模拟氮沉降对毛竹非结构性碳的影响*
2017-08-30鲍永新裘子炎吕茹冰宋新章
杜 莹 鲍永新 裘子炎 吕茹冰 宋 超 宋新章,2
(1. 浙江农林大学亚热带森林培育国家重点实验室培育基地 临安 311300; 2.浙江天目山森林生态系统定位观测研究站 临安 311300)
模拟氮沉降对毛竹非结构性碳的影响*
杜 莹1鲍永新1裘子炎1吕茹冰1宋 超1宋新章1,2
(1. 浙江农林大学亚热带森林培育国家重点实验室培育基地 临安 311300; 2.浙江天目山森林生态系统定位观测研究站 临安 311300)
【目的】 分析不同强度氮沉降对毛竹体内非结构性碳水化合物(NSC)含量的影响,为全面认识氮沉降对毛竹林生态系统的影响提供科学依据。【方法】 在临安市毛竹林长期试验基地,采用典型选样法,设置12块20 m×20 m样地,包括低氮(30 kg·hm-2a-1)、中氮(60 kg·hm-2a-1)、高氮(90 kg·hm-2a-1)和对照(0 kg·hm-2a-1)4种处理,每处理设3个重复样地,处理32个月,探讨模拟氮沉降对1龄和3龄毛竹NSC含量的影响。【结果】 可溶性糖和淀粉在NSC中所占比例分别为80%和20%,NSC主要以可溶性糖的形式存在; 竹叶NSC含量最高,分别是枝和秆的1.32和1.27倍; 3龄竹枝和秆NSC含量显著高于1龄竹(P< 0.05),竹龄对竹叶NSC含量影响不显著(P> 0.05); 氮沉降显著降低毛竹叶、3龄竹枝和1龄竹秆的NSC含量(P< 0.05); 氮沉降和竹龄的交互作用对毛竹枝、秆的NSC含量影响显著(P< 0.01),对毛竹叶影响不显著。【结论】 氮沉降显著影响毛竹地上器官的NSC含量及其分配,且随竹龄而显著变化,高强度的氮沉降不利于毛竹NSC的积累和固碳。
氮沉降; 毛竹; 非结构性碳; 可溶性糖; 淀粉
碳水化合物是植物光合作用的主要产物,根据其在植物体内的存在形式,通常分为结构性碳水化合物(structural carbohydrates,SC)和非结构性碳水化合物(non-structural carbohydrates,NSC)2种类型(Luoetal., 2006; 潘庆民等, 2002; 郑云普等, 2014)。SC(如木质素、纤维素和半纤维素等高分子化合物)主要用于支持植物体结构与形态的构建(江志坚等, 2012),NSC主要由淀粉、蔗糖、果糖和葡萄糖等可溶性糖组成,是供应植物生长代谢过程中所需能量的物质(Yeeetal., 2005; Sakamakietal., 2004)。在植物生长发育过程中,植物体内可溶性糖的含量变化较大(Trometal., 1989),且与环境因子密切相关,是植物通过生理调节适应环境变化的重要机制(Hochetal., 2003)。淀粉是植物贮藏器官中最主要的碳水化合物(李东胜等, 2012),主要供应植物将来的能量需求(Yeeetal., 2005; Bansaletal., 2009; Palacioetal., 2007)。作为植物碳水化合物在生产过剩时积累的暂时储存(Kozlowski, 1992),NSC通常是植物呼吸作用的底物,其含量变化直接影响着植物体与外界大气CO2交换的通量(Xuetal., 2008; Maieretal., 2010),反映了植物整体的碳供应状况(Yeeetal., 2005),是评价植物碳收支状况的重要量度指标(李迈和等, 2005; Korner, 2003; 张海燕等, 2013); 同时,NSC也表征了植物生长代谢和存活的能力以及应对外界胁迫干扰的缓冲能力(Wurthetal., 2005; Myersetal., 2007)。研究植物体内NSC及其组分的含量变化,可以了解植物碳水化合物的供应状况(施征等, 2012),对于深入理解生物地球化学碳循环过程、预测和评估未来全球气候变化对森林生态系统碳循环过程的影响均有重要科学意义(郑云普等, 2014)。目前,关于NSC的研究主要集中于NSC在植物体内的分配(印婧婧等, 2009; Kobeetal., 2010; 于丽敏等, 2011)和季节变化(Richardsonetal., 2013; 郑云普等, 2014)及对干旱胁迫的响应(O’Brienetal., 2014; 杜尧等, 2014)等方面,而关于大气氮沉降对植物NSC含量变化影响的研究则少见报道。
近年来,全球范围内的大气氮沉降现象日益加剧(Halletal., 1999; Denteneretal., 2006; Jiaetal., 2016)。多个全球氮沉降预测模型均估计,今后几十年内位于我国中南部的亚热带地区将成为全球大气氮沉降最严重的区域(Gallowayetal., 2008; Liuetal., 2013; Jiaetal., 2014)。事实上,目前我国中东部地区的氮沉降量已高达30 kg·hm-2a-1(Cuietal., 2014; 谢迎新等, 2008),已然成为我国氮沉降最严重的区域。大量的氮沉降将对陆地生态系统生产力和功能产生深远影响, 如增加植物叶片氮含量(Liuetal., 2013)、促进植物光合色素合成从而提高植物的光合能力(贾瑞丰等, 2012)、 增加森林生态系统生物量(Fangetal., 2014; Yuetal., 2014)等。Kobe 等(2010)观测到施氮会增加7种北方阔叶树幼苗根系NSC含量。蒋思思等(2015)发现氮添加会促进油松(Pinustabulaeformis)幼苗叶片中碳水化合物的转移和消耗,促进植物非光合器官的生长以及生物量累积。毛子军等(2010)发现在高CO2浓度和稍高温度下,增加氮素有利于蒙古栎(Quercusmongolica)幼苗NSC的积累。而有关氮沉降对毛竹(Phyllostachysedulis)NSC的影响在国内外均未见报道。
毛竹是我国南方重要的森林资源,面积达387万hm2,约占全国竹林面积的70%,占世界毛竹林面积的80%,且近年来种植面积有不断扩大的趋势(Songetal., 2011; 2015)。毛竹固碳潜力巨大,其乔木层年固碳量为5.097 t·hm-2(周国模等, 2010),是速生阶段杉木(Cunninghamialanceolata)林的1.46倍(方晰等, 2002)、热带山地雨林的6.53~14.99倍(陈德祥等, 2010)、苏南27年生杉木林的2.16倍(阮宏华等, 1997),因此毛竹林在全球碳吸存及减缓气候变化方面的重要作用已受到越来越多的关注。迄今为止,国内外关于氮沉降对毛竹林影响的研究主要集中在凋落物分解(Songetal., 2014; 2015)、化学计量学特征(翁俊等, 2015; Songetal., 2016)、土壤微生物多样性(Lietal., 2016)和土壤酶活性(彭春菊等, 2017)等方面,从NSC角度来探讨氮沉降对毛竹影响的研究尚未见报道。鉴于此,本研究以浙江省临安市青山镇毛竹林长期研究样地为对象,研究不同强度氮沉降对毛竹NSC含量的影响,以期为全面认识氮沉降对毛竹林生态系统的影响提供科学依据。
1 研究区概况
研究区位于浙江省临安市毛竹林长期试验基地(119°42′E,30°14′N)。该区属亚热带季风气候区,年均气温15.6 ℃,年均降水量1 420 mm,≥10 ℃年有效积温2 696.2 ℃,全年无霜期230天,土壤为黄壤土。试验地内毛竹林集约经营已有13年历史,经营措施主要是定期清除林下植被,每年9月施加1次复合肥(N:P2O5:K2O, 15∶6:20),用量为450 kg·hm-2,并结合施肥对土壤深翻30 cm(Songetal., 2015; 顾鸿昊等, 2015)。
2 研究方法
2.1 试验设计
在试验地设置立竹情况和环境状况较一致的20 m × 20 m样地12块,进行喷氮试验。样地之间有20 m宽的缓冲带以防相互影响。依据我国亚热带地区的实际氮沉降量(30~37 kg·hm-2a-1)(Cuietal., 2014; 谢迎新等, 2008)及未来增加趋势(Gallowayetal., 2008; Reayetal., 2008),氮沉降处理设置 4个水平: 对照(CK,0 kg·hm-2a-1)、低氮(N30,30 kg·hm-2a-1)、中氮(N60,60 hm-2a-1)和高氮(N90,90 kg·hm-2a-1),每个施氮水平设3个重复样地。不同水平样地的林分和土壤特征详见表1(顾鸿昊等, 2015)。2013年1月起开始模拟氮沉降喷施,每月1次。具体方法(Songetal., 2015)为: 每月月初将定量的NH4NO3充分溶解在10 L自来水中,用电动背式喷雾器均匀喷洒在每个样地的林地表面(相当于年增加降水0.3 mm)。对照样地喷施10 L不加氮的水。
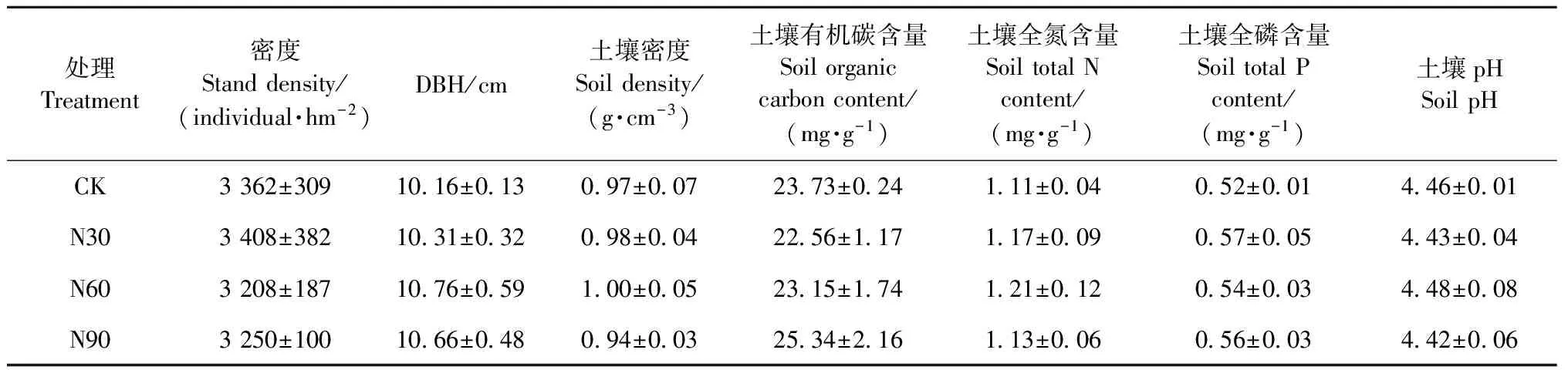
表1 毛竹林分和表层土壤 (0~20 cm) 初始特征
2.2 毛竹各器官采集与测定
试验地内竹林隔年留养新竹并采伐老竹,因此存在明显的大小年现象。竹林采伐经营遵循“存三去四”原则,一般4年生以上老竹于9月采伐,现存立竹分别是2012年(3龄竹)和2014年出土(1龄竹)。在模拟喷氮32个月后, 2015年8月底在各样地内随机选取1龄竹和3龄竹各5株,每株采集位于冠层中上部的枝、叶样品和距地面1和1.7 m处的竹秆样品各50 g,将同一样地内同龄竹的叶、枝、秆样品分别混合后带回实验室在105 ℃下杀青30 min,后在65 ℃下烘干48 h至恒质量,粉碎用于化学分析。本研究将NSC定义为可溶性糖(葡萄糖、蔗糖、果糖等)和淀粉的总和(Hochetal., 2003; 于丽敏等, 2011)。可溶性糖和淀粉含量采用改进的苯酚硫酸法进行测定,具体方法参考于丽敏等(2011)。NSC含量用干质量百分比(%)表示。
2.3 数据分析
运用SPSS16.0和Excel 2010软件进行数据分析。采用单因素方差分析(one-way ANOVA)和多重比较(Duncan)分析不同氮沉降强度下不同年龄段毛竹叶、枝、秆NSC含量的差异及同年龄段不同器官NSC含量差异,以氮沉降强度水平和毛竹年龄作为影响因子进行双因素方差分析。数据满足方差齐性检验要求。采用SigmaPlot12.3软件制图。
3 结果与分析
3.1 可溶性糖含量
由图1可知,3龄竹对照组叶的可溶性糖含量极显著高于处理组(P< 0.01),达4.7%,是氮沉降处理组的1.21~1.37倍,但3个氮沉降处理组间无显著差异(P> 0.05)。类似的,1龄竹对照组叶的可溶性糖含量也极显著高于处理组(P< 0.01),达5.1%,是氮沉降处理组的1.32~1.51倍,高氮处理组最低。3龄竹叶的可溶性糖含量除在高氮处理下显著高于1龄竹叶外(P< 0.05),在其他处理间均未表现出显著差异(P> 0.05)。各处理下枝的可溶性糖含量均低于叶片。低氮处理显著降低了3龄竹枝的可溶性糖含量(P< 0.05),中氮处理显著增加了1龄竹枝的可溶性糖含量(P< 0.05),其余处理间则未表现出显著差异(P> 0.05)。3龄竹秆的可溶性糖含量随氮沉降量的增加呈现先降低后增加的趋势,在中氮处理下最低。中、低强度的氮处理对1龄竹秆的可溶性糖含量无显著影响(P> 0.05),但高氮处理显著降低了其含量(P< 0.05)。低氮和高氮处理下1龄竹秆的可溶性糖含量均显著低于3龄竹(P< 0.05)。
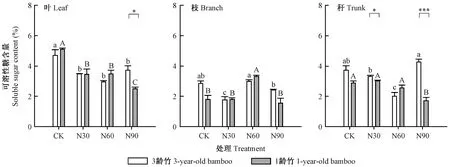
图1 不同氮沉降处理下毛竹叶、枝、秆的可溶性糖含量Fig.1 Soluble sugar content in leaf,branch and trunk of Phyllostachys edulis under different nitrogen addition treatments大写字母表示1龄竹,小写字母表示3龄竹,不同字母表示不同施氮水平间可溶性糖含量差异显著(P< 0.05)。星号表示相同氮处理下不同年龄毛竹各器官差异显著。*: P< 0.05; **: P< 0.01; ***: P< 0.001。下同。Capital letters indicated 1-year-old bamboo, small letters indicated 3-year-old bamboo, different letters indicated significant differences of soluble sugar content among different nitrogen addition rates at 0.05 level.Asterisks indicate significant differences of soluble sugar content of leaf,branch and trunk at different growth stage under the same nitrogen addition treatment. *: P < 0.05; **: P < 0.01; ***: P< 0.001.The same below.
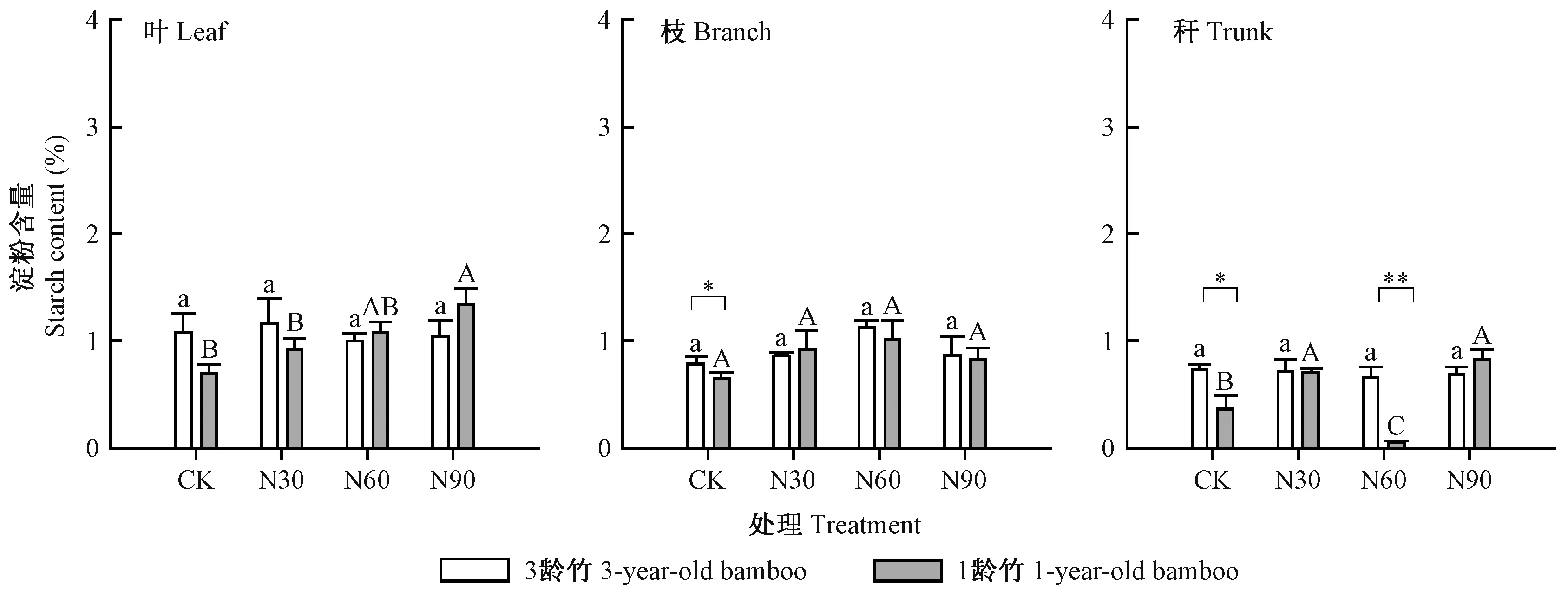
图2 不同氮沉降处理下毛竹叶、枝、秆的淀粉含量Fig.2 Starch content in leaf,branch and trunk of Phyllostachys edulis under different nitrogen addition treatments
3.2 淀粉含量
由图2可知,3龄竹叶的淀粉含量在各处理间无显著差异(P> 0.05),各处理含量为1.0%~1.2%。1龄竹叶的淀粉含量在高氮处理下最高,为1.4%,其余处理间无显著差异(P> 0.05)。3龄竹和1龄竹枝的淀粉含量在各处理间均无显著差异(P> 0.05),3龄竹枝的淀粉含量在对照处理下显著高于1龄竹枝(P< 0.05),但在施氮后二者未表现出显著差异(P> 0.05)。1龄竹秆的淀粉含量在高氮处理下最高,为0.8%,在中氮处理下最低。3龄竹秆的淀粉含量在各处理间无显著差异(P> 0.05),在对照和中氮处理下显著高于1龄竹秆(P< 0.05)。对比图1可知,1龄和3龄竹各器官的淀粉含量均显著低于可溶性糖含量(P< 0.05)。
3.3 NSC含量
由图3可知,对照组3龄竹叶的NSC含量显著高于处理组(P< 0.05),达5.8%,是氮沉降处理组的 1.19~1.31倍,但处理组间无显著差异(P> 0.05)。1龄竹叶的对照组NSC含量显著高于处理组(P< 0.01),达5.8%,是氮沉降处理组的1.21~1.25倍,处理组间无显著差异(P< 0.05)。枝的NSC含量同样低于叶。3龄竹枝的NSC含量在中氮处理下最高,为4.12%,是其他处理组的1.12~1.37倍,在低氮处理下最低。1龄竹枝的NSC含量同样在中氮处理下最高,为4.50%,是其他处理组的1.38~1.48倍,其余处理组间无显著差异(P> 0.05)。3龄竹枝NSC含量在对照处理下显著高于1龄竹枝(P< 0.05),而在中氮处理下显著低于1龄竹枝(P< 0.01)。中高强度的氮处理显著降低了1龄竹秆的NSC含量(P< 0.01)。3龄竹秆的NSC含量随氮沉降量的增加呈现先降低后增加的趋势,在对照和高氮处理下显著高于1龄竹秆(P< 0.05)。
3.4 氮沉降和竹龄对NSC含量的交互作用
方差分析结果(表2)表明,氮沉降水平对毛竹叶、枝、秆NSC含量均有显著影响(P< 0.001); 毛竹年龄显著影响枝NSC含量(P< 0.05),极显著影响秆的NSC含量(P< 0.001),而对叶NSC含量无显著影响(P> 0.05)。氮沉降与竹龄的交互作用显著影响毛竹枝和秆的NSC含量(P< 0.01)。
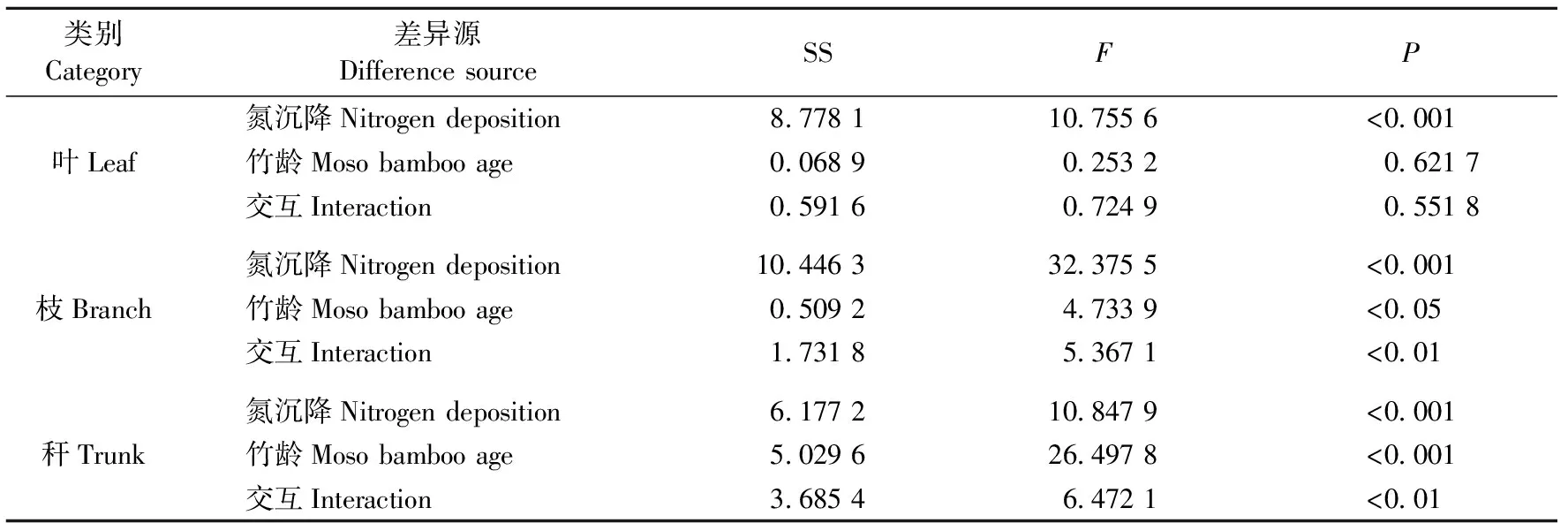
表2 氮沉降和竹龄对毛竹叶、枝、秆NSC含量影响的双因素方差分析
4 讨论
4.1 NSC的组成与分配
本研究中,竹叶可溶性糖含量占NSC的74.6%~83.9%,远高于淀粉(16.1%~25.4%),枝和秆也表现出同样趋势,这表明毛竹叶、枝和秆中的NSC主要以可溶性糖的形式存在。可溶性糖是植物中活跃的碳储存物质,其分配格局反映了植物生长状况(于丽敏等, 2011); 而淀粉是植物能量主要的长期贮存形式,主要供应植物将来的能量需求(Yeeetal., 2005; Bansaletal., 2009; Palacioetal., 2007)。植物NSC组成、分布、贮存和生理功能与植物器官有较大关系(Hochetal., 2003)。毛竹叶NSC含量(4.8%)显著高于枝(3.2%)和秆(3.5%)(P< 0.05)。叶、枝和秆分别占毛竹地上器官生物量的9.2%,15.6%和75.2%(王晨等, 2014),因此NSC在毛竹地上部分各器官中的储量表现为秆>枝>叶。印婧婧等(2009)和吴杰等(2010)也发现叶片是NSC的合成器官,树干是NSC的主要贮存器官。采样时间8月底处于毛竹生长旺季末期(施建敏等, 2007),毛竹在维持叶片NSC含量的同时,将合成的大部分NSC转运到秆中进行体内物质的积累和结构性生长(周国模等, 2010)。
4.2 氮沉降对NSC的影响
本研究发现,竹叶NSC含量在对照处理下最高,秆NSC含量在对照和低氮处理下较高,枝NSC含量在中氮处理下最高,表明毛竹各器官对氮添加有不同的适应性,毛竹能通过改变NSC在各器官的分配来适应氮的增加。叶是NSC的合成器官,秆是毛竹碳储量的主要贡献器官,其生物量和碳储量均占毛竹地上部分的75%以上(漆良华等, 2009; 周国模等, 2010; 王晨等, 2014),毛竹NSC的积累主要取决于叶和秆。由此可以预测,高强度的氮沉降(> 60 kg·hm-2a-1)不利于毛竹NSC的积累,进而影响其生长和固碳能力。
4.3 年龄对NSC组成与分配的影响
3龄竹叶、枝和秆的可溶性糖含量分别比1龄竹增加了2.4%、8.4%和1.2%。因为可溶性糖含量与植物的抗逆性密切相关(刘香萍等, 2010),因此这可能意味着3龄竹的抗逆性要高于1龄竹。
本研究采集的3龄竹和1龄竹的竹叶均是2015年春季长出的新叶,其年龄是一样的,这可能是竹龄对叶NSC含量影响不显著(表2)的一个重要原因。对照处理下,3龄竹枝和秆的NSC含量(3.4%和3.9%)均显著高于1龄竹(3.1%和3.0%)(图3),这可能是因为1龄竹仍处在干物质的快速积累阶段(周国模等, 2010),光合产物被更多地用于结构性生长,而3龄竹已基本成熟,光合产物被更多地以NSC的形式在枝和秆中储存起来。施氮后,多数氮处理下这种竹龄之间的显著差异消失了。
4.4 氮沉降和年龄对NSC的交互作用
尽管氮沉降显著影响竹叶的NSC含量(P< 0.001),但竹龄对叶的NSC含量无显著影响,二者的交互作用也没有显著影响竹叶的NSC含量,这表明竹龄和氮沉降对竹叶NSC含量没有显著的交互作用(P> 0.05)。氮沉降、竹龄及其交互作用均显著影响枝和秆的NSC含量。秆是毛竹生物量和碳储量的主要器官(漆良华等, 2009; 周国模等, 2010; 王晨等, 2014),该结果意味着氮沉降对毛竹地上部分NSC含量及其分配的影响将随着竹龄的增长而显著变化。
5 结论
3龄竹的NSC含量高于1龄竹,但施氮处理后,竹龄对毛竹NSC产生的影响明显减弱。适当的氮沉降能明显提高毛竹的NSC含量,然而超过一定阈值后,毛竹各器官中NSC分配比例将发生变化,不利于毛竹NSC积累;尤其当氮沉降大于60 kg·hm-2a-1时,毛竹叶和秆的NSC含量明显下降,不利于毛竹生长和固碳。
陈德祥, 李意德, Heping Liu, 等. 2010. 尖峰岭热带山地雨林生物量及碳库动态. 中国科学: 生命科学, 40(7): 596-609.
(Chen D X, Li Y D, Liu H,etal. 2010. Biomass and carbon dynamics of a tropical mountain rain forest in China. Scientia Sinica Vitae, 40(7): 596-609. [in Chinese])
杜 尧, 韩 轶, 王传宽. 2014. 干旱对兴安落叶松枝叶非结构性碳水化合物的影响. 生态学报, 34(21): 6090-6100.
(Du Y, Han Y, Wang C K. 2014. The influence of drought on non-structural carbohydrates in the needles and twigs ofLarixgmelinii. Acta Ecologica Sinica, 34(21): 6090-6100. [in Chinese])
方 晰, 田大伦, 项文化, 等. 2002. 第二代杉木中幼林生态系统碳动态与平衡. 中南林学院学报, 22(1): 1-6.
(Fa X, Tian D L, Xiang W H,etal. 2002. Carbon dynamics and balance in the ecosystem of the young and middle-aged second-generation Chinese fir plantation. Journal of Central South Forestry University, 22(1): 1-6. [in Chinese])
顾鸿昊, 翁 俊, 孔佳杰, 等. 2015. 粗放和集约经营毛竹林叶片的生态化学计量特征. 浙江农林大学学报, 32(5): 661-667.
(Gu H H, Weng J, Kong J J,etal. 2015. Ecological stoichiometry ofPhyllostachysedulisleaves with extensive and intensive management. Journal of Zhejiang A&F University. 32(5): 661-667.[in Chinese]).
贾瑞丰, 尹光天, 杨锦昌, 等. 2012. 不同氮素水平对红厚壳幼苗生长及光合特性的影响. 林业科学研究, 25(1): 23-29.
(Jia R F, Yin G T, Yang J C,etal. 2012. Effects of nitrogen application on the growth and photosynthetic characteristics ofCalophylluminophyllumseedlings. Forest Research, 25(1): 23-29. [in Chinese])
蒋思思, 魏丽萍, 杨 松, 等. 2015. 不同种源油松幼苗的光合色素和非结构性碳水化合物对模拟氮沉降的短期响应. 生态学报, 35(21): 7061-7070.
(Jiang S S, Wei L P, Yang S,etal. 2015. Short term responses of photosynthetic pigments and nonstructural carbohydrates to simulated nitrogen deposition in three provenances ofPinustabulaeformisCarr. seedlings. Acta Ecologica Sinica, 35(21): 7061-7070. [in Chinese])
江志坚, 黄小平, 张景平. 2012. 环境胁迫对海草非结构性碳水化合物储存和转移的影响. 生态学报, 32(19): 6242-6250.
(Jiang Z J, Huang X P, Zhang J P. 2012. Effect of environmental stress on non-structural carbohydrates reserves and transfer in seagrasses. Acta Ecologica Sinica, 32(19): 6242-6250. [in Chinese])
李东胜, 史作民, 刘世荣, 等. 2012. 南北样带温带区栎属树种种子化学组成与气候因子的关系. 生态学报, 32(24): 7857-7865.
(Li D S, Shi Z M, Liu S R,etal. 2012. Relationships between chemical compositions ofQuercusspecies seeds and climatic factors in temperate zone of NSTEC. Acta Ecologica Sinica, 32(24): 7857-7865. [in Chinese])
李迈和, Norbert K. 2005. 全球高山林线研究现状与发展方向. 四川林业科技, 26(4): 36-42.
(Li M H, Norbert K. 2005. The state of knowledge on alpine treeline and suggestions for future research. Journal of Sichuan Forestry Science and Technology, 26(4): 36-42. [in Chinese])
刘香萍, 崔国文, 李国良, 等. 2010. 紫花苜蓿主根内非结构性碳水化合物累积及其与抗寒性的关系. 中国草地学报, 32(2): 113-115.
(Liu X P, Cui G W, Li G L,etal. 2010. Relationships between non-structure carbohydrates accumulation in taproot of alfalfa and coldhardiness. Chinese Journal of Grassland, 32(2): 113-115. [in Chinese])
毛子军, 贾桂梅, 刘林馨, 等. 2010. 温度增高、CO2浓度升高、施氮对蒙古栎幼苗非结构碳水化合物积累及其分配的综合影响. 植物生态学报, 34(10): 1174-1184.
(Mao Z J, Jia G M, Liu L X,etal. 2010. Combined effects of elevated temperature, elevated [CO2] and nitrogen supply on non-structural carbohydrate accumulation and allocation inQuercusmongolicaseedlings. Chinese Journal of Plant Ecology, 34(10): 1174-1184. [in Chinese])
潘庆民, 韩兴国, 白永飞, 等. 2002. 植物非结构性贮藏碳水化合物的生理生态学研究进展. 植物学通报, 19(1): 30-38.
(Pan Q M, Han X G, Bai Y F,etal. 2002. Advances in physiology and ecology studies on stored non-structure carbohydrates in plants. Chinese Bulletin of Botany, 19(1): 30-38. [in Chinese])
彭春菊, 李 全,顾鸿昊,等.2017. 模拟氮沉降及经营方式对毛竹林土壤酶活性的影响. 应用生态学报, 28(2): 423-429.
(Peng C J, Li Q, Gu H H,etal. 2017. Effects of simulated nitrogen deposition and management type on soil enzyme activities in moso bamboo forest. Chinese Journal of Applied Ecology, 28(2): 423-429.[in Chinese])
漆良华, 刘广路, 范少辉, 等. 2009. 不同抚育措施对闽西毛竹林碳密度、碳贮量与碳格局的影响. 生态学杂志, 28(8): 1482-1488.
(Qi L H, Liu G L, Fan S H,etal. 2009. Effects of different tending measures on carbon density, storage, and allocation pattern ofPhyllostachyedulisforests in western Fujian Province. Chinese Journal of Ecology, 28(8): 1482-1488. [in Chinese])
阮宏华, 姜志林, 高苏铭. 1997. 苏南丘陵主要森林类型碳循环研究: 含量与分布规律. 生态学杂志, 16(6): 17-21.
(Ruan H H, Jiang Z L, Gao S M. 1997. Preliminary studies of carbon cycling in three types of forests in the hilly regions of southern Jiangsu Province. Chinese Journal of Ecology, 16(6): 17-21.[in Chinese])
施建敏, 郭起荣, 杨光耀, 等. 2007. 毛竹光合作用对环境因子的季节响应. 广西植物, 27(6): 923-928.
(Shi J M, Guo Q R, Yang G Y,etal. 2007. Seasonal photosynthetic responses ofPhyllostachysedulisto environmental factors. Guihaia, 27 (6): 923-928. [in Chinese])
施 征, 白登忠, 雷静品, 等. 2012. 祁连圆柏光合色素与非结构性碳水化合物含量对海拔变化的响应. 西北植物学报, 32(11): 2286-2292.
(Shi Z, Bai D Z, Lei J P,etal. 2012. Variations of chloroplast pigments and non-structural carbohydrates of sabinaprzewalskiialong altitude in Qilian Mountains timberline. Acta Botanica Boreali-Occidentalia Sinica, 32(11): 2286-2292. [in Chinese])
王 晨, 江泽慧, 郭起荣, 等. 2014. 毛竹地上器官的生物量分配及其随个体大小变化的规律. 生态学杂志, 33(8): 2019-2024.
(Wang C, Jiang Z H, Guo Q R,etal. 2014. Biomass allocation of aboveground components ofPhyllostachysedulisand its variation with body size. Chinese Journal of Ecology, 33(8): 2019-2024. [in Chinese])
翁 俊, 顾鸿昊, 王志坤, 等. 2015. 氮沉降对毛竹叶片生态化学计量特征的影响. 生态科学, 34(2): 63-70.
(Weng J, Gu H H, Wang Z K,etal. 2015. The effects of nitrogen deposition on ecological stoichiometry of leaf of moso bamboo with extensive management. Ecological Science, 34(2): 63-70. [in Chinese])
吴 杰, 潘红丽, 杜 忠, 等. 2010. 卧龙竹类非结构性碳水化合物与叶氮含量对海拔的响应. 生态学报, 30(3): 0610-0618.
(Wu J, Pan H L, Du Z,etal. 2010. Responses of tissue non-structural carbohydrates and leaf nitrogen contents to altitude in two dwarf bamboos in Wolong. Acta Ecologica Sinica, 30(3): 0610-0618. [in Chinese])
于丽敏, 王传宽, 王兴昌. 2011. 三种温带树种非结构性碳水化合物的分配. 植物生态学报, 35 (12): 1245-1255.
(Yu L M, Wang C K, Wang X C. 2011. Allocation of nonstructural carbohydrates for three temperate tree species in Northeast China. Chinese Journal of Plant Ecology, 35 (12): 1245-1255.[in Chinese])
印婧婧, 郭大立, 何思源, 等. 2009. 内蒙古半干旱区树木非结构性碳、氮、磷的分配格局. 北京大学学报:自然科学版, 45(3): 519-527.
(Yin J J, Guo D L, He S Y,etal. 2009. Non-structural carbohydrate, N, and P allocation patterns of two temperate tree species in a semi-arid region of Inner Mongolia. Acta Scientiarum Naturalium Universitatis Pekinensis, 45 (3): 519-527. [in Chinese])
张海燕, 王传宽, 王兴昌, 等. 2013. 白桦和紫椴树干非结构性碳水化合物的空间变异. 应用生态学报. 24(11): 3050-3056.
(Zhang H Y, Wang C K, Wang X C,etal. 2013. Spatial variation of non-structural carbohydrates inBetulaplatyphyllaandTiliaamurensisstems. Chinese Journal of Applied Ecology, 24(11): 3050-3056. [in Chinese])
郑云普, 王贺新, 娄 鑫, 等. 2014. 木本植物非结构性碳水化合物变化及其影响因子研究进展. 应用生态学报, 25(4): 1188-1196.
(Zheng Y P, Wang H X, Lou X,etal. 2014. Changes of non-structural carbohydrates and its impact factors in trees. Chinese Journal of Applied Ecology, 25(4): 1188-1196. [in Chinese])
周国模, 姜培坤, 徐秋芳. 2010. 竹林生态系统中的碳的固定与转化. 北京: 科学出版社.
(Zhou G M, Jiang P K, Xu Q F. 2010.Carbon fixing and transition in the ecosystem of bamboo forest. Beijing: Science Press.[in Chinese])
Bansal S, Germino M J. 2009. Temporal variation of non-structural carbohydrates in montane conifers: similarities and differences among developmental stages, species and environmental conditions. Tree Physiology, 29(4): 559-568.
Cui J, Zhou J, Peng Y,etal. 2014. Atmospheric wet deposition of nitrogen and sulfur in the agroecosystem in developing and developed areas of Southeastern China. Atmospheric Environment, 89(2): 102-108.
Dentener F, Drevet J, Lamarque J F,etal. 2006. Nitrogen and sulfur deposition on regional and global scales: a multimodel evaluation. Global Biogeochemical Cycles, 20(4): 16615-16615.
Fang J, Kato T, Guo Z,etal. 2014. Evidence for environmentally enhanced forest growth. Proceedings of the National Academy of Sciences of the United States of America, 111(26): 9527-9532.
Galloway J N, Townsend A R, Erisman J W,etal. 2008. Transformation of the nitrogen cycle: recent trends, questions, and potential solutions. Science, 320(5878): 889-892.
Hall S J, Matson P A. 1999. Nitrogen oxide emissions after nitrogen additions in tropical forests. Nature International Weekly Journal of Science, 400(6740): 152-155.
Hoch G, Richer A, Korner C. 2003.Non-structural carbon compounds in temperate forest trees. Plant Cell and Environment, 26(7): 1067-1081.
Jia Y, Yu G, He N,etal. 2014. Spatial and decadal variations in inorganic nitrogen wet deposition in China induced by human activity. Scientific Reports, 4: 3763.
Jia Y, Yu G, Gao Y,etal. 2016. Global inorganic nitrogen dry deposition inferred from ground- and space-based measurements. Scientific Reports, 6: 19810.
Kobe R K, Iyer M, Walters M B. 2010. Optimal partitioning theory revisited: nonstructural carbohydrates dominate root mass responses to nitrogen. Ecology, 91(1): 166-179.
Korner C. 2003. Carbon limitation in trees. Journal of Ecology, 91(1): 4-17.
Kozlowski T T. 1992. Carbohydrate sources and sinks in woody plants. The Botanical Review, 58(2): 107-222.
Li Q, Song X Z, Gu H,etal. 2016. Nitrogen deposition and management practices increase soil microbial biomass carbon but decrease diversity in Moso bamboo plantations. Scientific Reports, 6: 28235.
Liu X J, Zhang Y, Han W X,etal. 2013. Enhanced nitrogen deposition over China. Nature, 494(7438): 459-462.
Luo Z B, Calfapietra C, Liberloo M,etal. 2006. Carbon partitioning to mobile and structural fractions in poplar wood under elevated CO2(EUROFACE) and N fertilization. Global Change Biology, 12(2): 272-283.
Maier C A, Johnsen K H, Clinton B D,etal. 2010. Relationships between stem CO2efflux, substrate supply, and growth in young loblolly pine trees. New Phytologist, 185(2): 502-513.
Myers J A, Kitajima K. 2007. Carbohydrate storage enhances seedling shade and stress tolerance in a neotropical forest. Journal of Ecology, 95(2): 383-395.
O’Brien M J, Leuzinger S, Philipson C D,etal. 2014. Drought survival of tropical tree seedlings enhanced by non-structural carbohydrate levels. Nature Climate Change, 4(8): 710-714.
Palacio S, Millard P, Maestro M,etal. 2007. Non-structural carbohydrates and nitrogen dynamics in mediterranean sub-shrubs: an analysis of the functional role of overwintering leaves. Plant Biology, 9(1): 49-58.
Reay D S, Dentener F, Smith P,etal. 2008. Global nitrogen deposition and carbon sinks. Nature Geoscience, 1(7): 430-437.
Richardson A D, Carbone M S, Keenan T F,etal. 2013. Seasonal dynamics and age of stemwood nonstructural carbohydrates in temperate forest trees. New Phytologist, 197(3): 850-861.
Sakamaki Y, Ino Y. 2004. Response ofnon-structural carbohydrate content of belowground parts in Equisetum arvense according to the irradiance change during a growing season. Journal of Plant Research, 117(5): 385-391.
Song X Z, Gu H H, Wang M,etal. 2016. Management practices regulate the response of moso bamboo foliar stoichiometry to nitrogen deposition. Scientific Reports, 6: 24107.
Song X Z, Jiang H, Zhang Z,etal. 2014. Interactive effects of elevated UV-B radiation and N deposition on moso bamboo litter decomposition. Soil Biology & Biochemistry, 69(1): 11-16.
Song X Z, Zhou G M, Gu H H,etal. 2015. Management practices amplify the effects of N deposition on leaf litter decomposition of the moso bamboo forest. Plant and Soil, 395(1): 391-400.
Song X Z, Zhou G M, Jiang H,etal. 2011. Carbon sequestration by Chinese bamboo forests, and their ecological benefits: assessment of potential, problems, and future challenges. Environmental Reviews, 19(1): 418-428.
Trom E R, Sheath G W, Bryant A M. 1989. Seasonal variations in total nonstructural carbohydrate and major element levels in perennial ryegrass and paspalum in mixed pasture. New Zealand Journal of Agricultural Research, 32(2): 157-165.
Wurth M K, PelezRiedl S, Wright S J,etal. 2005. Nonstructural carbohydrate pools in a tropical forest. Oecologia, 143(1): 11-24.
Xu X, Kuzyakov Y, Wanek W,etal. 2008. Root-derived respiration and non-structural carbon of rice seedlings. European Journal of Soil Biology, 44(1): 22-29.
Yee D A, Tissue D T. 2005. Relationships betweennon-structural carbohydrate concentration and flowering in a subtropical herb,Heliconiacaribaea(Heliconiaceae). Caribbean Journal Ofence, 41(2): 243-249.
Yu G, Chen Z, Piao S,etal. 2014. High carbon dioxide uptake by subtropical forest ecosystems in the East Asian monsoon region. Proceedings of the National Academy of Sciences of the United States of America, 111(13): 4910-4915.
(责任编辑 于静娴)
Effects of Simulated Nitrogen Deposition on Non-Structural Carbohydrates of Moso Bamboo
Du Ying1Bao Yongxin1Qiu Ziyan1Lü Rubing1Song Chao1Song Xinzhang1,2
(1.StateKeyLaboratoryofSubtropicalSilviculture,ZhejiangA&FUniversityLin’an311300; 2.TianmuMountainForestEcosystemResearchStation,ZhejiangProvinceLin’an311300)
【Objective】 The subtropical zone in south-central China has become the region with the greatest nitrogen (N) deposition in the world. Most moso bamboo (Phyllostachysedulis) plantations of China are distributed in the center of this region. The change of non-structural carbohydrate (NSC) in plant can reflect the effects of environmental factors on plant. However, the effects of N deposition on NSC content of moso bamboo remain unclear. The aim of this study is to estimate the effects of N deposition on the NSC contents of leaf, branch and trunk of moso bamboo at different growth stages.【Method】 The experiment was conducted in moso bamboo plantations over a 32-month period and included 4 N content levels(30, 60, 90 kg·hm-2a-1and zero as control)were applied in the study.Each N-content was replicated for 3 times.【Result】 The soluble sugar and starch accounted for 80% and 20% of NSC, respectively. Bamboo leaf has the highest NSC content, 1.32 and 1.27 times higher than branch and trunk, respectively. The NSC content of branch and trunk of 3-year-old bamboo were significantly higher than that of 1-year-old bamboo (P< 0.05). No significant effect of moso bamboo age on the NSC content of leaf was observed (P> 0.05). The NSC content of leaf and branch of 3-year-old bamboo, and trunk of 1-year-old bamboo (P< 0.05) was significantly decreased by N deposition. The interaction between N deposition and moso bamboo ages significantly influenced the NSC content of branch and trunk (P< 0.01) but not leaf were.【Conclusion】 The content and distribution of NSC in moso bamboo were significantly impacted by N deposition, and these effects varied significantly with the bamboo ages. Our result indicated that the excessive atmospheric N deposition (≥ 90 kg·hm-2a-1) in the future would restrain the NSC accumulation in moso bamboo and thus its carbon sequestration capacity.
nitrogen deposition; moso bamboo; non-structural carbohydrates; soluble sugar; starch
10.11707/j.1001-7488.20170702
2015-11-28;
2017-06-09。
国家林业局“948”项目(2013-4-55);国家自然科学基金项目(31470529, 31270517)。
S718.43
A
1001-7488(2017)07-0010-08
*宋新章为通讯作者。
