藏铁路格尔木至安多段沿线高寒植被、土壤特性与人工植被恢复研究. 中国沙漠, 2011, 31(4): 894- 905. [11] 陈玉福, 董鸣. 毛乌素沙地景观的植被与土壤特征空间格局及其相关分析. 植物生态学报, 2001, 25(3): 265- 269. [12] Mueller K E, Hobbie S E, Tilman D, Reich P B. Effects of plant diversity, N fertilization, and elevated carbon dioxide on grassland soil N cycling in a long-term experiment. Global Change Biology, 2013, 19(4): 1249- 1261. [13] De Vries F T, Jørgensen H B, Hedlund K, Bardgett R D. Disentangling plant and soil microbial controls on carbon and nitrogen loss in grassland mesocosms. Journal of Ecology, 2015, 103(3): 629- 640. [14] Strecker T, Barnard R L, Niklaus P A, Scherer-Lorenzen M, Weigelt A, Scheu S, Eisenhauer N. Effects of plant diversity, functional group composition, and fertilization on soil microbial properties in experimental grassland. PLoS One, 2015, 10(5): e0125678. [15] 赵凯, 李振国, 魏宏农, 张静, 马久远, 王国祥. 太湖贡湖湾水生植被分布现状(2012年). 湖泊科学, 2015, 27(3): 421- 428. [16] Zhang R B, Qian X, Zhu W T, Gao H L, Hu W, Wang J H. Simulation and evaluation of pollution load reduction scenarios for water environmental management: a case study of inflow river of Taihu Lake, China. International Journal of Environmental Research and Public Health, 2014, 11(9): 9306- 9324. [17] Lu S Y, Chen F X, Ngo H H, Guo W S, Feng C P,Wu J A, Zheng B H. Effect of Straw and Polyacrylamide on the Stability of Land/water Ecotone Soil and the Field Implementation. Ecological Engineering, 2016, 94: 12- 21 [18] 金晶, 张饮江, 黎臻, 卢少勇, 段婷, 翟斯凡, 郑天罡. 两种挺水植物强化底泥抗蚀效能研究. 安全与环境学报, 2013, 13(4): 88- 93. [19] 张文慧, 胡小贞, 许秋瑾, 赵阳. 湖泊生态修复评价研究进展. 环境工程技术学报, 2015, 5(6): 545- 550. [20] 张艳晴, 毕雪娟, 闫晖敏, 程花, 林超. 太湖地区贡湖湾生态修复区围隔系统消浪挡藻技术研究. 安徽农业科学, 2016, 44(2): 142- 145, 149- 149. [21] 陈方鑫, 吴文涛, 卢少勇, 薛巍, 吴纪南, 郑西强. 贡湖湾退渔还湖区的基底稳定性的时空特征. 环境工程学报, 2015, 9(6): 3024- 3030. [22] 方精云, 王襄平, 沈泽昊, 唐志尧, 贺金生, 于丹, 江源, 王志恒, 郑成洋, 朱江玲, 郭兆迪. 植物群落清查的主要内容、方法和技术规范. 生物多样性, 2009, 17(6): 533- 548. [23] 丁献华, 毕润成, 张慧芳, 闫明. 霍山七里峪植物群落的种类组成和生活型谱分析. 安徽农业科学, 2010, 38(22): 12032- 12033. [24] 中国土壤学会农业化学专业委员会. 土壤农业化学常规分析方法. 北京: 科学出版社, 1983: 44- 44, 55- 55, 68- 68. [25] 赵成章, 石福习, 董小刚, 任珩, 盛亚萍, 高福元, 杨文斌. 祁连山北坡退化林地植被群落的自然恢复过程及土壤特征变化. 生态学报, 2011, 31(1): 115- 122. [26] 徐远杰, 陈亚宁, 李卫红, 付爱红, 马晓东, 桂东伟, 陈亚鹏. 伊犁河谷山地植物群落物种多样性分布格局及环境解释. 植物生态学报, 2010, 34(10): 1142- 1154. [27] 高贤明, 黄建辉, 万师强, 陈灵芝. 秦岭太白山弃耕地植物群落演替的生态学研究. 生态学报, 1997, 17(6): 619- 625. [28] 王勇, 吴金清, 黄宏文, 刘松柏. 三峡库区消涨带植物群落的数量分析. 武汉植物学研究, 2004, 22(4): 307- 314. [29] 孙荣, 袁兴中, 刘红, 陈忠礼, 张跃伟. 三峡水库消落带植物群落组成及物种多样性. 生态学杂志, 2011, 30(2): 208- 214. [30] 王欣, 高贤明. 模拟水淹对三峡库区常见一年生草本植物种子萌发的影响. 植物生态学报, 2010, 34(12): 1404- 1413. [31] 王业春, 雷波, 张晟. 三峡库区消落带不同水位高程植被和土壤特征差异. 湖泊科学, 2012, 24(2): 206- 212. [32] 尹锴, 崔胜辉, 石龙宇, 吝涛, 郭青海, 吕静. 人为干扰对城市森林灌草层植物多样性的影响——以厦门市为例. 生态学报, 2009, 29(2): 563- 572. [33] Van der Veken S, Verheyen K, Hermy M. Plant species loss in an urban area (Turnhout, Belgium) from 1880 to 1999 and its environmental determinants. Flora-Morphology, Distribution, Functional Ecology of Plants, 2004, 199(6): 516- 523. [34] Carreiro M M, Howe K, Parkhurst D F, Pouyat P V. Variation in quality and decomposability of red oak leaf litter along an urban-rural gradient. Biology and Fertility of Soils, 1999, 30(3): 258- 268. [35] 李扬, 王冬梅, 信忠保. 漓江水陆交错带植被与土壤空间分异规律. 农业工程学报, 2013, 29(6): 121- 128. [36] 李天杰, 郑应顺, 王云. 土壤地理学(第2版). 北京: 高等教育出版社, 1983: 21- 21. [37] 唐炎林, 邓晓保, 李玉武, 张顺宾. 西双版纳不同林分土壤机械组成及其肥力比较. 中南林业科技大学学报: 自然科学版, 2007, 27(1): 70- 75. [38] 张保华, 何毓蓉, 周红艺, 程根伟. 长江上游典型区亚高山不同林型土壤的结构性与水分效应. 水土保持学报, 2002, 16(4): 127- 129. [39] 张燕, 彭补拙, 高翔, 唐翔宇, 杨浩. 人类干扰对土壤侵蚀及土壤质量的影响——以苏南宜兴低山丘陵区为例. 地理科学, 2002, 22(3): 336- 341. [40] 王丽, 胡金明, 宋长春, 杨涛. 水分梯度对三江平原典型湿地植物小叶章地上生物量的影响. 草业学报, 2008, 17(4): 19- 25. [41] Xiao H L. Climate change in relation to soil organic matter. Soil and Environment Science, 1999, 8(4): 300- 304. [42] Watson R T, Zinyowera M C, Moss R H. Climate Change 1995 Impacts: Adaptations and Mitigation of Climate Change: Scientific-Technical Analysis. Cambridge: Cambridge University Press, 1996. [43] 李小坤, 鲁剑巍, 吴礼树. 土壤钾素固定和释放机制研究进展. 湖北农业科学, 2008, 47(4): 473- 477. [44] 盛茂银, 熊康宁, 崔高仰, 刘洋. 贵州喀斯特石漠化地区植物多样性与土壤理化性质. 生态学报, 2015, 35(2): 434- 448. [45] 赵洋毅, 王玉杰, 王云琦, 赵占军, 刘楠, 陈林. 渝北不同模式水源涵养林植物多样性及其与土壤特征的关系. 生态环境学报, 2009, 18(6): 2260- 2266. Plant diversity and soil property changes in the restored Land/Water Ecotones of Gonghu bay ZHANG Senlin1, LU Shaoyong2,*, CHEN Fangxin3, BI Runcheng4 1 School of Life Science, Shanxi Normal University, Linfen 041000, China2 Engineering and Technology Centre of Lake, State Environmental Protection Scientific Observation and Research Station for Lake Dongtinghu, Research Centre of Lake Environment,State Environmental Protection Key Laboratory for Lake Pollution Control, State Key Laboratory of Environmental Criteria and Risk Assessment, Chinese Research Academy of Environmental Sciences, Beijing 100012, China3 China University of Geosciences(Beijing), Institute of Water Resources and the Environment, Beijing 100083, China4 School of Life Science,Shanxi Normal University, Linfen 041000, China The aim of the present study was to determine the variation in vegetation distribution patterns, diversity indices, and soil characteristics under water level elevations in land and water ecotones, in the northern part of Gonghu Bay in October 2015. Observing and records of phytocoenosium formations and soil features showed that the majority of all 165 species, subordinated to 142 categories and 65 families, were perennial plants that were seriously affected by human activities. The higher the water levels, the higher the abundance of plant communities (R,H,D,J) and were affected by the duration of submergence. The particle and silt proportions in soil bulk density and its formation increased with increasing water levels. The soil total nitrogen level and community richness also indicated a positive relationship. In contrast, the diversity and dominance of plant communities showed a negative relation with TN and organic matter (OM). The OM content in soil decreased from 12.69 to 9.14 mg/kg initially and then increased to 14.78 mg/kg, showing a “V” type distribution. The total nitrogen content in soil increased linearly from 0.45 to 0.684 g/kg-1. However, no obvious fluctuation was found in the available potassium (EP) content since it remained at 13.50 mg/kg, and the available phosphorus (AP) content decreased from 21.28 to 18.70 mg/kg and then remained unchanged. There was a positive correlation among the four diversity indices. Total nitrogen had a significantly positive relation with OM (R2= 0.533); OM had a negative correlation with EP (R2= -0.144), soil total nitrogen had a positive relationship with community richness, diversity, and dominance; available phosphorus had a positive relationship with the four diversity indices. The pH value had a negative relationship with the four diversity indices. The results of the regression analysis indicated that total nitrogen and OM are the key factors of vegetation diversity. Therefore, scientific vegetation management and comprehensive adjustment of nutrient salts are necessary to enhance the vegetation of the land/inland water ecotone. land/inland water ecotone; water level elevation; floral composition; diversity index; soil 附表1 研究区内植物物种统计表 G1:草带 Grass zone,G2:灌草带 Shrub and grass zone,G3:乔灌草带 Tree shrub and grass zone 国家水体污染控制与治理科技重大专项(2013ZX07101- 014,2012ZX07105-002); 国家自然科学基金(41373027);科技基础性工作专项重点项目(2015FY110900) 2016- 03- 13; 网络出版日期:2017- 02- 23 10.5846/stxb201603130446 *通讯作者Corresponding author.E-mail: lushy2000@163.com 张森霖,卢少勇,陈方鑫,毕润成.贡湖湾退圩还湖区水位高程下植被分布格局与土壤特征.生态学报,2017,37(13):4400- 4413. Zhang S L, Lu S Y, Chen F X, Bi R C.Plant diversity and soil property changes in the restored Land/Water Ecotones of Gonghu bay.Acta Ecologica Sinica,2017,37(13):4400- 4413.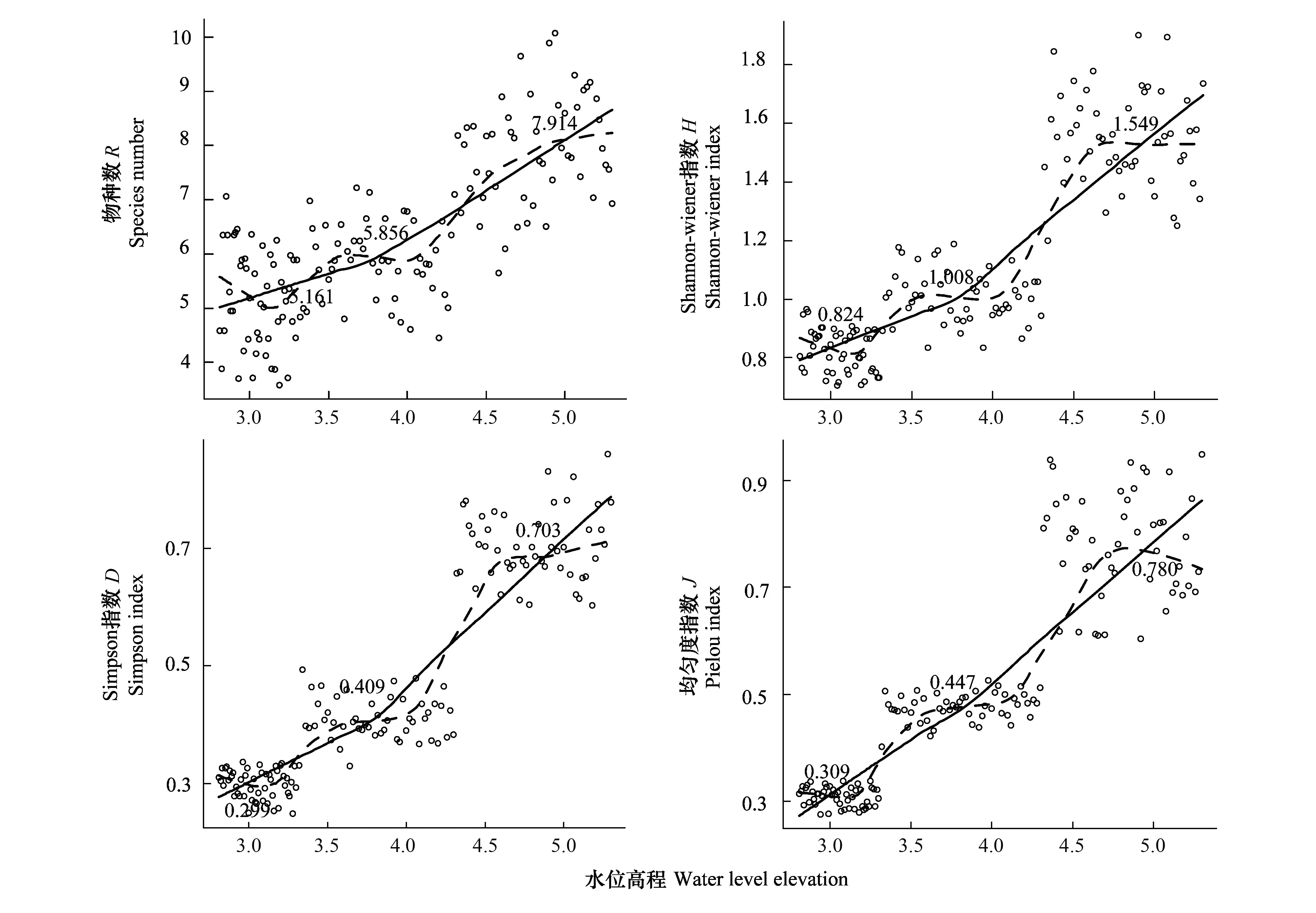



3 结论

