安徽黄山风景区微毛樱桃萌枝种群动态研究
2017-08-31尤禄祥陈志伟王华辰伊贤贵南程慧王贤荣
尤禄祥, 陈志伟, 王华辰, 伊贤贵, 南程慧, 朱 弘, 王贤荣,①
(1. 南京林业大学: a. 南方现代林业协同创新中心, b. 生物与环境学院, 江苏 南京 210037;2. 福建省漳州市林业局, 福建 漳州 363000; 3. 南京森林警察学院, 江苏 南京 210023)
安徽黄山风景区微毛樱桃萌枝种群动态研究
尤禄祥1a,1b, 陈志伟2, 王华辰1a,1b, 伊贤贵1a,1b, 南程慧3, 朱 弘1a,1b, 王贤荣1a,1b,①
(1. 南京林业大学: a. 南方现代林业协同创新中心, b. 生物与环境学院, 江苏 南京 210037;2. 福建省漳州市林业局, 福建 漳州 363000; 3. 南京森林警察学院, 江苏 南京 210023)
为探明安徽黄山风景区微毛樱桃〔Cerasusclarofolia(Schneid.) Yu et Li〕萌枝种群的生长规律和更新特点,在野外调查基础上,分别对该种群的萌枝类型、龄级结构〔由径级结构代替,根据胸径(DBH)和树高(H)划分径级〕、静态生命表、存活曲线和生存函数进行了分析。结果表明:该种群萌枝主要以根颈萌枝为主,根蘖萌枝较少,且多与根颈萌枝同时出现;幼龄萌枝数量较多、中龄萌枝数量较少、老龄萌枝数量更少,说明该种群的幼苗和幼树众多,而中龄和老龄植株却很少。该种群的萌枝数量变化动态指数在自然状态下为56.39%,而在考虑干扰时为5.64%。总体上看,Ⅰ级(DBH<3 cm,H<130 cm)至Ⅲ级(3 cm≤DBH<8 cm)萌枝的存活数和标准化存活数均较多,而Ⅷ级(28 cm≤DBH<33 cm)至Ⅹ级(DBH≥38 cm)萌枝的存活数和标准化存活数均较少;Ⅱ级(DBH<3 cm,H≥130 cm)萌枝的期望寿命最高(2.980),而Ⅹ级萌枝的期望寿命最低(0.500);各龄级萌枝的死亡率和损失度变化趋势基本一致,均有2个峰值,分别出现在Ⅰ级和Ⅸ级萌枝。该种群的存活曲线介于Deevey Ⅱ型和Deevey Ⅲ型之间,但更趋向Deevey Ⅱ型。Ⅰ级萌枝的死亡密度函数和危险率函数均最大,Ⅱ级萌枝的死亡密度函数和危险率函数均为0.000,其余龄级萌枝的死亡密度函数和危险率函数则趋于平缓;随龄级增大,萌枝的积累死亡率函数持续上升,而生存率函数却持续下降。上述结果表明:黄山风景区微毛樱桃萌枝种群总体上处于动态的稳定状态;从种群发展角度考虑,建议对该种群进行适当的小尺度的人为干扰,以改善幼苗的生存环境,促进种群更新。
微毛樱桃; 萌枝种群; 黄山风景区; 静态生命表; 存活曲线; 生存函数
萌枝是植物响应外界干扰的一种生存策略。具有萌枝能力的植物在遭受火灾、雪灾、飓风、泥石流、人工砍伐等干扰后能够在原地快速恢复生长,重新占领原来的空间资源,从而降低外界干扰对种群的影响,维持群落的稳定状态[1]。实际上,大多数植物都具有一定的萌枝能力,其中,裸子植物中具有萌枝能力的种类较少,而被子植物中蔷薇科(Rosaceae)和壳斗科(Fagaceae)具有萌枝能力的种类却较多,分别约占各科总种数的80%和78%[2-3]。在20世纪90年代,国内研究者开始关注植物萌枝方面的研究,目前,萌枝作为一种直接的植物再生方式越来越受到学者们的广泛关注[4-5]。
微毛樱桃〔Cerasusclarofolia(Schneid.) Yu et Li〕为蔷薇科樱属(CerasusMill.)落叶灌木或小乔木,主要分布在安徽、云南、贵州、湖北等省[6]。其中,安徽黄山风景区是微毛樱桃在华东地区最典型的分布点,多生于山坡林内、灌丛、林缘或路边等区域[7]。微毛樱桃的花期为4月至6月,果期为6月至7月;花瓣白色或粉红色,倒卵形至近圆形;花序伞形或近伞形,具花2~4朵,花叶同开;萼筒钟状,萼片卵状三角形或披针状三角形;核果红色,呈长椭圆形。实际上,微毛樱桃的花和果均具有极高的观赏价值。然而,实际调查结果[8]表明:受光照、雨雪、强风和地被物等自然环境条件的限制,安徽黄山风景区的微毛樱桃种子更新困难,实生苗存活率较低,主要靠萌枝方式进行繁殖和更新,从而降低其种群对种子更新的依赖程度,并保持其原来的生态位和竞争优势。迄今为止,国内外研究者对樱属植物萌枝特性的研究均较少,仅见野生欧洲甜樱桃〔C.avium(Linn.) Moench.〕萌枝方面的研究报道[9-11],而对微毛樱桃萌枝更新及种群动态变化的机制尚未深入研究,极大地限制了微毛樱桃的开发利用,并制约了其保护策略的制定。
鉴于此,在大量野外调查的基础上,采用样地调查法分别对安徽黄山风景区微毛樱桃萌枝种群的萌枝类型、龄级结构、静态生命表、存活曲线及生存函数进行了分析,以期明确黄山风景区微毛樱桃种群的生长规律和更新特点,并为微毛樱桃天然种群的保护及可持续开发利用提供参考资料。
1 研究区概况和研究方法
1.1 研究区概况
黄山风景区位于安徽省南部,具体地理坐标为东经118°01′~118°17′、北纬30°01′~30°18′,最高峰为莲花峰(海拔1 864.7 m)。该区域属亚热带季风气候,但山顶的年均温仅7.8 ℃,最冷月(1月份)均温-3.1 ℃,最热月(7月份)均温17.7 ℃,年均降水量2 398.5 mm;土壤类型主要为山地黄壤和黄棕壤,局部地区还发育有山地沼泽土和草甸土等。
安徽黄山风景区属于南、北植物区系交替的过渡带,为第四纪冰期动植物的避难所[12],植物区系古老、复杂、种类丰富,共有高等植物222科826属1 809种(含中国特有属22属)[7]。由于垂直高差显著、地形复杂,景区内植物群落自下而上依次为次生林、常绿阔叶林、常绿落叶阔叶混交林、落叶阔叶林、山地矮林、山地灌丛等。
相关调查结果[8]表明:微毛樱桃主要集中分布在黄山风景区内的落叶阔叶林(南坡海拔1 400 m,北坡海拔1 100~1 500 m)和山地矮林(海拔1 500~1 650 m)中,少数散布于黄山松(PinustaiwanensisHayata)林中;其分布的海拔下限位于北坡清凉台至松谷庵途中约海拔950 m处,其分布的海拔上限则位于莲花峰海拔1 830 m处(仅见零星分布)。伴生乔木种类主要有白檀〔Symplocospaniculata(Thunb.) Miq.〕、茅栗(CastaneaseguiniiDode)、四照花〔Cornuskousasubsp.chinensis(Osborn) Q. Y. Xiang〕、色木槭(AcermonoMaxim.)和黄山栎(QuercusstewardiiRehd.);伴生灌木种类主要有华箬竹(SasasinicaKeng)、中国绣球(HydrangeachinensisMaxim.)、三花悬钩子(RubustrianthusFocke)、半边月〔Weigelajaponicavar.sinica(Rehd.) Bailey〕和华空木(StephanandrachinensisHance)等;伴生草本植物和藤本植物主要有一把伞南星〔Arisaemaerubescens(Wall.) Schott〕、华东唐松草(ThalictrumfortuneiS. Moore)、灰背清风藤(SabiadiscolorDunn)、多花勾儿茶〔Berchemiafloribunda(Wall.) Brongn.〕和大芽南蛇藤(CelastrusgemmatusLoes.)等。
1.2 研究方法
根据安徽黄山风景区内微毛樱桃的分布特点和生物学特性,参照陈志伟[13]的样地和样线相结合的方法进行野外实地调查。在黄山风景区内,选择微毛樱桃分布较集中的地段设置9个面积20 m×20 m的样方,选择微毛樱桃植株密度较低的地段设置3条样线。参考方精云等[14]的方法,分别对样方内及样线上5 m范围内的木本植物进行每木调查,并详细记录每个样方的海拔、坡向、坡度等信息。由于微毛樱桃属于多茎干植物,其每个萌枝丛的茎干均源自同一主根,故视为一个个体(即基株,genet)。为了便于统计和分析,在实际调查过程中,将每个萌枝丛中胸径最大的茎干视为母株,其余茎干视为萌枝。认真统计每丛微毛樱桃的茎干数量,并测量萌枝丛中胸径小于3 cm的茎干高度及胸径大于3 cm的茎干胸径。按照Tredici[15]的萌枝分类系统对微毛樱桃萌枝种群的萌枝类型进行分析。
1.3 数据统计及分析
采用径级结构代替龄级结构的方法[16-17]来分析黄山风景区微毛樱桃萌枝种群的动态。根据微毛樱桃的生活史特点及研究目的,参考相关文献的径级划分标准,依据胸径(DBH)和树高(H)将萌枝龄级划分为10个等级:其中,DBH<3 cm的龄级有2个,包括Ⅰ级(H<130 cm)和Ⅱ级(H≥130 cm);DBH≥3 cm的龄级有8个,包括Ⅲ级(3 cm≤DBH<8 cm)、Ⅳ级(8 cm≤DBH<13 cm)、Ⅴ级(13 cm≤DBH<18 cm)、Ⅵ级(18 cm≤DBH<23 cm)、Ⅶ级(23 cm≤DBH<28 cm)、Ⅷ级(28 cm≤DBH<33 cm)、Ⅸ级(33 cm≤DBH<38 cm)和Ⅹ级(DBH≥38 cm)。分别统计各龄级萌枝的数量,据此编制微毛樱桃萌枝种群的静态生命表。
采用陈晓德[18]的方法对微毛樱桃萌枝种群结构进行动态量化分析,计算种群内各龄级萌枝数量变化动态指数(Vn)和自然状态下整个种群萌枝数量变化动态指数(Vpi),并计算考虑外部干扰时整个种群萌枝数量变化动态指数(Vpi′)。Vn和Vpi值为正数、负数和零时分别表示相邻龄级或整个种群个体数量的增长、衰退和稳定的动态关系。
参照洪伟等[19]的方法,根据微毛樱桃萌枝种群中不同龄级的萌枝数量编制静态生命表。采用EXCEL 2007软件绘制微毛樱桃萌枝种群的存活曲线,并选用Deevey Ⅱ和Deevey Ⅲ 2种数学模型对微毛樱桃萌枝种群的存活曲线类型进行检验和分析,其中,指数方程y=a·eb·x用于描述Deevey Ⅱ型存活曲线,幂函数方程y=a·xb用于描述Deevey Ⅲ型存活曲线[20]。
为了更好地阐明微毛樱桃萌枝种群的生存规律,参照杨凤翔等[20]的方法计算生存函数,包括死亡密度函数f(t)、危险率函数λ(t)、积累死亡率函数F(t)和生存率函数S(t)。
2 结果和分析
2.1 安徽黄山风景区微毛樱桃萌枝种群的萌枝类型分析
调查结果显示:安徽黄山风景区的微毛樱桃具有很强的萌枝能力,109株(丛)微毛樱桃的多干指数(multi-stemming index)达到93.6%;单株(丛)萌枝数量的最高值为62,平均值为13.4,中位数为9。微毛樱桃的萌枝类型主要有根颈萌枝和根蘖萌枝2种,其中,102株(丛)微毛樱桃存在根颈萌枝,而根蘖萌枝则不常见,且多与根颈萌枝同时出现。
2.2 安徽黄山风景区微毛樱桃萌枝种群的龄级结构分析
根据调查结果绘制的安徽黄山风景区微毛樱桃萌枝种群的龄级结构见图1。从图1可见:安徽黄山风景区微毛樱桃萌枝种群的龄级结构表现为幼龄萌枝数量较多、中龄萌枝数量较少、老龄萌枝数量更少,说明该微毛樱桃萌枝种群处于持续增长的状态。

根据胸径(DBH)和树高(H)划分龄级 According to diameter at breast height (DBH) and height (H) for dividing age class. Ⅰ: DBH<3 cm, H<130 cm; Ⅱ: DBH<3 cm, H≥130 cm; Ⅲ: 3 cm≤DBH<8 cm; Ⅳ: 8 cm≤DBH<13 cm; Ⅴ: 13 cm≤DBH<18 cm; Ⅵ: 18 cm≤DBH<23 cm; Ⅶ: 23 cm≤DBH<28 cm; Ⅷ: 28 cm≤DBH<33 cm; Ⅸ: 33 cm≤DBH<38 cm; Ⅹ: DBH≥38 cm.图1 安徽黄山风景区微毛樱桃萌枝种群的龄级结构Fig. 1 Age class structure of sprouting population of Cerasus clarofolia (Schneid.) Yu et Li in Huangshan Mountain Scenic Area of Anhui
该微毛樱桃萌枝种群结构的动态量化分析结果表明:Ⅰ级到Ⅱ级、Ⅱ级到Ⅲ级、Ⅲ级到Ⅳ级、Ⅳ级到Ⅴ级、Ⅴ级到Ⅵ级、Ⅵ级到Ⅶ级、Ⅶ级到Ⅷ级、Ⅷ级到Ⅸ级以及Ⅸ级到Ⅹ级的萌枝数量变化动态指数(Vn)分别为77.13%、-21.19%、55.82%、42.73%、44.44%、60.00%、50.00%、0.00%和85.71%。自然状态下整个种群萌枝数量变化动态指数(Vpi)为56.39%,说明该微毛樱桃萌枝种群属于增长型种群。在考虑外部干扰时,整个种群萌枝数量变化动态指数(Vpi′)为5.64%,明显高于0%,说明该微毛樱桃萌枝种群处于稳定状态。
2.3 安徽黄山风景区微毛樱桃萌枝种群的静态生命表分析
安徽黄山风景区微毛樱桃萌枝种群的静态生命表见表1。由表1可见:微毛樱桃萌枝种群的结构存在波动性。其中,Ⅰ级萌枝的存活数(ax)和标准化存活数(lx)最多(分别为857和1 000);Ⅱ级萌枝的ax和lx值显著减少(分别为196和229);Ⅲ级萌枝的ax和lx值明显增多(分别为249和291);在Ⅲ级至Ⅹ级间,随着龄级的增大,各龄级萌枝的ax和lx值呈逐渐减少的趋势;Ⅹ级萌枝的ax和lx值均为1。总体来看,Ⅷ级至Ⅹ级萌枝的ax和lx值较低,而Ⅰ级至Ⅲ级萌枝的ax和lx值却较高。
表1 安徽黄山风景区微毛樱桃萌枝种群的静态生命表1)
Table 1 Static life table of sprouting population ofCerasusclarofolia(Schneid.) Yu et Li in Huangshan Mountain Scenic Area of Anhui1)
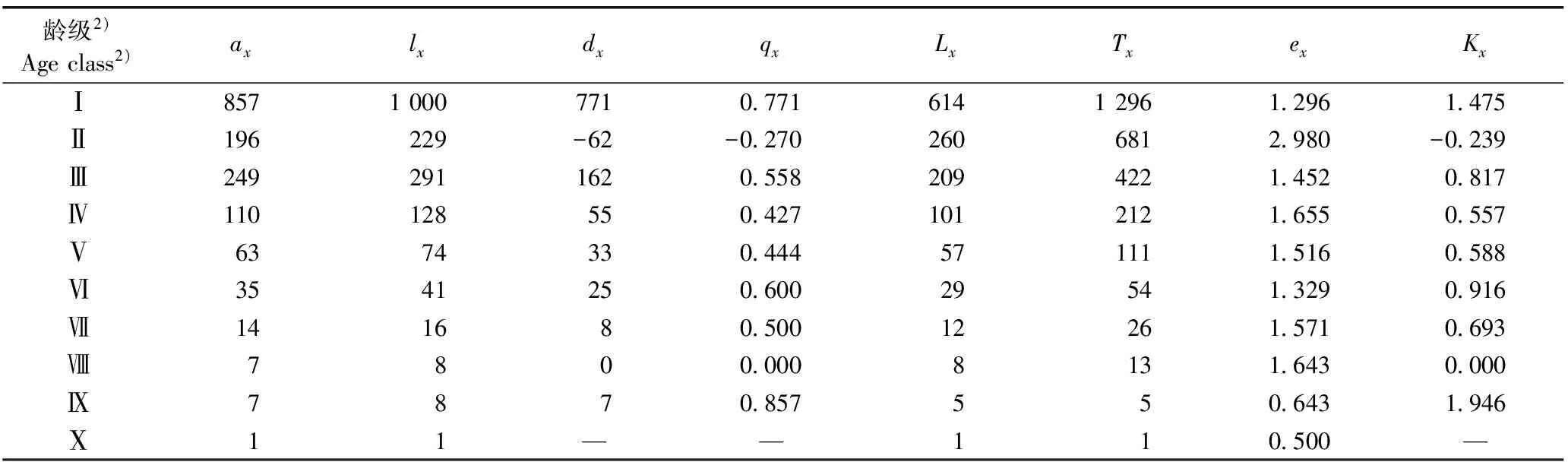
龄级2)Ageclass2)axlxdxqxLxTxexKxⅠ85710007710 77161412961 2961 475Ⅱ196229-62-0 2702606812 980-0 239Ⅲ2492911620 5582094221 4520 817Ⅳ110128550 4271012121 6550 557Ⅴ6374330 444571111 5160 588Ⅵ3541250 60029541 3290 916Ⅶ141680 50012261 5710 693Ⅷ7800 0008131 6430 000Ⅸ7870 857550 6431 946Ⅹ11——110 500—
1)ax:x龄级萌枝的存活数Survival number of sprouting branches atxage class;lx:x龄级萌枝的标准化存活数 Standardized survival number of sprouting branches atxage class;dx:x龄级到x+1龄级间萌枝的标准化死亡数 Standardized mortality number of sprouting branches fromxage class tox+1 age class;qx:x龄级到x+1龄级间萌枝的死亡率Mortality rate of sprouting branch fromxage class tox+1 age class;Lx:x龄级到x+1龄级间萌枝的存活数 Survival number of sprouting branches fromxage class tox+1 age class;Tx:x龄级及x以上龄级萌枝的存活总数 Total survival number of sprouting branches fromxage class to higher age class;ex:x龄级萌枝的期望寿命 Life expectancy of sprouting branch atxage class;Kx:x龄级萌枝的损失度 Loss degree of sprouting branch atxage class. —: 无对应理论值 No corresponding theoretical value.
2)根据胸径(DBH)和树高(H)划分龄级 According to diameter at breast height (DBH) and height (H) for dividing age class. Ⅰ: DBH<3 cm, H<130 cm; Ⅱ: DBH<3 cm, H≥130 cm; Ⅲ: 3 cm≤DBH<8 cm; Ⅳ: 8 cm≤DBH<13 cm; Ⅴ: 13 cm≤DBH<18 cm; Ⅵ: 18 cm≤DBH<23 cm; Ⅶ: 23 cm≤DBH<28 cm; Ⅷ: 28 cm≤DBH<33 cm; Ⅸ: 33 cm≤DBH<38 cm; Ⅹ: DBH≥38 cm.
由表1还可见:安徽黄山风景区微毛樱桃萌枝种群Ⅱ级萌枝的期望寿命(ex)最高(2.980),说明该龄级萌枝的生存质量最优,生理活动也很旺盛;Ⅰ级及Ⅲ级至Ⅷ级萌枝的ex值均较高,为1.296~1.655;Ⅸ级和Ⅹ级萌枝的ex值较低,分别为0.643和0.500,说明这2个龄级萌枝进入生理衰退阶段。
由表1还可见:安徽黄山风景区微毛樱桃萌枝种群各龄级萌枝的死亡率(qx)和损失度(Kx)变化趋势一致,均有2个峰值,分别出现在Ⅰ级和Ⅸ级萌枝,说明各龄级萌枝的qx和Kx值存在小周期波动。
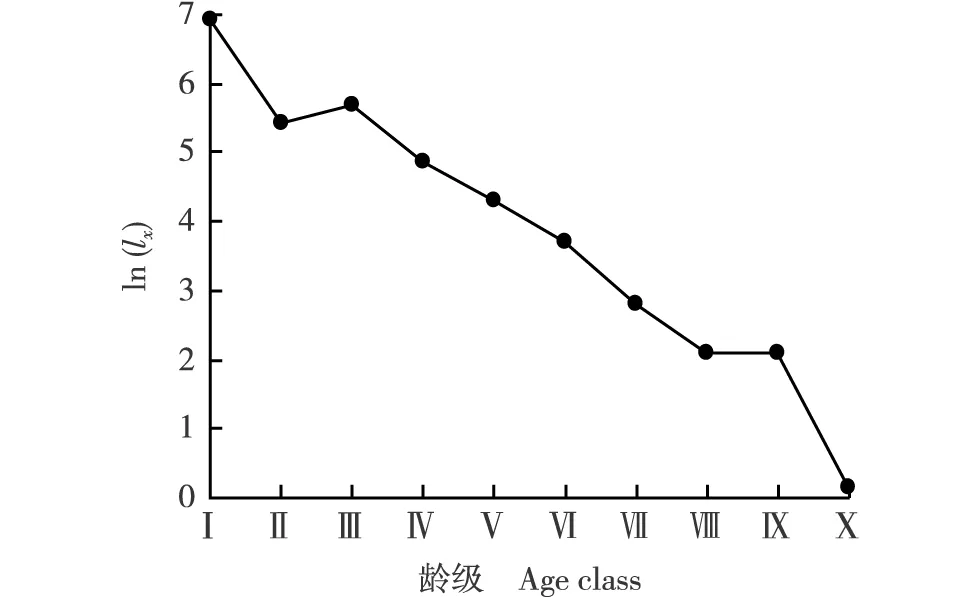
ln(lx): x龄级开始时萌枝标准化存活数的自然对数 Natural logarithm of standardized survival number starting from x age class. 根据胸径(DBH)和树高(H)划分龄级 According to diameter at breast height (DBH) and height (H) for dividing age class. Ⅰ: DBH<3 cm, H<130 cm; Ⅱ: DBH<3 cm, H≥130 cm; Ⅲ: 3 cm≤DBH<8 cm; Ⅳ: 8 cm≤DBH<13 cm; Ⅴ: 13 cm≤DBH<18 cm; Ⅵ: 18 cm≤DBH<23 cm; Ⅶ: 23 cm≤DBH<28 cm; Ⅷ: 28 cm≤DBH<33 cm; Ⅸ: 33 cm≤DBH<38 cm; Ⅹ: DBH≥38 cm.图2 安徽黄山风景区微毛樱桃萌枝种群的存活曲线Fig. 2 Survival curve of sprouting population of Cerasus clarofolia (Schneid.) Yu et Li in Huangshan Mountain Scenic Area of Anhui
2.4 安徽黄山风景区微毛樱桃萌枝种群的存活曲线分析
以x龄级开始时的萌枝标准化存活数的自然对数〔ln(lx)〕为纵坐标(y)、龄级为横坐标(x)绘制安徽黄山风景区微毛樱桃萌枝种群的存活曲线,结果见图2。由图2可见:该种群的存活曲线总体上呈现逐渐下降的趋势。根据该存活曲线拟合获得的指数方程为y=13.77e-0.28x(R2=0.631),获得的幂函数方程为y=12.61x-0.99(R2=0.438),并且指数方程的R2值明显大于幂函数方程,表明该微毛樱桃萌枝种群的存活曲线介于Deevey Ⅱ型和Deevey Ⅲ型之间,但更趋向Deevey Ⅱ型,说明该微毛樱桃萌枝种群总体上处于动态的稳定状态。
2.5 安徽黄山风景区微毛樱桃萌枝种群的生存函数分析
由安徽黄山风景区微毛樱桃萌枝种群的生存函数计算结果(表2)可见:随着龄级的增大,该种群的死亡密度函数和危险率函数呈波动的变化趋势,最高值均出现在Ⅰ级萌枝中,分别为0.262和0.437;Ⅱ级萌枝的死亡密度函数和危险率函数均为0.000;Ⅲ级至Ⅹ级萌枝的死亡密度函数和危险率函数变化总体上趋于平缓,仅伴随小周期波动。
由表2还可见:总体上看,安徽黄山风景区微毛樱桃萌枝种群各龄级萌枝的积累死亡率函数持续升高,而其生存率函数却持续下降。该种群Ⅵ级至Ⅹ级萌枝的积累死亡率函数在0.95以上,生存率函数在0.05以下,说明在生长中、后期该微毛樱桃萌枝种群萌枝的生存率稳定在较低水平,但存在小周期波动,并与其对应的死亡率和损失度的变化趋势相似。
表2 安徽黄山风景区微毛樱桃萌枝种群的生存函数1)
Table 2 Survival functions of sprouting population ofCerasusclarofolia(Schneid.) Yu et Li in Huangshan Mountain Scenic Area of Anhui1)

龄级2)Ageclass2)f(t)λ(t)F(t)S(t)Ⅰ0 2620 4370 7850 215Ⅱ0 0000 0000 7850 215Ⅲ0 0190 0990 8410 159Ⅳ0 0250 2050 9160 084Ⅴ0 0090 1340 9440 056Ⅵ0 0070 1650 9660 034Ⅶ0 0000 0000 9660 034Ⅷ0 0050 1900 9810 019Ⅸ0 0020 1020 9860 014Ⅹ0 0000 1880 9920 008
1)f(t): 死亡密度函数 Mortality density function;λ(t): 危险率函数 Hazard rate function;F(t): 积累死亡率函数Cumulative mortality function;S(t): 生存率函数Survival rate function.
2)根据胸径(DBH)和树高(H)划分龄级 According to diameter at breast height (DBH) and height (H) for dividing age class. Ⅰ: DBH<3 cm, H<130 cm; Ⅱ: DBH<3 cm, H≥130 cm; Ⅲ: 3 cm≤DBH<8 cm; Ⅳ: 8 cm≤DBH<13 cm; Ⅴ: 13 cm≤DBH<18 cm; Ⅵ: 18 cm≤DBH<23 cm; Ⅶ: 23 cm≤DBH<28 cm; Ⅷ: 28 cm≤DBH<33 cm; Ⅸ: 33 cm≤DBH<38 cm; Ⅹ: DBH≥38 cm.
3 讨 论
Cook[21]认为,萌枝是植物替代有性繁殖的一种繁殖方式,并且,萌枝更新较实生更新具有更多优越性,可能是对缓解植物个体高死亡率危机的一种适应措施。幼苗或幼树的萌枝生长速率远远高于实生苗,萌枝可利用基株自身广大的根系以及树木其余部分储存的代谢产物进行高生长以占据较高的生存空间,从而不易被遮光[22]。静态生命表结合生存函数能够很好地反映种群的结构和动态变化[23],而存活曲线则能够直观地反映出种群在发育过程中各龄级个体的死亡状况,从而反映出种群的个体数量动态变化[24]。安徽黄山风景区微毛樱桃萌枝种群萌枝的死亡率(qx)和损失度(Kx)在整个生长发育过程中共出现2个峰值,说明该种群萌枝有2个死亡高峰,其中第1个死亡高峰出现在Ⅰ级萌枝,并在Ⅱ级萌枝迅速降低,这可能主要是因为微毛樱桃为喜光树种,Ⅰ级萌枝主要由1年生和2年生萌枝组成,因此,萌枝生长对光照的依赖性较强,而处于林内的实生幼苗往往受林冠层和灌草层的遮蔽,光合作用受到明显抑制,加上幼树密度高,随着龄级的提高,为了争夺空间和光能,萌枝间发生激烈的竞争,导致仅有少量萌枝能够发育为幼树。对朴树(CeltissinensisPers.)[25]、沉水樟〔Cinnamomummicranthum(Hay.) Hay.〕[26]、大叶榉(ZelkovaschneiderianaHand.-Mazz.)[27]、木荷(SchimasuperbaGardn. et Champ.)[28]和红椿(ToonaciliataRoem.)[29]等树种种群的相关研究结果均表明幼苗和幼树的死亡率较高。当部分Ⅰ级萌枝生长为Ⅱ级萌枝时,由于生长前期萌枝能够借助基株储存的养分快速进行高生长,其生存空间亦较Ⅰ级萌枝有所改善,而且能够较好地摆脱华箬竹及草本植物的干扰,故死亡率明显下降。本研究中,安徽黄山风景区微毛樱桃萌枝种群Ⅵ级至Ⅹ级萌枝的生存率函数均稳定在0.05以下,并处于小周期的波动状态,说明该种群萌枝的生存率稳定在较低水平。该种群萌枝的积累死亡率函数则随龄级增大而不断升高,这可能是因为该种群的少数萌枝(即老龄级萌枝)在进入林冠层以后对光照、营养和空间资源的需求均增大,与上层林木树种发生资源生态位重叠的可能性也增大,且这些萌枝均处于生长后期,各项生理机能开始退化,对风害、雪灾及病虫害等的抵抗能力下降,最终导致萌枝死亡率上升。
作者在调查过程中还发现,在黄山风景区内,微毛樱桃多为萌枝苗,其实生幼苗仅在少数区域呈斑块状分布,并且,实生幼苗的数量十分有限,且分布零散。这可能是内部调节因素和外界环境双重影响的结果。一方面,黄山风景区微毛樱桃萌枝幼苗的分布较集中,导致其在发育过程中进行大量的自疏作用,致使萌枝种群的成活率较低;另一方面,黄山风景区微毛樱桃林下地表的枯枝落叶层较厚,并且华箬竹的根系盘结,对微毛樱桃种子入土萌发和幼苗定居非常不利;再者,微毛樱桃种群所处的山顶区域气候恶劣,风大、湿度高、冬季气温低、雨雪天较多,植株的结实率年际波动较大,且果实受啮齿类动物及鸟类取食明显;此外,林地岩石裸露度高、地表径流、人为干扰等因素也可能是导致微毛樱桃实生幼苗稀少的重要原因。上述生境特点是微毛樱桃种子更新的瓶颈,导致其种群更新主要依赖萌枝方式。萌枝更新对种群的长期存留、打破更新瓶颈及维持群落的稳定性均具有重要意义[1]。由于营养繁殖的风险低于有性繁殖,风险分摊(risk-spreading)和整合作用(integration)可以提高植物萌枝苗对逆境的忍耐力及其与其他物种的竞争力,因而,通常萌枝苗的定居数和存活率远高于实生苗。但是,由于萌枝苗普遍存在林木衰退早、易感染病虫害、应变能力差等方面的严重缺陷和不足,最终导致萌枝种群的遗传多样性降低、生产力下降,对环境变化的反应能力也减弱[16]。
芽库是营养繁殖的潜在来源,植物可通过调控芽库中芽的输入率、输出率和密度应对外界干扰,进而调控植被的组成和动态[30]。在一些极端生境中,植物甚至放弃有性繁殖方式,以芽库作为潜在种群完成更新过程[31]。邓正苗等[31]的研究结果表明:以萌枝为主的植物更新与其自身的芽库有关。作者在实际调查中发现,黄山风景区微毛樱桃的茎干、基部根颈和地下侧根上均含有丰富的芽体,当受到外界干扰后,这些芽体能够迅速萌生大量的幼苗。例如,山体滑坡冲击导致微毛樱桃植株遭到破坏后,其茎干上能够萌发数以百计的幼苗,以延续母体的生活史;再如,生长在岩石裸露或地表径流较为严重的山坡上的微毛樱桃的侧根通常暴露在土壤或岩石外面,这种情况下其植株侧根上的芽体会萌发形成根蘖萌枝。本研究中,微毛樱桃的根蘖萌枝并不常见,且多与根颈萌枝同时出现,而同属植物欧洲甜樱桃的野生植株则以根蘖萌枝为主[2,10],二者萌枝类型有明显区别。
相关研究结果[17,32]表明:植物种群的数量动态与林窗的形成和郁闭度密切相关。安徽黄山风景区微毛樱桃萌枝种群在生长过程中出现2个死亡高峰,极易在局部区域形成林窗,进而扩大更新幼苗的数量、提高种群抵御外界随机干扰的能力。从种群长远发展角度考虑,为了维护黄山风景区微毛樱桃种群的数量,可适当进行小尺度的人为干扰,增加林窗数量以改善其幼苗的生存环境,从而促进种群尤其是数量较少的实生苗种群的更新。综上所述,建议加强对黄山风景区微毛樱桃萌枝种群的抚育管理,并提高种群长期存活的能力,这是该种群可持续发展的必由之路。
[1] BOND W J, MIDGLEY J J. Ecology of sprouting in woody plants: the persistence niche[J]. Trends in Ecology and Evolution, 2001, 16(1): 45-51.
[2] VESK P A, WESTOBY M. Sprouting ability across diverse disturbances and vegetation types worldwide[J]. Journal of Ecology, 2004, 92(2): 310-320.
[3] NISHIMURA S, YONEDA T, FUJII S, et al. Sprouting traits of Fagaceae species in a hill dipterocarp forest, UluGadut, West Sumatra[J]. Journal of Tropical Ecology, 2011, 27(1): 107-110.
[4] 闫恩荣, 王希华, 施家月, 等. 木本植物萌枝生态学研究进展[J]. 应用生态学报, 2005, 16(12): 2359-2464.
[5] 陈 沐, 曹 敏, 林露湘. 木本植物萌生更新研究进展[J]. 生态学杂志, 2007, 26(7): 1114-1118.
[6] 中国科学院中国植物志编辑委员会. 中国植物志: 第三十八卷[M]. 北京: 科学出版社, 1986.
[7] 胡嘉琪, 梁师文. 黄山植物[M]. 上海: 复旦大学出版社, 1996: 2412-2417.
[8] 陈志伟, 伊贤贵, 王贤荣, 等. 黄山微毛樱主要种群生态位特征[J]. 南京林业大学学报(自然科学版), 2014, 38(S1): 39-46.
[9] VAUGHAN S P, COTTRELL J E, MOODLEY D J, et al. Clonal structure and recruitment in British wild cherry (PrunusaviumL.)[J]. Forest Ecology and Management, 2007, 242(2): 419-430.
[10] HÖLTKEN A M, GREGORIUS H-R. Detecting local establishment strategies of wild cherry (PrunusaviumL.)[J]. BMC Ecology, 2006, 6: 13.
[11] DUCCI F, SANTI F. The distribution of clones in managed and unmanaged populations of wild cherry (Prunusavium)[J]. Canadian Journal of Forest Research, 1997, 27(12): 1998-2004.
[12] 丁 晖, 方炎明, 杨新虎, 等. 黄山亚热带常绿阔叶林的群落特征[J]. 生物多样性, 2016, 24(8): 875-887.
[13] 陈志伟. 黄山微毛樱群落数量特征及离体快繁研究[D]. 南京: 南京林业大学森林资源与环境学院, 2011.
[14] 方精云, 王襄平, 沈泽昊, 等. 植物群落清查的主要内容、方法和技术规范[J]. 生物多样性, 2009, 17(6): 533-548.
[15] TREDICI P D. Sprouting in temperate trees: a morphological and ecological review[J]. The Botanical Review, 2001, 67(2): 121-140.
[16] 高贤明, 王 巍, 杜晓军, 等. 北京山区辽东栎林的径级结构、种群起源及生态学意义[J]. 植物生态学报, 2001, 25(6): 673-678.
[17] 刘 峰, 陈伟烈, 贺金生. 神农架地区锐齿槲栎种群结构与更新的研究[J]. 植物生态学报, 2000, 24(4): 396-401.
[18] 陈晓德. 植物种群与群落结构动态量化分析方法研究[J]. 生态学报, 1998, 18(2): 214-217.
[19] 洪 伟, 王新功, 吴承祯, 等. 濒危植物南方红豆杉种群生命表及谱分析[J]. 应用生态学报, 2004, 15(6): 1109-1112.
[20] 杨凤翔, 王顺庆, 徐海根, 等. 生存分析理论及其在研究生命表中的应用[J]. 生态学报, 1991, 11(2): 153-158.
[21] COOK R E. Asexual reproduction: a further consideration[J]. The American Naturalist, 1979, 113(5): 769-772.
[22] BELLINGHAM P J, TANNER E V J, HEALEY J R. Sprouting of trees in Jamaican montane forests, after a hurricane[J]. Journal of Ecology, 1994, 82(4): 747-758.
[23] 吴承祯, 洪 伟, 谢金寿, 等. 珍稀濒危植物长苞铁杉种群生命表分析[J]. 应用生态学报, 2000, 11(3): 333-336.
[24] 李 玲, 张光富, 王 锐, 等. 天目山自然保护区银杏天然种群生命表[J]. 生态学杂志, 2011, 30(1): 53-58.
[25] 闫淑君, 洪 伟, 吴承祯. 闽江口琅岐岛朴树种群结构与空间分布格局[J]. 生态学杂志, 2011, 30(2): 221-226.
[26] 陈远征, 马祥庆, 冯丽贞, 等. 濒危植物沉水樟的种群生命表和谱分析[J]. 生态学报, 2006, 26(12): 4267-4272.
[27] 程红梅. 大蜀山大叶榉种群数量动态研究[J]. 中南林业科技大学学报, 2009, 29(3): 65-69.
[28] 胡喜生, 洪 伟, 吴承祯, 等. 木荷天然种群生命表分析[J]. 广西植物, 2007, 27(3): 469-474.
[29] 汪 洋, 闫魁星, 滕家喜, 等. 鄂西北濒危植物红椿天然种群动态分析[J]. 植物资源与环境学报, 2016, 25(3): 96-102.
[30] DALGLEISH H J, HARTNETT D C. The effects of fire frequency and grazing on tallgrass prairie productivity and plant composition are mediated through bud bank demography[J]. Plant Ecology, 2009, 201(2): 411-420.
[31] 邓正苗, 陈心胜, 谢永宏. 植物芽库的研究进展[J]. 生态学杂志, 2010, 29(9): 1812-1819.
[32] 江明喜, 金义兴, 张全发. 米心水青冈生长过程中的抑制和释放[J]. 应用生态学报, 1995, 6(S1): 153-155.
(责任编辑: 佟金凤)
Study on dynamics of sprouting population ofCerasusclarofoliain Huangshan Mountain Scenic Area of Anhui
YOU Luxiang1a,1b, CHEN Zhiwei2, WANG Huachen1a,1b, YI Xiangui1a,1b, NAN Chenghui3, ZHU Hong1a,1b, WANG Xianrong1a,1b,①
(1. Nanjing Forestry University: a. Co-Innovation Center for Sustainable Forestry in Southern China, b. College of Biology and the Environment, Nanjing 210037, China; 2. Zhangzhou City Forestry Bureau of Fujian Province, Zhangzhou 363000, China; 3. Nanjing Forest Police College, Nanjing 210023, China),J.PlantResour. &Environ., 2017, 26(2): 83-89
In order to explore the growth rule and regeneration characteristics of sprouting population ofCerasusclarofolia(Schneid.) Yu et Li in Huangshan Mountain Scenic Area of Anhui, sprouting branch type, age class structure 〔replaced by diameter class structure, according to diameter at breast height (DBH) and height (H) for dividing diameter class〕, static life table, survival curve and survival function of this population were analyzed respectively on the basis of field survey. The results show that the main sprouting branch in this population is root collar sprouting branch, root sucker sprouting branch is fewer and mostly appears simultaneously with root collar sprouting branch. Number of young sprouting branches is more, that of middle age sprouting branches is less, while that of old age sprouting branches is even less, indicating that there are many seedlings and young trees but a few middle age and old plants in this population. Dynamic index of variation of sprouting branch number of this population is 56.39% in natural state, while that is 5.64% when considering disturbance. On the whole, survival number and standardized survival number of sprouting branches are more from Ⅰ class (DBH<3 cm, H<130 cm) to Ⅲ class (3 cm≤DBH<8 cm), and those are less from Ⅷ class (28 cm≤DBH<33 cm) to Ⅹ class (DBH≥38 cm). Life expectancy of sprouting branch at Ⅱ class (DBH<3 cm, H≥130 cm) is the highest (2.980), while that at Ⅹ class is the lowest (0.500). Variation tendencies of mortality rate and loss degree of sprouting branch at each class are basically consistent with two peak values appearing in sprouting branches at Ⅰ and Ⅸ class, respectively. Survival curve of this population is between Deevey Ⅱ type and Deevey Ⅲ type, but is closer to Deevey Ⅱ type. Mortality density function and hazard rate function of sprouting branch at Ⅰ class are the largest, those at Ⅱ class are 0.000, and those at other age classes tend to flatten. With increasing of age class, cumulative mortality function of sprouting branch elevates continually, while survival rate function decreases continually. It is suggested that in general, sprouting population ofC.clarofoliain Huangshan Mountain Scenic Area is in a dynamic stable state. From the view of population development, appropriate minor artificial disturbance is recommended to improve living environment of seedlings and promote regeneration of the population.
Cerasusclarofolia(Schneid.) Yu et Li; sprouting population; Huangshan Mountain Scenic Area; static life table; survival curve; survival function
2016-10-27
江苏省林业三新工程项目(LYSX[2015]17); 江苏省高校优势学科建设工程资助项目(PAPD)
尤禄祥(1965—),男,江苏南京人,硕士,高级工程师,从事植物资源开发与利用研究。
①通信作者E-mail: wangxianrong66@njfu.edu.cn
Q948.15; Q949.751.8
A
1674-7895(2017)02-0083-07
10.3969/j.issn.1674-7895.2017.02.11
