大渡河干旱河谷四川牡丹种群结构特征及与环境因子的关系
2017-08-07夏小梅彭培好刘贤安
夏小梅,王 娟,彭培好,刘贤安
(成都理工大学 生态资源与景观研究所,四川 成都 610059)
大渡河干旱河谷四川牡丹种群结构特征及与环境因子的关系
夏小梅,王 娟,彭培好,刘贤安
(成都理工大学 生态资源与景观研究所,四川 成都 610059)
2014年对四川省阿坝州四川牡丹分布较为集中的地方设置19个样方进行调查,根据其数据编制静态生命表,采用主成分分析等方法对大渡河干旱河谷四川牡丹种群结构、种群特征与环境因子的关系进行研究。结果表明,不同生境的四川牡丹种群年龄级分布存在一定差异,在以灌木为优势种的群落生境中,由于郁闭度低、透光率高,四川牡丹幼苗数量多,种群的更新状态较好,属于增长型或稳定型种群;在以乔木为优势种的群落生境中,四川牡丹幼苗死亡率高,种群属于衰退型;主成分分析表明,四川牡丹平均基径和种群密度受环境因子影响显著,其中坡度是影响四川牡丹种群特征的首要环境因子,其次是土壤自然含水率。
大渡河干旱河谷;四川牡丹;种群结构特征;环境因子;主成分分析
四川牡丹Paeonia szechuanica为毛茛科Ranunculaceae芍药属Paeonia落叶小灌木,属国家Ⅱ级重点保护植物[1],主要分布于岷江上游、大渡河两岸的马尔康、金川、丹巴、康定等地[2]。目前,关于四川牡丹的研究主要集中在种子萌发[3-4]、种子生理生化特性[5]等方面,对四川牡丹种群与环境因子关系的研究尚未见报道。四川牡丹是大渡河干旱河谷的乡土物种之一。本文通过研究不同群落类型中的四川牡丹种群结构以及环境因子对其种群结构的影响,为四川牡丹种群保护措施的制定提供科学依据,同时为大渡河干旱河谷区植被恢复工作的开展提供一定的理论基础。
1 研究地区与研究方法
1.1研究地概况
研究区位于四川省阿坝藏族羌族自治州马尔康县、金川县、小金县,102°01′ ~ 103°16′E,30°20′ ~ 31°59′N,海拔2 204 ~ 2 880 m。属亚热带季风气候,年均降水量600 ~ 650 mm,年均蒸发量1 490 ~ 2 510 mm,干湿季节分明,干旱时间长。土壤主要是山地燥红土、燥褐土,具有熟化度低、土层薄、石砾多、渗透性强、保水保肥性差,有机质含量低、贫氮、缺磷、富钾的特点[6-7]。
1.2研究方法
1.2.1 取样方法及野外调查
根据国家林业局《第二次重点保护野生植物资源调查实施细则》,在研究区域内选取6个四川牡丹分布较为集中、群落类型具有一定差异的点,分别记录各个点的地点、海拔、生境,群落物种等环境因子(表1)。根据每个点四川牡丹的分布情况,共设置19个10 m ×10 m的样方,测定经纬度,记录海拔、坡度、坡向、坡位、土壤自然含水率、pH、乔木层盖度、人为干扰率共8个环境因子,统计每样方四川牡丹的株数,测定冠幅(C)、基径(D)和高度(H),株龄用茎上芽鳞痕的计数方法逐株查记。

表1 大渡河干旱河谷区四川牡丹样地概况Table 1 Sample information of P.szechuanica
1.2.2 数据统计分析
根据野外调查数据,统计四川牡丹各个龄级的个体数量,绘制6个样地的年龄结构图,判断不同群落类型中四川牡丹种群的未来发展趋势。编制6个样地四川牡丹静态生命表,从级间死亡率、龄级个体的平均期望寿命、致死力等方面分析该种群结构动态与更新现状[8]。对19个样方四川牡丹种群特征,包括平均基径和种群密度的数据进行统计,利用SPSS21.0 统计分析软件中的CCA(Canonical correspondence analysis)模块分析四川牡丹基径和种群密度与8个环境因子的相关性,同时对8个环境因子进行主成分分析(Principal component analysis/PCA),得到四川牡丹基径和种群密度与4个主份量的相关性,筛选出生境状况的主要环境因子。
2 结果与分析
2.1四川牡丹种群结构
据样地调查资料,以各年龄级种群相对存活量和年龄级为坐标,绘制6个样地种群年龄结构图(图1)。由图1可知,不同样地中种群的年龄级分布存在一定差异。以川西锦鸡儿、金露梅为群落优势种的样地1、样地3、样地5中,四川牡丹种群年龄结构呈金字塔型,个体集中在中龄阶段,老龄和幼龄个体所占比例相对较少,属于稳定性种群;以样地4为代表的群落环境中,岷江柏木、川楝等乔木数量较多,透光率较低,四川牡丹个体集中在幼龄阶段,中龄和老龄个体数量极度减少,种群的更新状态较差,不利于四川牡丹的种群结构发展,属于衰退型种群。以金花小檗、金露梅、小果蔷薇、平枝栒子等为群落优势种的样地2和样地6中,四川牡丹各龄级个体数量多,种群的更新状态较好,种群结构比较稳定,属于增长型种群。
通过编制四川牡丹静态生命表(表2),分析四川牡丹种群动态,内容包括:单位时间年龄等级的中值(x)、x龄级内现有个体数(ax)、修正值(ax´)、在x龄级开始时标准化存活个体数(lx)(一般均转化为1 000)、从x到x+1龄级间隔期内标准化死亡数(dx)、从x到x+1龄级间隔期间死亡率(qx)、进入x龄级个体的生命期望或平均期望寿命(Ex)、致死力(Kx)等。

图1 不同样地四川牡丹种群年龄结构Figure1 Age class structure of Paeonia decomposita in different community environment
根据表2可知,样地1、样地2、样地3、样地5、样地6四川牡丹种群大龄级相对小龄级有较高的死亡率,说明上述种群的更新较旺盛,种群不断发展。相反,样地4 四川牡丹种群小龄级死亡率较高,表明种群更新不良,说明在岷江柏木、川楝为群落优势种的郁闭林下四川牡丹种群幼苗阶段自我更新较差的特点。样地1、样地2、样地3、样地5、样地6种群各龄级植株的生命期望值,幼苗幼株的最高,表明当地环境条件适宜四川牡丹的存活生长。但随着种间竞争的逐步增强,生命期望寿命也逐渐降低。样地4中,四川牡丹幼苗的期望寿命相对较低,与实地调查发现,人为干扰是影响四川牡丹幼苗期望寿命的主要因素。
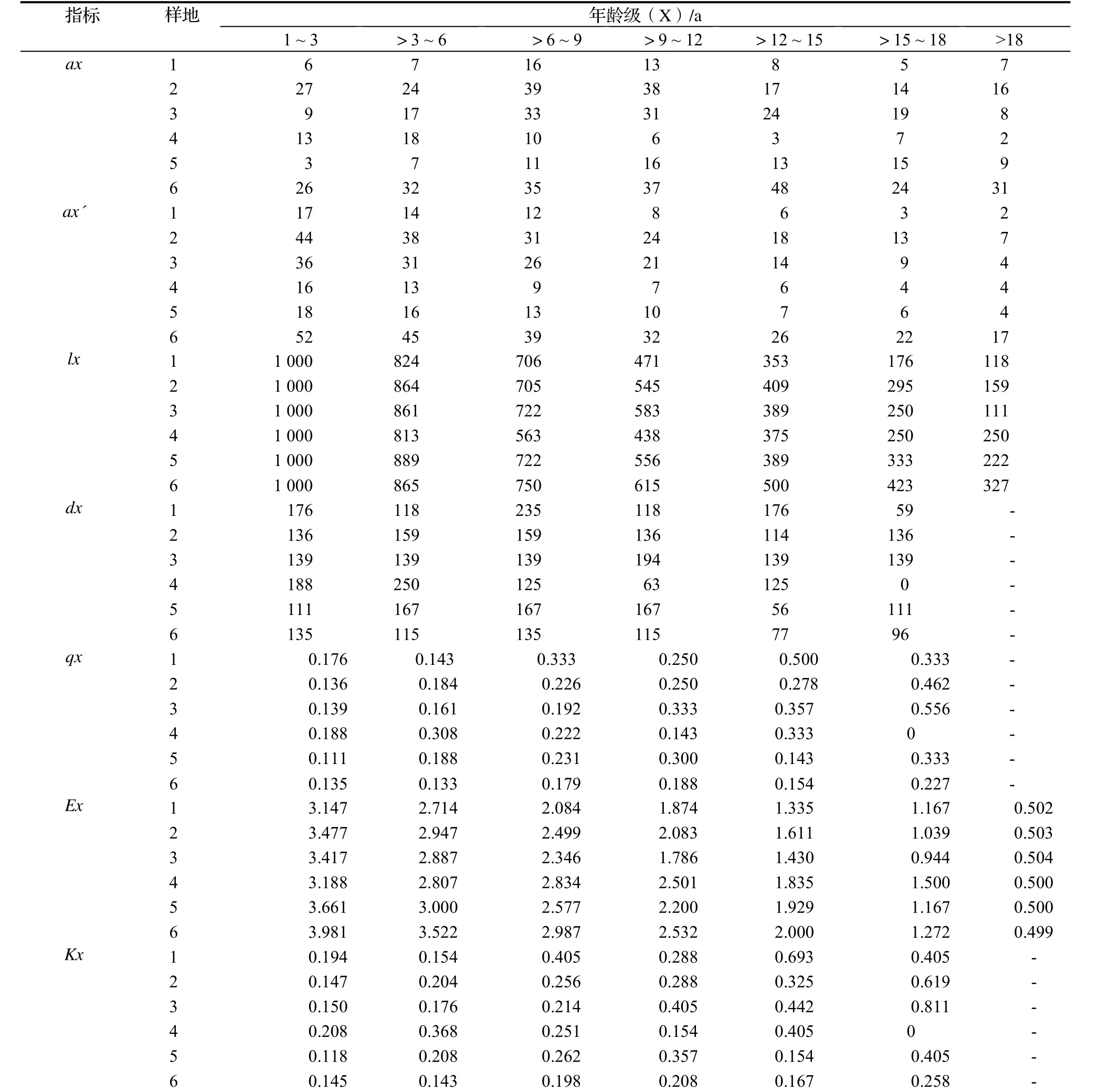
表2 不同样地四川牡丹静态生命表Table 2 Static life table of P. szechuanica in different community
2.2四川牡丹种群特征与环境因子的关系
对19个样方的经纬度、四川牡丹种群特征,包括种群密度和平均基径进行统计(表3)。种群密度可分为5个等级:1级,种群密度≤0.5株·m-2;2级,种群密度在>0.5 ~ 1.0株·m-2;3级,种群密度在>1.0 ~ 1.5株·m-2;4级,种群密度在>1. 5 ~ 2.0株·m-2;5级,种群密度>2.0株·m-2。
利用SPSS21.0软件对19个样方的8个环境因子的原始数据进行主成分分析,然后选取环境因子特征值较高的主分量PC1,PC2,PC3,PC4,特征值分别是2.688,1.786,1.215和1.078,贡献率分别为33.600%,22.320%,15.183%和13.470%,累积贡献率为84.572%。以选取较高贡献的主分量为基础,绘制四川牡丹种群特征与PCA主份量的相关性(见图2),由图2可知,四川牡丹平均基径和种群密度与PC1,PC2和PC3不存在显著相关,与PC4呈显著和极显著相关。由此可知,四川牡丹平均基径和种群密度受环境因子影响显著。
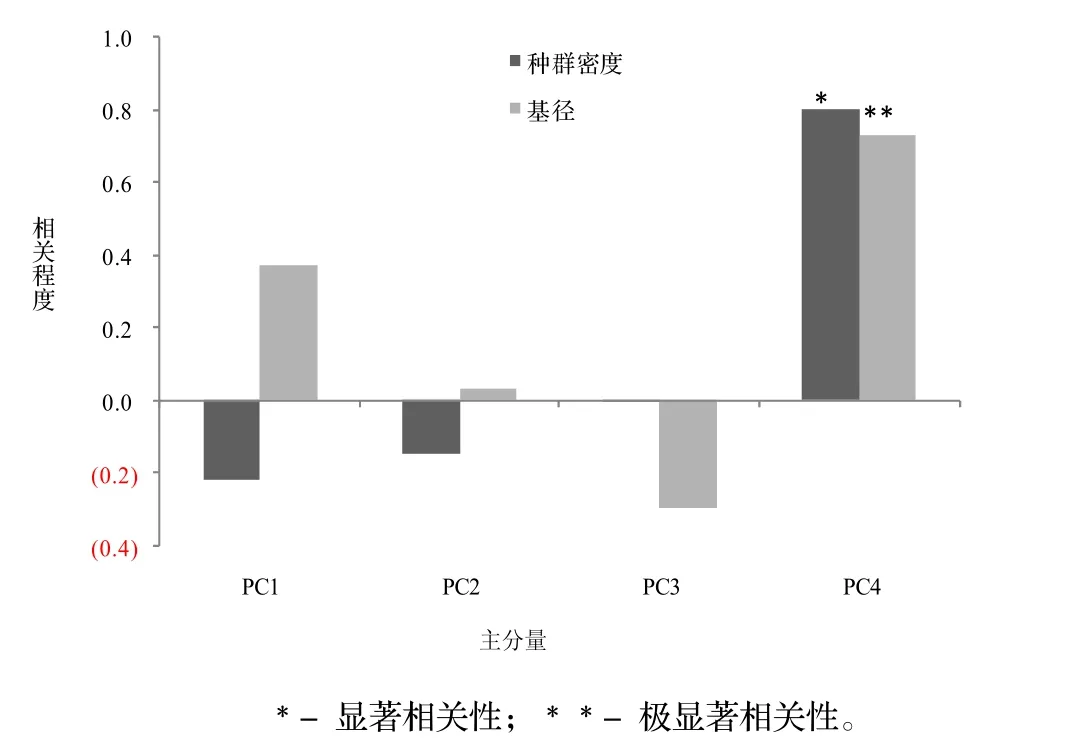
图2 四川牡丹种群特征与PCA主份量的相关性Figure 2 Correlation between population properties of P. szechuanica and principal components of PCA
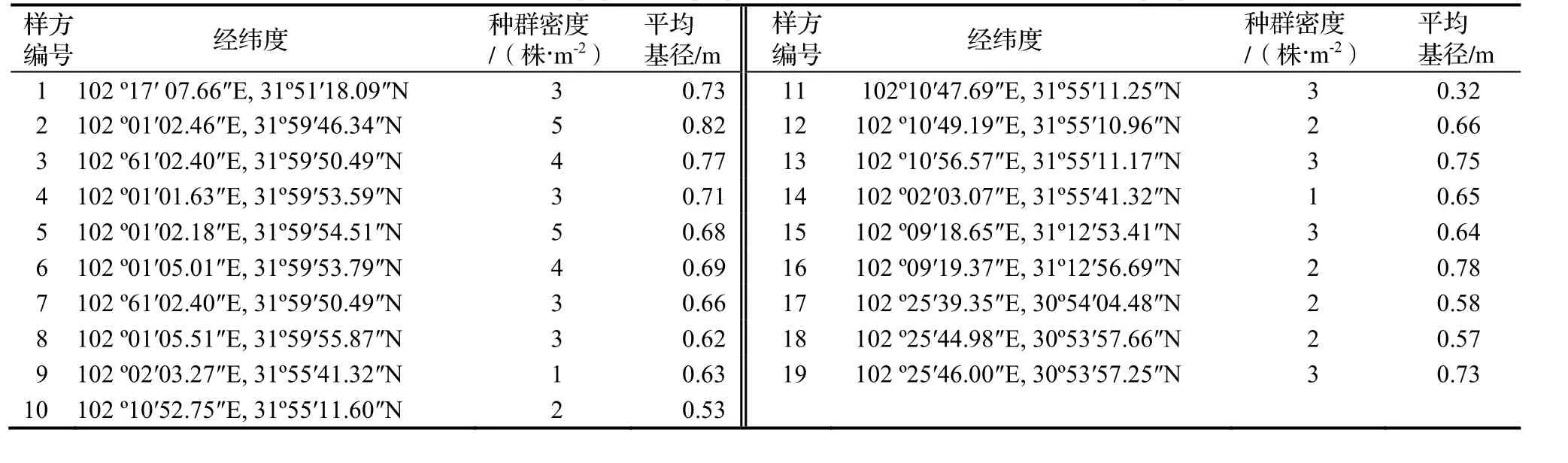
表3 大渡河干旱河谷四川牡丹分布点及种群特征Table 3 Distribution and population properties of P. szechuanica in different sample plots
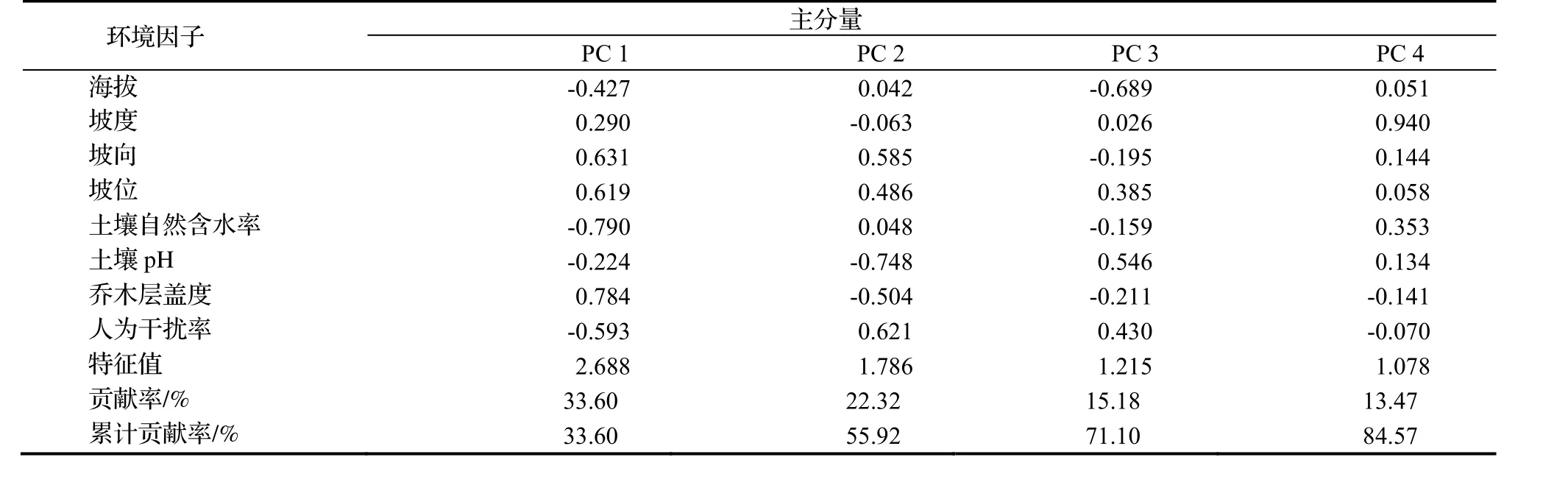
表4 四川牡丹生境中各环境因子的主成份分析结果Table 4 Principal component analysis on environmental factors of P. szechuanica habitats
表4为8个环境因子对主分量的贡献率。由表4可知,第1轴负荷量较大的4个因子是土壤自然含水率(-0.790)、乔木层盖度(0.784)、坡向(0.631)、坡位(0.619);第2轴负荷量较大的2个因子是土壤pH(-0.748)、人为干扰率(0.621);第3轴负荷量较大的2个因子是海拔(-0.689)。第4轴负荷量较大的的因子是坡度(0.940),结合图3可知,地形因子中坡度是影响大渡河干旱河谷区四川牡丹平均基径和种群密度的首要环境因子,其次是土壤自然含水率。
3 结论与讨论
植物种群结构可反映种群的数量动态、发展趋势与环境之间的相互关系[9],也可揭示植物种群与生境相适应的结果,对预测群落未来发展趋势具有重要意义[10-14]。四川牡丹年龄结构在不同样地中有较大差异,各样地存在数量不等的幼龄个体,以乔木为优势种的群落环境,具有郁闭度高、透光率低的特点,四川牡丹种群呈现出明显的衰退迹象;以灌木为优势种的生境中,郁闭度低透光率较高,四川牡丹种群更新情况良好。有研究表明,郁闭度的大小会对种群幼苗更新的光环境形成直接影响,从而影响植物种群幼苗的更新[19-20],但人为干扰导致种群生境的破碎化是种群走向衰退的主要原因[17-18],与本文对四川牡丹种群结构的研究结果一致。
通过环境因子的主成分分析结果表明,四川牡丹平均基径和种群密度受环境因子显著影响,地形因子中坡度是影响四川牡丹种群特征的最主要因子,其次是土壤自然含水率。植物种群特征通常由最大限制环境因子决定,因此,不同植物种群特征与环境因子关系的研究结果并不一致,如有些学者研究表明乔木层郁闭度、土壤pH、人为干扰等是影响物种种群特征的重要环境因子[19-20]。
四川牡丹为大渡河干旱河谷区特有物种,旅游、放牧、祭祀、采摘等人为活动已经严重破坏了四川牡丹生长环境。针对不同群落类型的四川牡丹种群结构及种群特征与环境因子的关系,在郁闭度较高,透光率较差的群落可适当砍伐非主要树种,透光率较好的群落应以保护原有生境为主,同时禁止或减少人类破环活动,为四川牡丹种群的良好生长提供有利条件。另一方面,对乡土植物四川牡丹种群结构及种群与环境因子关系的研究也可为干旱河谷区植被恢复工作的顺利开展提供一定理论基础。
[1] 傅立国主编. 中国植物红皮书:稀有濒危植物第一册. 北京:科学出版社,1999,530-537.
[2] 洪德元,潘开玉.芍药属牡丹组的分类历史和分类处理[J]. 植物分类学报,1999,37(4):351-368.
[3] 景新明,郑光华,裴颜龙,等. 野生紫斑牡丹和四川牡丹种子萌发特性及与其致濒的关系[J]. 生物多样性,1995,3(2):84-87.
[4] Song H X, Liu G L, Chen Q B. Effects of aqueous extracts of Paeoniadecomposita seeds on germination and some metabolic activities associated with growth of wheat seedlings[J]. Acta Biol Hung,2012,63(3):362-71.
[5] 汪源,刘光立,张倩,等. 层积处理对四川牡丹种子生理生化特性的影响[J]. 北方园艺,2013,24:59-62.
[6] 谢学强. 干旱河谷不同立地条件对引种湿热型植物的影响[J]. 贵州农业科学.2011,39(12):202-204.
[7] 谢学强,肖晓凤,秦丹,等. 大渡河干旱河谷野生半夏及其驯化栽培技术[J]. 现代园艺,2014,(19):25-26.
[8] 江洪. 云杉种群生态学[M]. 北京:中国林业出版社,1992.
[9] 李奎,郑宝强等. 滇牡丹自然种群数量动态[J]. 植物生态学报,2012,06:522-529.
[10] Khan D, Shaukat S S. The population structure, intraspecific competition and phasic development of growth in a population of Urochondra setulosa (Trin.) C.E Hubb. a coastal halophytic grass of pakistan[J]. Pakist J Bot,1997,29(2):271-288.
[11] Svensson J S, Jeglum J K. Structure and dynamics of an undisturbed old-growth Norway spruce forest on the rising Bothnian coastline[J]. For Eco Manage,2001,151(1/3):67-79.
[12] Fuchsa M A, Krannitzb P G, Harestad A S. Factors affecting emergence and first-year survival of seedlings of Garry oaks(Quercus garryana)in British Columbia[J]. Can For Ecol Manage,2000,137(1 /3):209-219.
[13] Molles M C. Ecology: Concept and Applications.2nd [M]. New York:McGraw-Hill Companies,2002.
[14] Wu J X, Zhang X M, Deng C Z, et al. Structure and dynamic of Populus euphratica population along Tarim River[J]. Pakist J Bot,2012,44(5):1651-1656.
[15] 王永杰,张首军. 不同郁闭度下天然白皮松林更新的研究[J]. 山西师范大学学报:自然科学版,2008,22(4):83-85.
[16] 李永宁,张宾兰等. 郁闭度及其测定方法研究与应用[J]. 世界林业研究,2008,21(1):40-46.
[17] 陈国科,彭华. 不同人为干扰条件下毒药树种群数量特征的比较[J]. 植物生态学报,2006,30(3):426-421.
[18] 尚进,李旭光. 人为干扰对桫椤群落植物种间协变影响的研究[J]. 西南师范大学学报:自然科学版,2003,28(6):947-951.
[19] 张文辉,王延平等. 太白红杉种群结构与环境的关系[J]. 生态学报,2004,01:41-47.
[20] 张小平,郝朝运等. 濒危植物永瓣藤的种群结构及与环境的关系[J]. 应用生态学报,2008,03:474-480.
Relationship of Population Structure of Paeonia szechuanica with Environmental Factors in Sichuan Province
XIA Xiao-mei,WANG Juan,PENG Pei-hao,LIU Xian-an
(Institute of Ecological Resources and Landscape, Chengdu University of Technology, Chengdu 610059, China)
Investigations were implemented in 2014 on 19 samples plots of Paeonia szechuanica communities relatively more distributed in Aba, Sichuan province. Static life table was compiled based on survey data. Relationship of population structure and properties with environmental factors was analyzed by principal component analysis. The results showed that age class distribution of P. szechuanica population had difference among different sample plots. At habitats dominated by bush species, P. szechuanica had more seedlings, indicating better regeneration and belonging to stable and growing population. At habitats dominated by tree species, P. szechuanica had higher death rate of seedlings, indicating declining population. Principal component analysis showed that the average ground diameter and population density of P. szechuanica was significantly affected by environmental factors, especially slope and soil moisture content.
dry valley of Dadu River; Paeonia szechuanica; population structure; environment factors; principal component analysis
S718.54
A
1001-3776(2017)02-0030-06
10.3969/j.issn.1001-3776.2017.02.005
2016-09-23;
2017-03-09
阿坝州重点保护野生植物资源调查(80303-SZA001)
夏小梅,硕士研究生,从事植物生态研究;E-mail:905878484@qq.com。通信作者:彭培好,博士生导师,从事植物生态研究;E-mail:peihaop@163.com。
