缺氧和充氧栽培对水稻Cd积累及OsHMA2基因表达的影响
2017-08-07杨耀帅梅秀芹李取生周雪芳陈柯罕
杨耀帅,梅秀芹,李取生,魏 佳,周雪芳,陈柯罕,周 婷
(暨南大学 环境学院/广州市环境暴露与健康重点实验室/广东省环境污染控制与修复材料中心,广东 广州 510632)
缺氧和充氧栽培对水稻Cd积累及OsHMA2基因表达的影响
杨耀帅,梅秀芹,李取生,魏 佳,周雪芳,陈柯罕,周 婷
(暨南大学 环境学院/广州市环境暴露与健康重点实验室/广东省环境污染控制与修复材料中心,广东 广州 510632)
【目的】研究缺氧和充氧栽培对水稻重金属Cd的吸收和转运及OsHMA2基因在茎中表达的影响,为降低水稻中重金属的吸收和累积奠定基础。【方法】以水稻品种五丰优2168为试验材料,通过营养液缺氧和营养液充氧的水培盆栽试验,分析Cd质量浓度为0、0.6、1.2 mg·L–1时,Cd的吸收累积以及茎中OsHMA2基因表达水平。【结果】缺氧栽培条件下,水稻生长发育受Cd抑制作用不显著,而充氧栽培条件下水稻生长受到显著抑制,表现为根、地上部干质量显著下降。在0.6和1.2 mg·L–1Cd处理条件下,缺氧栽培的水稻根部和地上部Cd累积量均低于充氧栽培。2种栽培条件下,根部Cd累积量均随Cd处理浓度的增大而增大。水稻地上部对Cd的累积在缺氧栽培条件下,0.6和1.2 mg·L–1Cd处理无显著差异,充氧栽培的水稻地上部Cd累积随处理浓度增大而增大。与对照相比,水稻茎OsHMA2表达量在缺氧和充氧栽培下,0.6 mg·L–1Cd处理上调,而1.2 mg·L–1Cd处理下调;在Cd处理下,OsHMA2表达量缺氧栽培高于充氧栽培。【结论】缺氧栽培能抑制水稻对Cd的吸收和累积;Cd累积量达到一定值时能够下调OsHMA2的表达。
水稻;缺氧;充氧;镉;OsHMA2;转运系数
镉(Cd)是植物和人体的非必须元素,食物的摄入是Cd进入人体的主要途径[1-2]。Cd对人体的危害极高,主要表现为在人体的肝、肾、胰腺、甲状腺和骨骼中累积并引发各种疾病,20世纪发生在日本的骨痛病就是Cd在人体内大量累积而引起的。Cd能抑制植物的生长发育,在植物体内累积并通过食物链进入人体[3-4]。水稻Oryza sativa作为主要的粮食作物,也是Cd进入人体的主要途径[5]。水稻在淹水厌氧的条件下,根系会释放O2促使根表铁膜的形成,降低或促进水稻对重金属和有毒元素的吸收[6-7]。纪雄辉等[8]研究水稻在长期淹水、间歇灌溉和湿润灌溉的条件下对Cd的吸收,结果表明随着淹水程度的提高,水稻根表铁膜吸附的Fe2+、Mn2+显著增加,根表铁膜变厚,对Cd的吸收程度是湿润灌溉>间歇灌溉>长期淹水。研究[9-10]表明,在淹水条件下植物根际土壤与大气的交换受阻,土壤中的微生物大量消耗氧分,导致还原性物质增多,质子被消耗,pH升高,导致土壤对Cd的吸附增强,降低了Cd的生物有效性,Cd在氧化条件下的生物有效性比还原条件的强,更容易由无效态转化为水溶态和交换态。Reddy等[11]认为,随着水稻根系的氧分增加,氧化还原电位增大,土壤的生物有效态Cd、植物吸收的Cd和地上部分的Cd含量也随之升高。
OsHMA2是水稻中负责运输Zn和Cd的转运蛋白,在Zn和Cd向木质部的装载中发挥作用,并且参与这些金属从地下部向地上部的转运[12-13],而重金属Cd要通过某些转运基因将其装载至木质部才能到达谷粒中[14]。
本研究通过营养液缺氧和营养液充氧栽培试验模拟厌氧和好氧的土壤环境,在2个不同Cd浓度条件下,研究水稻的生长发育,水稻中Cd、Zn的累积变化以及茎中OsHMA2基因的表达水平,了解不同氧分管理下,水稻对Cd的吸收和转运及OsHMA2基因的表达水平,为研究重金属转运基因如何降低稻谷中重金属的累积奠定基础。
1 材料与方法
1.1 材料
供试水稻品种选用广东省金稻种业有限公司的五丰优2168。选取健壮饱满的种子,用w为30%的过氧化氢浸泡15 min消毒。充分漂洗后,在35 ℃条件下浸种24 h,待种子破胸露白后,将种子转移至清洗干净的石英砂中,待种子长出绿芽后,转移至光照培养箱中培养,培养温度为白天30 ℃,晚上20 ℃,每天施用1/4 Hoagland营养液,营养液含0.11 mg·L–1的ZnSO4·7H2O。
1.2 方法
1.2.1 试验设计 试验在暨南大学环境学院温室内进行。水培试验参考文献[15]的方法,分为营养液缺氧和营养液充氧2个栽培条件。营养液缺氧栽培条件为1 g·L–1的琼脂营养液通N224 排氧,营养充氧栽培为栽培水稻时连续充氧。营养液为1/2 Hoagland营养液。待幼苗培养至3叶期,选择整齐一致的幼苗,移至高21 cm、直径15 cm的塑料桶中,每盆2株,加入Hoagland完全营养液,待植物生长稳定时,分别加入缺氧营养液和有氧营养液, 并加入Cd(NO3)2。每隔5 d换1次营养液。设置2个Cd的质量浓度:0.6、1.2 mg·L–1并设置空白对照(CK)。每个处理4个平行。处理30 d后收获水稻。
1.2.2 样品处理 收获时,将水稻根部用自来水冲洗,再用去离子水洗3次。测定每株水稻的生物量包括根质量、地上部质量和地上部高度。用已消毒灭菌的剪刀剪下根尖2~3 cm和1~2 cm茎节,分别用锡箔纸包好,放入液氮中速冻,再转移至–80 ℃冰箱保存,采用Trizol法提取RNA,测定RNA纯度及逆转录,使用ViiA7 software进行定量。
剩余的根和地上部在105 ℃杀青后70 ℃烘干至恒质量,称质量,然后磨碎,加入8 mL HNO3、180 ℃条件下在微波消解仪中消解,冷却后定容至25 mL,定性滤纸过滤后,用原子吸收分光光度计(PE 900T)测定Cd的含量,以国家标准物质GBW 07602(GSV-1)为内标控制分析质量,同时做对应空白试验。
1.3 数据处理
植物地上部和根的生物量均为干质量。水稻Cd转运系数为地上部Cd含量与根部Cd含量的比值。
采用Excel进行数据统计,SPSS 19.0软件进行单因素方差分析,Duncan’s新复极差法检验处理间的差异,Origin 9软件作图。
2 结果与分析
2.1 对水稻生长的影响
表1是五丰优2168在2种栽培条件下不同Cd浓度处理下的生物量。由表1可知无论是加Cd处理还是未加Cd处理,水稻根、地上部干质量均为营养液缺氧栽培高于营养液充氧栽培。缺氧栽培条件下,3种Cd浓度处理的根、地上部干质量无显著差异。充氧栽培条件下,0.6和1.2 mg·L–1Cd处理的根、地上部均无显著差异,但相比空白对照,加Cd处理的根、地上部干质量显著下降(P<0.05)。水稻根长度均为充氧栽培高于缺氧栽培,而根的干质量却比缺氧的小,这表明营养液充氧的水稻根比较细长且多。试验结果表明水稻在营养液缺氧和充氧的条件下种植,其生物量和根部的生长状况存在差异,且一定浓度的重金属Cd在充氧的环境下更能抑制水稻的生长。
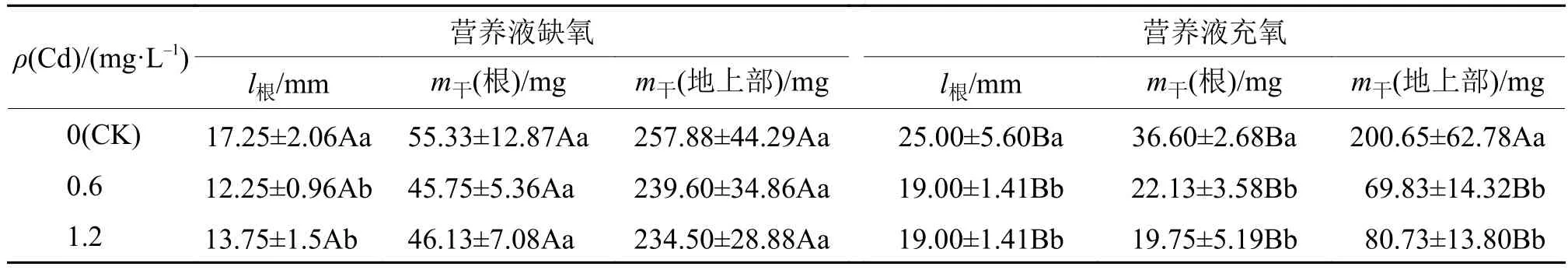
表1 不同Cd处理下水稻的生长状况1)Tab. 1 Growth status of rice plant under different cadmium treatments
2.2 对水稻根和地上部Cd、Zn含量的影响
由图1A可知,在营养液缺氧与营养液充氧2种栽培条件下,水稻根部对重金属Cd的累积量均随着加入的Cd浓度的增大而增加,而营养液充氧栽培的水稻根部的Cd的累积量在Cd质量浓度为0.6和1.2 mg·L–1处理时,都显著高于营养液缺氧栽培的(P<0.05)。缺氧栽培的水稻根部Cd累积量最大为1.2 mg·L–1Cd处理的水稻,累积量为690.8 mg·kg–1,充氧栽培的水稻根部Cd累积量最大值也是1.2 mg·L–1处理的水稻,累积量为1 472.1 mg·kg–1,是缺氧栽培在1.2 mg·L–1处理时累积量的2.13倍。同样的,0.6 mg·L–1Cd浓度处理下,充氧栽培的水稻根部Cd累积量为缺氧栽培水稻Cd累积量的2.21倍。从图1B可知,水稻根部Zn的累积量随Cd处理浓度的增加而减少,且营养液充氧栽培的Zn累积量均显著高于营养液缺氧栽培(P<0.05)。
由图1C可知,2种栽培条件下水稻地上部Cd累积量在加Cd处理时均有显著增加,但在0.6与 1.2 mg·L–1Cd处理下,营养液缺氧栽培的水稻地上部Cd累积量均无显著差异;营养液充氧栽培的水稻地上部在0.6 mg·L–1Cd浓度处理时,Cd累积量为79.92 mg·kg–1;在1.2 mg·L–1Cd浓度处理时,Cd累积量为85.46 mg·kg–1,相比0.6 mg·L–1Cd处理增加了5.54 mg·kg–1。水稻地上部Cd累积量均为充氧栽培高于缺氧栽培。0.6 mg·L–1Cd处理下,充氧栽培的水稻地上部Cd累积量为缺氧栽培的4.73倍;1.2 mg·L–1Cd处理下,充氧栽培的水稻地上部Cd累积量为缺氧栽培的5.22倍。从图1D可知,水稻地上部对Zn的累积量由于加入Cd而减少,且营养液充氧栽培对Zn的累积量均显著高于营养液缺氧栽培的。
2.3 对Cd的转运系数的影响
由图2可知,不同栽培条件和Cd浓度处理的Cd转运系数(即地上部Cd含量与根部Cd含量的比值)存在显著差异(P<0.05)。营养液充氧栽培的Cd转运能力高于营养液缺氧栽培的,同时水稻Cd转运能力均为0.6 mg·L–1Cd处理的大于1.2 mg·L–1Cd处理的。结果表明在一定Cd浓度胁迫下,水稻对Cd的转运会下降,同时营养液缺氧栽培更能抑制水稻Cd转运。
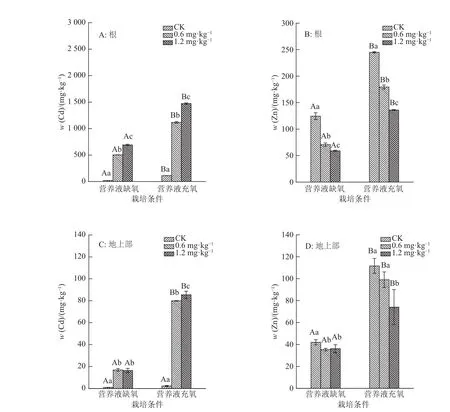
图1 不同栽培条件和Cd浓度处理下根和地上部的Cd、Zn含量Fig. 1 Cd and Zn concentrations in roots and above ground parts of rice under different cultivation conditions and Cd levels
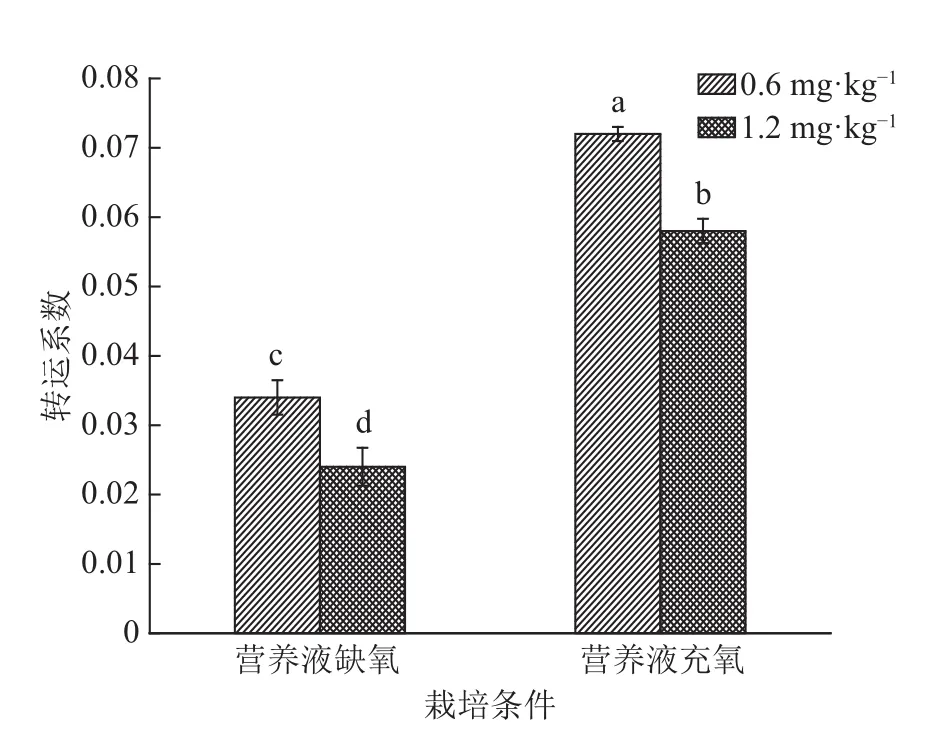
图2 不同栽培条件和不同Cd浓度处理下Cd的转运系数Fig. 2 Transfer coefficients of Cd in rice under different cultivation conditions and Cd levels
2.4 对OsHMA2表达的影响
由图3可知,在2种栽培条件下,水稻茎中OsHMA2的表达量均表现为随着Cd质量浓度的增大先增加后减少,OsHMA2表达量在0.6 mg·L–1Cd浓度处理时均增加,而在1.2 mg·L–1Cd浓度处理时均出现下降。营养液缺氧栽培的水稻茎中OsHMA2的表达量在1.2 mg·L–1Cd处理时减少,是0.6 mg·L–1Cd处理的0.27倍,但相比空白对照表达量是显著上调的。营养液充氧栽培的水稻茎中OsHMA2表达量在1.2 mg·L–1Cd处理时减少,是0.6 mg·L–1Cd的0.23倍,相比空白对照表达量下调但不显著。缺氧栽培的水稻茎中OsHMA2的表达量在Cd质量浓度为 0.6和1.2 mg·L–1时均比营养液充氧栽培的高,在0.6 mg·L–1Cd处理中,缺氧栽培的水稻茎中OsHMA2的表达量是充氧栽培的3.32倍,在1.2 mg·L–1Cd处理中,缺氧栽培的水稻茎中OsHMA2的表达量是充氧栽培的3.89倍。
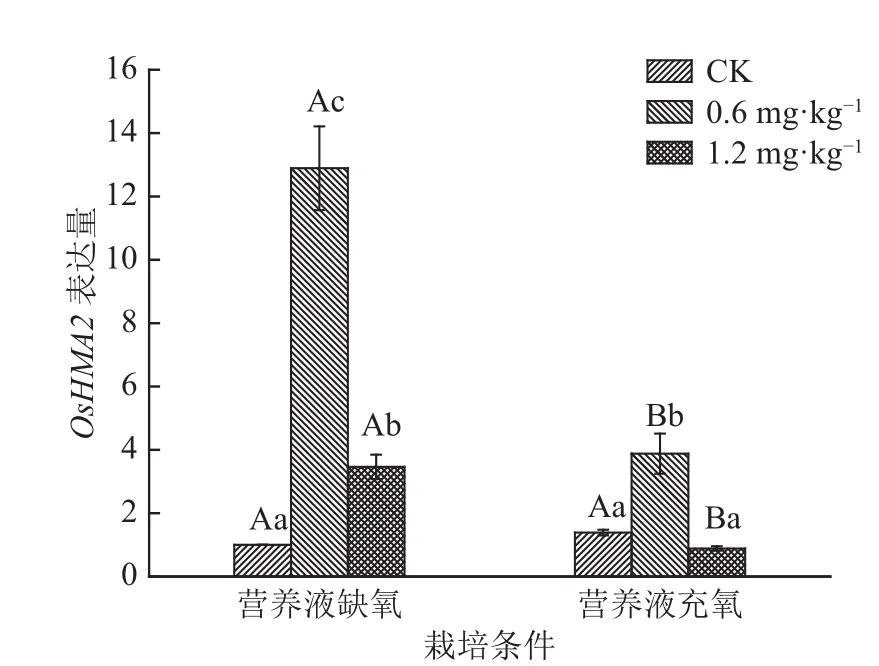
图3 不同栽培条件和不同Cd浓度处理下OsHMA2的表达Fig. 3 Expression of OsHMA2 in shoots of rice under different cultivation conditions and Cd levels
3 讨论与结论
受到重金属Cd的胁迫,植物生长发育会受到影响,表现为植株矮小、叶片发黄[16-17],本研究结果表明,水稻在营养液充氧栽培及Cd长时间处理,根部、地上部的生长发育都会受到显著的抑制,表现为根和地上部干质量下降,说明Cd对水稻生长发育的抑制作用受栽培条件的影响。缺氧和充氧栽培条件下,氧分作用于水稻根部而影响其生长,进而影响根系的活力。研究[18-20]表明,通气条件有利于根系的生长,水稻根变得细长且数量较多,根的比表面积增大,进入植物的Cd增多,抑制作用增强。另有研究[21]表明水稻在淹水缺氧的条件下,根部的缺氧屏障、通气组织以及其他的解剖学特征会显现出来,在缺氧的条件下,水稻的通气组织会在根系的周围释放氧,根系周围的Fe2+就会被氧化成铁膜附着在根的表面[22],而根系表面的铁膜会影响根对重金属的吸收[23-24]。在高浓度Cd(1.2 mg·L–1)处理条件下,营养液缺氧栽培的水稻的干质量相比空白对照只下降了10.46%,而营养液充氧种植的水稻受到Cd的抑制作用较大,下降了57.65%。
在相同处理条件下,同一水稻的不同器官对重金属Cd的吸收和累积存在较大的差异[25]。本研究发现,随着Cd处理浓度的增加,根部Cd的累积量增大,但地上部对Cd的累积达到一定量后并不随着Cd处理浓度的增加而增大,这可能与水稻本身对重金属的抗性和解毒机制有关。Sasaki等[26]研究发现,在Cd胁迫下,水稻根部基因OsHMA3过量表达,部分Cd、Zn等离子转运至液泡中隔离,导致根部Cd和Zn累积量增加,地上部Cd和Zn的累积量反而降低。薛永等[27]认为CDF家族蛋白、AtMRP3、AtPDR8等为重金属解毒蛋白,能将重金属隔离或排出体外,从而减少重金属对植物的毒害。本研究中0.6 mg·L–1Cd处理的转运系数均大于1.2 mg·L–1Cd处理,这表明在高Cd浓度胁迫下,因大部分Cd被固定在根部或是水稻的解毒作用而减少了Cd向地上部的转运。周全等[28]认为水稻植株对Cd的累积量在不同的栽培条件和一定的生长发育阶段存在饱和现象,在高浓度Cd(0.5~2.0 mg·kg–1)处理条件下,水稻植株地上部或者根部的Cd含量增加较少,甚至有些品种出现显著下降。这表明,水稻对Cd的转运系数在一定的Cd浓度胁迫下会出现下降的现象,而饱和的浓度还需要进一步的验证。
OsHMA2是水稻根和地上部运输Zn和Cd的一个转运蛋白[12],本研究结果分析表明水稻对Zn的累积量由于加入Cd而减少,即一定量的Cd对水稻吸收Zn有抑制作用,这与一些研究[29-30]结果相似,表明Cd与Zn存在竞争作用。本研究发现在不同Cd浓度处理的营养液栽培的水稻茎中,OsHMA2表达量在低浓度处理下增加,而在高浓度处理时却减少了,这可能是导致1.2 mg·L–1Cd处理中转运系数降低的原因之一。当水稻地上部分的Cd累积浓度低时,大量OsHMA2表达,从而参与了将Cd从根部向地上部分转运,而当地上部分的Cd累积达到一定量时,水稻可能通过调节相关基因的活性或是组织器官受到不同程度的损伤[31-32],或是通过分泌相关的络合物,降低了Cd和Zn向地上的转运能力,本研究发现,水稻植株在高浓度Cd处理下,OsHMA2的表达量减少,从而出现了Cd的转运系数降低、地上部Cd累积增加较少和Zn的累积量下降的现象。这表明不同氧分栽培条件下,水稻茎中OsHMA2的表达量存在差异,营养液缺氧栽培的水稻茎中OsHMA2的表达量要高于营养液充氧栽培的,同时Cd累积量较少时大量的OsHMA2表达,而当Cd累积量达到一定值时会降低OsHMA2的表达。
[1]COLMER T D. Aerenchyma and an inducible barrier to radial oxygen loss facilitate root aeration in upland, paddy and deep-water rice (Oryza sativa L.)[J]. Ann Bot-London, 2003, 91(2): 301-309.
[2]SATARUG S, BAKER J R, URBENJAPOL S, et al. A global perspective on cadmium pollution and tox-icity in non-occupationally exposed population[J]. Toxicol Lett, 2003, 137(1): 65-83.
[3]王晓娟, 王文斌, 杨龙, 等. 重金属镉(Cd)在植物体内的转运途径及其调控机制[J]. 生态学报, 2015, 35(23): 7921-7929.
[4]UENO D, YAMAJI N, KONO I, et al. Gene limiting cadmium accumulation in rice[J]. Proc Natl Acad Sci, 2010, 107(38): 16500-16505.
[5]CHENG F M, ZHAO N C, XU H M, et al. Cadmium and lead contamination in japonica rice grains and its variation among the different locations in southeast China[J]. Sci Total Environ, 2006, 395(1/2/3): 156-166.
[6]崔玉静, 赵中秋, 刘文菊, 等. 镉在土壤–植物–人体系统中迁移积累及其影响因子[J]. 生态学报, 2003, 23 (10): 2133-2143.
[7]IREMONGER S F, KELLY D L. The responses of four Irish wetland tree species to raised soil water levels[J]. New Phytol, 1988, 109(4): 491-497.
[8]纪雄辉, 梁永超, 鲁艳红, 等. 污染稻田水分管理对水稻吸收积累镉的影响及其作用机理[J]. 生态学报, 2007, 27(9): 3930-3939.
[9]徐加宽, 严贞, 袁玲花, 等. 稻米重金属污染的农艺治理途径及其研究进展[J]. 江苏农业科学, 2007, 35(5): 220-226.
[10]郑绍建, 胡蔼堂. 淹水对污染土壤镉形态转化的影响[J]. 环境科学学报, 1995, 15(2): 142-147.
[11]REDDY C N, PATRICK W H. Effect of redox potential and pH on the uptake of cadmium and lead by rice plants[J]. J Environ Qual, 1977, 6(3): 259-262.
[12]SATOH-NAGASAWA N, MORI M, NAKAZAWA N, et al. Mutations in rice (Oryza sativa) heavy metal ATPase 2 (OsHMA2) restrict the translocation of zinc and cadmium[J]. Plant Cell Physiol, 2012, 53(1): 213-224.
[13]TAKAHASHI R, ISHIMARUY, SHIMO H, et al. The OsHMA2 transporter is involved in root-to-shoot translocation of Zn and Cd in rice[J]. Plant Cell Environ, 2012, 35(11): 1948-1957.
[14]YAMAGUCHI N, ISHIKAWA S, ABE T, et al. Role of the node in controlling traffic of cadmium, zinc, and manganese in rice[J]. J Exp Bot, 2012, 63(7): 2729-2737.
[15]刘志彦, 杨俊兴, 田耀武, 等. 琼脂培养基质条件下砷在水稻幼苗中的积累转运[J]. 安徽农业科学, 2010, 38 (4): 1755-1758.
[16]宋文恩, 陈世宝 .基于水稻根伸长的不同土壤中镉(Cd)毒性阈值(EC_x)及预测模型[J]. 中国农业科学, 2014, 47(17): 3434-3443.
[17]DAS P, SAMANTARAY S, ROUT G R. Studies on cadmium toxicity in plants: A review[J]. Environ Pollut, 1997, 665(1): 29-36.
[18]朱智伟, 陈铭学, 牟仁祥, 等. 水稻镉代谢与控制研究进展[J]. 中国农业科学, 2014, 47(18): 3633-3640.
[19]秦华东, 江立庚, 肖巧珍. 水分管理对免耕抛秧水稻根系生长及产量的影响[J]. 中国水稻科学, 2013, 27 (2): 209-212.
[20]沈阿林, 刘春增, 张付申. 不同水分管理对水稻生长与氮素利用的影响[J]. 植物营养与肥料学报, 1997(2): 111-116.
[21]COLMER T D, COX M C, VOESENEK L A. Root aeration in rice (Oryza sativa): Evaluation of oxygen, carbon dioxide, and ethylene as possible regulators of root acclimatizations[J]. New Phytol, 2006, 170(4): 767-778.
[22]ARMSTRONG W. Oxygen diffusion from the roots of some britishbogplants[J]. Nature, 1964, 204: 801-802.
[23]ZHANG X K, ZHANG F S, MAO D R. Effect of iron plaque outside roots on nutrient uptake by rice (Oryza sativa L.): Zinc uptake by Fe-deficient rice[J]. Plant Soil, 1998, 202(1): 33-39.
[24]胡莹, 黄益宗, 黄艳超, 等. 不同生育期水稻根表铁膜的形成及其对水稻吸收和转运Cd的影响[J]. 农业环境科学学报, 2013, 32(3): 432-437.
[25]刘侯俊, 梁吉哲, 韩晓日, 等. 东北地区不同水稻品种对Cd的累积特性研究[J]. 农业环境科学学报, 2011, 30(2): 220-227.
[26]SASAKI A, YAMAJI N, MA J F, et al. Overexpression of OsHMA3 enhances Cd tolerance and expression of Zn transporter genes in rice[J]. J Exp Bot, 2014, 65(20): 6013-6021.
[27]薛永, 王苑螈, 姚泉洪, 等. 植物对土壤重金属镉抗性的研究进展[J]. 生态环境学报, 2014, 22(3): 528-534.
[28]周全, 王宏, 张迎信, 等. 不同镉浓度处理下水稻植株镉含量变化及其镉调控相关基因表达分析[J]. 中国水稻科学, 2016, 30(4): 380-388.
[29]KUKIER U, CHANEY R L. Growing rice grain with contronlled cadmium concentrations[J]. J Plant Nutr, 2002, 25(8): 1793-1820.
[30]龚伟群, 李恋卿, 潘根兴. 杂交水稻对Cd的吸收与籽粒积累: 土壤和品种的交互影响[J]. 环境科学, 2006, 27(8): 1647-1653.
[31]MOULIS J M. Cellular mechanisms of cadmium toxicity related to the homeostasis of essential metals[J]. Biometals, 2010, 23(5): 877-896.
[32]刘耀明, 余志涛, 朱文雅, 等. 三种重金属对中华稻蝗金属硫蛋白基因表达的影响[J]. 农业环境科学学报, 2015, 34(2): 227-232.
【责任编辑 霍 欢】
Effects of anoxic and aerobic cultivation conditions on cadmium accumulation and OsHMA2 expression in rice
YANG Yaoshuai, MEI Xiuqin, LI Qusheng, WEI Jia, ZHOU Xuefang, CHEN Kehan, ZHOU Ting
(School of Environment, Jinan University/Key Laboratory of Environmental Exposure and Health of Guangzhou City/Guangdong Provincial Research Center of Environmental Pollution Control and Remediation Material, Guangzhou 510632, China)
【Objective】To study the effects of anoxic and aerobic cultivation conditions on cadmium (Cd) uptake and transfer and OsHMA2 expression in rice shoots, and provide a theoretical basis for reducing the uptake and accumulation of heavy metals in rice.【Method】Using rice cultivar ‘Wufengyou 2168’, pot experiments were performed hydroponically under anoxic(nutrient solution with agar) and aerobic(oxygenized nutrient solution) conditions. Cd uptake and accumulation, and OsHMA2 expression levels in rice shoots were analyzed at three Cd levels(0, 0.6, 1.2 mg·L–1).【Result】Rice growth was not significantly inhibited by Cd under anoxic condition, but was significantly inhibited under aerobic condition with the dry weight of roots and above ground parts significantly decreased. At the Cd concentration of 0.6 and 1.2 mg·L–1, Cd accumulations in roots and above ground parts of rice under anoxic conditionwere lower than those under aerobic condition. Under both anoxic and aerobic conditions, Cd accumulation in roots increased with Cd concentration. For above ground parts of rice, Cd accumulation was not significantly different between 0.6 and 1.2 mg·L–1Cd levels under anoxic condition, but increased with Cd concentration under aerobic condition. Compared with the control (0 mg·L–1Cd), OsHMA2 expression level increased at the Cd concentration of 0.6 mg·L–1but decreased at 1.2 mg·L–1in shoots under both anoxic and aerobic condition, and OsHMA2 expression level in rice was higher under anoxic condition than those aerobic condition at each Cd level.【Conclusion】Anoxic culture can inhibit Cd uptake and accumulation in rice, and OsHMA2 expression decreases when Cd accumulation reaches a certain value.
rice; anoxic codition; aerobic condition; Cd; OsHMA2; transfer coefficient
Q945;S511
A
1001-411X(2017)05-0024-06
杨耀帅, 梅秀芹, 李取生, 等. 缺氧和充氧栽培对水稻Cd积累及OsHMA2基因表达的影响[J]. 华南农业大学学报, 2017, 38(5): 24-29.
2017-02-28 优先出版时间:2017-07-14
优先出版网址:http://kns.cnki.net/kcms/detail/44.1110.s.20170714.0855.010.html
杨耀帅(1990—),男,硕士研究生,E-mail: 313247890@qq.com; 通信作者: 梅秀芹(1978—),女,副研究员,博士,E-mail: twindymei@jnu.edu.cn;李取生(1965—),男,教授,博士,E-mail: liqusheng@sina.com
国家自然科学基金(41401365);中国博士后科学基金 (2015T80939, 2014M552282)
