三种药用甘草种子位置效应对幼苗建植特性的影响
2017-07-21韩春陆嘉惠周玲玲牛清东宋凤王跃杨旭
韩春,陆嘉惠,2,3*,周玲玲,牛清东,宋凤,王跃,杨旭
(1.石河子大学生命科学学院,新疆 石河子 832003;2.新疆兵团绿洲生态农业重点实验室,新疆 石河子 832003;3.石河子大学甘草研究所,新疆 石河子 832003)
三种药用甘草种子位置效应对幼苗建植特性的影响
韩春1,陆嘉惠1,2,3*,周玲玲1,牛清东1,宋凤1,王跃1,杨旭1
(1.石河子大学生命科学学院,新疆 石河子 832003;2.新疆兵团绿洲生态农业重点实验室,新疆 石河子 832003;3.石河子大学甘草研究所,新疆 石河子 832003)
通过对甘草属3种植物,光果甘草、胀果甘草、乌拉尔甘草植株不同部位种子的萌发特性、幼苗生长特性、叶绿素含量、叶绿素荧光参数的测定和比较,探讨了3种药用甘草植株不同部位种子的萌发策略、幼苗的生长适应性,为更清楚地认识甘草属植物种子萌发与幼苗建植策略提供科学依据。结果表明,1)光果甘草植株中、上部,胀果甘草和乌拉尔甘草植株中、下部种子具有较高的出苗率,是种子繁殖的主要保障,但存活率无差异。2)具有较高出苗率的种子萌发形成的幼苗具有较大的生物量、较长的根长,且根冠比也相对较高,其分配到地下部分生物量多于地上部分,发达的根系可增强幼苗的抗逆性和对环境的适应性;而地上部分叶片的叶绿素相对含量、可变荧光(Fv)、最大荧光产量(Fm)、PSⅡ光化学效率(Fv/Fm)、PSⅡ潜在的光化学活性(Fv/F0)和光反应阶段电子传递的速率也相对较高,荧光动力学曲线也反映出其具有较强的荧光强度、电子捕获和光能利用率,进而增强了幼苗建植能力。3)具有较低出苗率的种子萌发形成的幼苗具有较高的株高,较多的叶片数,可以通过优先占领空间和光源来维持生长。综上所述,3种药用甘草不同位置的种子具有不同种子萌发和幼苗生长策略,这对野生甘草自然种群的幼苗建植和种群更新有着重要的生态学意义。
药用甘草;种子;位置效应;幼苗建植
众所周知,种子萌发、幼苗的生长和建植过程是植物生活史中最重要的阶段,直接影响着种群的扩大和延续。长期以来人们热衷于关注外界环境条件对种子萌发和幼苗生长的影响,如温度[1]、光照[2]、竞争[3]和生境[4]等,也有关于对种子大小的研究,证实了大种子比小种子具有更高的萌发率、出苗率、存活率、幼苗生长速度和竞争能力等[5-7]。但亲本种子生产的位置效应对种子萌发和幼苗建植的影响往往被忽略,有文献表明花序内的部位差异会影响幼苗的存活率[8],Buide[9]也发现二歧聚伞花序草本植物Sileneacutifolia近轴端花序的种子萌发率最低,而远轴端花序的种子萌发率最高。谢田朋等[10]通过对黄帚橐吾(Ligulariavirgaurea)幼苗建植的研究发现,黄帚橐吾花序基部种子的萌发开始时间比顶部种子早,幼苗的根冠比和光合速率表现出顶部大于基部的变化趋势,这说明植株不同部位种子存在差异,且部位间种子的萌发和幼苗生长对策不同。
豆科(Leguimnosae)甘草属(Glycyrrhiza)植物全世界约20种[11-12],光果甘草(Glycyrrhizaglabra)、胀果甘草(Glycyrrhizainflata)、乌拉尔甘草(Glycyrrhizauralensis)是《中国药典》[13]收载的药用甘草的3种甘草原植物。目前对这3种药用甘草的有性生殖过程的相关报道较多,如甘草属植物的传粉机制和繁育系统[14]、有性生殖过程及生殖败育[15]、种子结实特性[16]等。前期对甘草属几种植物繁殖资源分配模式和种子生产格局的研究也发现,3种药用甘草(乌拉尔甘草、胀果甘草、光果甘草)种子生产模式存在位置效应[17],在植株不同部位繁殖资源投入和繁殖成功有明显差异。但这种种子生产的位置效应是否对甘草子代幼苗建植也有影响,尚不清楚,综合种子生产和幼苗建植能力的分析,才能更全面理解和认识甘草属植物有性繁殖的“位置效应”的生态学内涵。本研究通过室内种子萌发和盆栽控制实验,分析植株不同部位种子的萌发、幼苗生长状况和光合能力,探讨种子生产的位置效应对子代建植能力的影响,具体拟解决以下2个问题:1)植株不同部位的种子萌发特性、幼苗存活及生长特性是否也存在差异?2)如果存在,产生这种现象的原因可能是什么,对种群建植和物种更新的适应意义是什么?
1 材料与方法
1.1 研究区概况
本研究选取种植30年左右,处于半野生状态的甘草居群。光果甘草、乌拉尔甘草居群位于石河子大学农学院试验站(123°45′ E,44°45′ N),胀果甘草居群位于石河子150团(123°45′ E,45°6′ N)。海拔350~450 m,年平均气温6~10 ℃,无霜期平均155~186 d,≥10 ℃年有效积温3000~3500 ℃,平均年降水量300~500 mm,平均年蒸发量1500~2000 mm。
1.2 实验材料
甘草属植物有几个至数十个腋生总状花序,小花排列稠密,花序长度约为5~20 cm(不同种甘草有差异)。开花顺序由下向上渐次开花,每个小花均产生一个果荚,胚珠多数,果熟期9-10月。于2014年9-10月,在果熟期采集完整植株,带回实验室进行取样。3种甘草各选取5株完整植株作为5个重复,从植株下部第一个花序着生部位开始,将植株平均分为上、中、下3部分,取种子备用。
1.3 实验方法
1.3.1 种子萌发试验 本研究种子采集自野生、半野生状态下的甘草属3种植物(光果甘草、胀果甘草、乌拉尔甘草)[18],每种甘草标记5株,为5个重复;每株植株各部位随机选取30粒无虫害的种子,共计90粒,每种甘草5个重复,共计450粒种子,进行种子萌发和幼苗生长实验。用85%的浓H2SO4处理30~90 min后,用无菌水冲洗3遍,0.1%的HgCl2消毒10 min。然后进行室内盆栽试验,每盆种15粒种子。用实验站周围草甸土作为盆内基质,过筛除杂以除去其他杂质和种子库。花盆(内径20 cm,深度20 cm)装土后再在土上盖一层沙。将甘草种子均匀撒在沙上后再盖一层细沙以防止种子被水冲走。室内温度(25±5) ℃,每天光照12 h (10:00-22:00),隔2 d浇水一次,每天统计一次种子萌发数、幼苗出土数和幼苗死亡数,10 d后计算不同部位种子的出苗率,50 d后统计存活率。
1.3.2 幼苗建植指标测定
出苗率(emergence percentage)=(每盆萌发出土的幼苗总数/每盆播种的种子总数)×100%
存活率(survival percentage)=(每盆试验结束前建成植株的个体总数/每盆出土的幼苗总数)×100%
1.3.3 叶绿素含量和叶绿素荧光动力学参数测定 播种50 d后,每种甘草每个部位各随机选取50株健康幼苗,编号,对每株幼苗单独进行叶绿素含量和叶绿素荧光动力学参数测定,每株幼苗取倒数第2片完全展开的叶片,避开中脉进行测定。用英国产叶绿素测定仪(SPAD)测定叶绿素相对含量。用连续激发式荧光仪(PEA, Hansatech, 英国)进行叶绿素含量和叶绿素荧光动力学参数测定。叶绿素荧光动力学参数测定前叶片暗处理20 min,测定时间为9:30-12:30。叶绿素荧光主要包括以下参数:F0(初始荧光)、Fm(最大荧光)、Fv(可变荧光,Fv=Fm-Fo)、Fv/Fm(原初光能转化率)和Fv/Fo(PSⅡ潜在的光化学活性)。
1.3.4 幼苗生长指标的测定 每株幼苗单独进行生长指标的测定,包括叶片数、株高(地上部分高度)、根长、地上部分生物量(鲜重)、地下部分生物量(鲜重)。生物量用OHAUS(上海奥豪斯有限公司)1/1000电子天平进行称量。
根冠比(root-shoot ratio)=地下部分的生物量/地上部分的生物量×100%
1.4 数据处理
用Excel软件整理所有数据,通过SPSS 19.0统计分析。采用One-way ANOVA分析方法检测植株不同部位间差异的显著性,采用Duncan新复极差法(SSR)进行多重比较,分析3种甘草植株不同部位的种子萌发情况、生物量、根冠比、株高、根长、叶片数和叶绿素荧光参数的差异(P=0.05)。用Bivariate Correlation对叶绿素含量与生物量的相关性进行分析,找到最适回归模型,然后采用幂函数(power)Y=aXb回归方程对幼苗叶绿素含量与生物量之间的相关性进行分析。用Origin 9.0绘图。
2 结果与分析
2.1 植株不同部位种子萌发及幼苗存活情况
由图1A可看出,光果甘草植株中上部种子的出苗率高于植株下部种子的出苗率,胀果甘草和乌拉尔甘草植株中下部种子的出苗率高于植株上部种子的出苗率。由图1B可以看出,3种甘草植株不同部位种子的存活率均无显著差异,说明种子生产的位置效应影响种子的萌发及幼苗存活。
2.2 种子生产的位置效应对幼苗生物量与生长特性的影响
2.2.1 对幼苗生物量和根冠比的影响 由图2A可看出,光果甘草植株中上部种子萌发形成的幼苗总生物量高于植株下部种子萌发形成幼苗的总生物量,而胀果甘草和乌拉尔甘草植株中下部种子萌发形成的幼苗总生物量高于植株上部种子萌发形成幼苗的总生物量。由图2B可以看出,光果甘草植株中上部种子萌发形成的幼苗根冠比相对较高,与植株下部种子萌发形成的幼苗相比具有显著差异(P<0.05)。胀果甘草和乌拉尔甘草植株中下部种子萌发形成的幼苗根冠比相对较高,与植株上部种子萌发形成的幼苗相比具有显著差异(P<0.05)。
2.2.2 对幼苗生长特性的影响 由图3可以看出,3种甘草植株不同部位种子萌发形成的幼苗生长特性不同。光果甘草植株下部种子萌发形成的幼苗株高较高、叶片数较多, 与植株中、上部种子萌发形成的幼苗相比都具有显著差异(P<0.05),而植株中上部种子萌发形成的幼苗根长较长。胀果甘草和乌拉尔甘草是植株上部种子萌发形成的幼苗株高较高、叶片数较多,植株中下部种子萌发形成的幼苗根长较长。

图1 3种甘草植株不同部位种子萌发特性比较(平均值±标准误)Fig.1 Comparisons of the seed germination traits at different positions of three species of Glycyrrhiza (mean±SE) 不同小写字母表示同种类甘草不同部位间的差异显著(P<0.05),下同。Different lowercase letters show significant difference (P<0.05) at different positions of plant in same species of Glycyrrhiza. The same below.

图2 3种甘草植株不同部位种子萌发形成的幼苗总生物量和根冠比的比较(平均值±标准误)Fig.2 Comparisons of the total biomass and the root-shoot ratio of seedlings from different positions seed of three species of Glycyrrhiza (mean±SE)

图3 3种甘草植株不同部位种子萌发形成的幼苗株高、根长和叶片数的比较(平均值±标准误)Fig.3 Comparisons of the plant height, root length and the number of leaves of seedlings from different positions of three species of Glycyrrhiza (mean±SE)
2.3 种子生产的位置效应对幼苗叶绿素含量及叶绿素荧光参数的影响
2.3.1 对叶绿素相对含量的影响 3种甘草植株不同部位种子萌发形成的幼苗叶绿素相对含量(SPAD值)与植株总生物量的相关性分析见图4。光果甘草植株中部(R2=0.61,P<0.01)和上部(R2=0.59,P<0.01)种子萌发形成的幼苗叶绿素相对含量与植株总生物量有极显著的正相关关系,相关性系数也较高(图4A),且植株中上部种子萌发形成的幼苗叶绿素相对含量高于植株下部种子萌发形成的幼苗叶绿素相对含量。胀果甘草幼苗叶绿素相对含量(SPAD值)与植株总生物量的相关性系数很低(图4B),但植株中下部种子萌发形成的幼苗叶绿素相对含量高于植株上部种子萌发形成的幼苗叶绿素相对含量。乌拉尔甘草植株中部(R2=0.53,P<0.01)和下部(R2=0.62,P<0.01)种子萌发形成的幼苗叶绿素相对含量与植株生物量有极显著的正相关关系,相关性系数也较高(图4C),且植株中下部所结种子萌发形成的幼苗叶绿素相对含量高于植株上部种子萌发形成的幼苗叶绿素相对含量(P<0.05)。

图4 3种甘草植株不同部位种子萌发形成的幼苗叶绿素相对含量 (SPAD值)与幼苗生物量的关系Fig.4 The relationship between chlorophyll relative content (SPAD value) and seedling biomass of seedlings at different positions of three species of Glycyrrhiza A: 光果甘草 G. glabra; B: 胀果甘草 G. inflata; C: 乌拉尔甘草 G. uralensis.
2.3.2 对叶绿素荧光参数的影响 由表1数据可知,3种甘草植株不同部位种子萌发形成的幼苗的初始荧光(F0)没有显著差异,光果甘草植株上部种子萌发形成的幼苗最大荧光产量(Fm)和可变荧光(Fv)明显高于植株下部种子萌发形成的幼苗,胀果甘草和乌拉尔甘草植株中下部种子萌发形成的幼苗最大荧光产量(Fm)和可变荧光(Fv)高于植株上部种子萌发形成的幼苗,且有显著差异(P<0.05)。3种甘草幼苗的Fv/Fm值集中在0.80~0.85,Fv/F0值均在4.83~5.22之间,表明幼苗没有受到显著的光胁迫,幼苗PSⅡ系统光能转换效率处于正常水平。光果甘草在植株上部种子萌发形成的幼苗的PSⅡ系统光能转换效率高于植株中下部种子萌发形成的幼苗的PSⅡ系统光能转换效率,无显著差异。胀果甘草和乌拉尔甘草在植株中下部种子萌发形成的幼苗的PSⅡ系统光能转换效率较高,与植株上部种子萌发形成的幼苗的PSⅡ系统光能转换效率相比都具有显著差异(P<0.05)。光果甘草植株上部种子萌发形成的幼苗Fv/F0值高于植株中下部种子萌发形成的幼苗,而胀果甘草和乌拉尔甘草则是植株中下部种子萌发形成的幼苗Fv/F0值高于植株上部种子萌发形成的幼苗,具有显著差异(P<0.05)。说明光果甘草植株上部,胀果甘草和乌拉尔甘草植株中下部种子萌发形成的幼苗PSⅡ潜在的光化学活性高,有活性的PSⅡ反应中心的数量也多,光能转化率也较高。
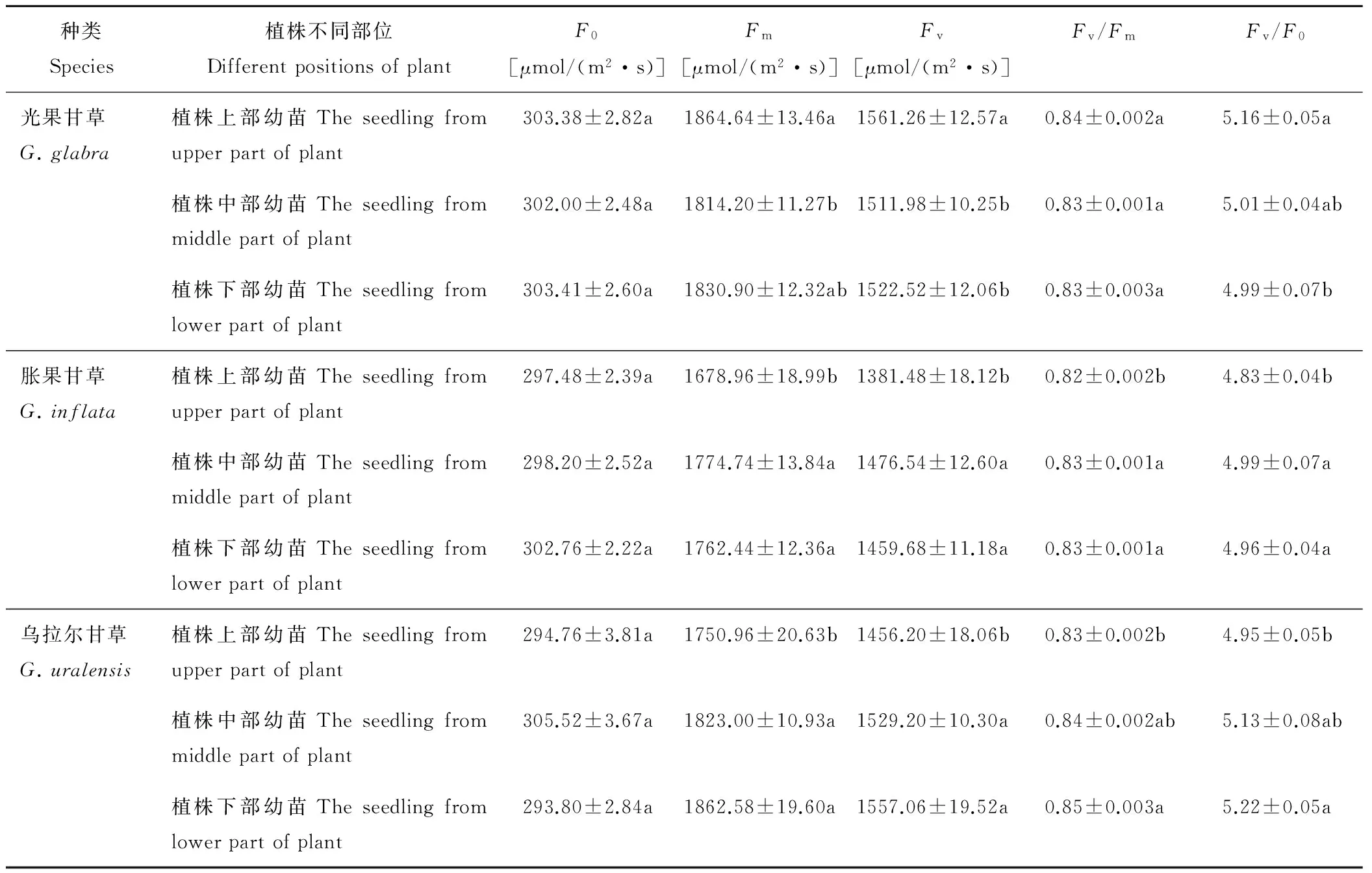
表1 3种甘草植株不同部位种子萌发形成的幼苗叶片叶绿素荧光参数的比较Table 1 Comparisons of the chlorophyll fluorescence parameter of seedlings at different positions of three species of Glycyrrhiza
F0: 初始荧光Initial fluorescence;Fm: 最大荧光Maximal fluorescence;Fv: 可变荧光Variable fluorescence;Fv/Fm: PSⅡ最大光化学效率Maximal photochemical efficiency of PSⅡ;Fv/F0: PSⅡ潜在活性Potential activity of PSⅡ. 同列不同小写字母表示同种甘草植株不同部位在0.05水平上差异显著。Different lowercase letters in the same column indicate the significant differences among different positions of plant at 0.05 level.
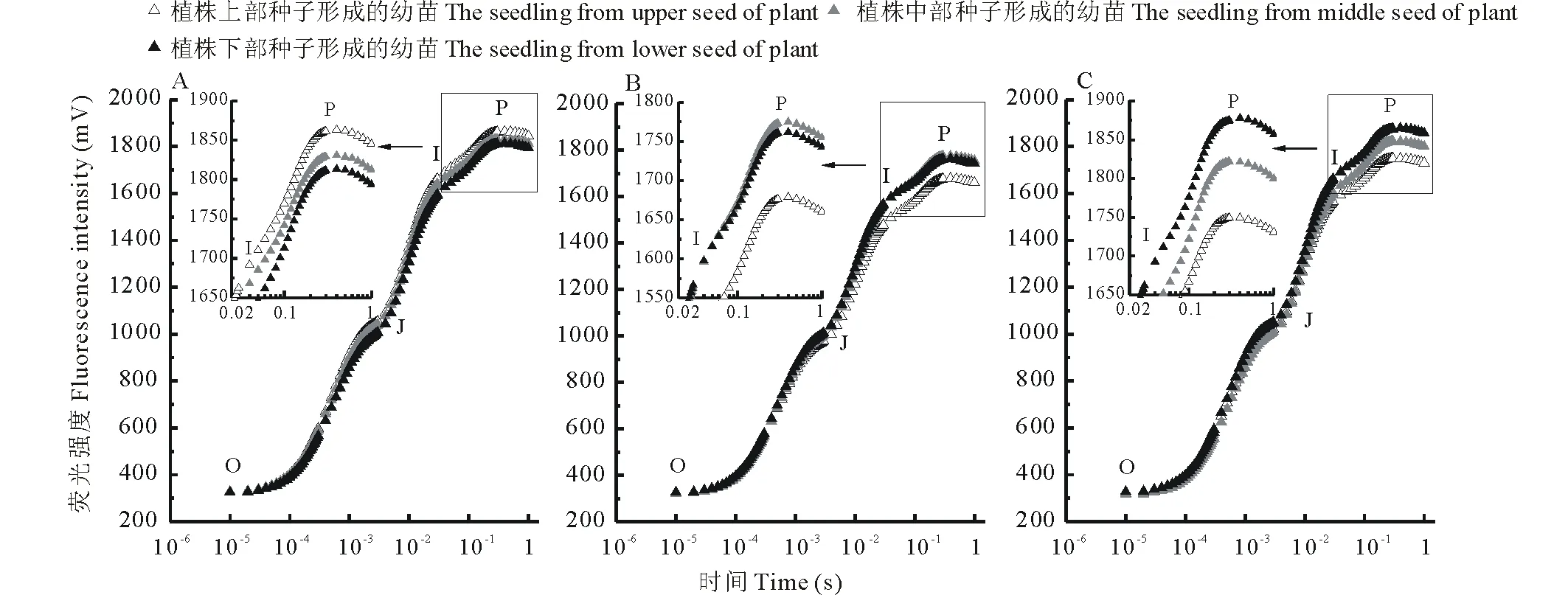
图5 3种甘草不同部位种子萌发形成的幼苗叶片叶绿素荧光诱导动力学曲线的比较Fig.5 The fast chlorophyll fluorescence induction curves of seedlings leaves at different positions of three species of Glycyrrhiza A: 光果甘草 G. glabra; B: 胀果甘草 G. inflata; C: 乌拉尔甘草 G. uralensis.O:初始荧光Initial fluorescence;J:2 ms时的荧光Fluorescence at 2 ms;I:30 ms时的荧光Fluorescence at 30 ms;P:最大荧光Maximal fluorescence.
2.3.3 对叶绿素荧光动力学的影响 3种甘草不同部位种子萌发形成的幼苗叶片叶绿素荧光诱导动力学曲线见图5。由图5可以看出,3种甘草健康功能叶片的叶绿素荧光动力学曲线属于典型的快速叶绿素荧光诱导动力学曲线(O-J-I-P曲线)。相同叶面积的光照条件下,虽到达P点的时间也无差异,但光果甘草植株中上部种子萌发形成的幼苗叶片荧光信号强于植株下部种子萌发形成的幼苗叶片荧光信号(图5A),而胀果甘草(图5B)和乌拉尔甘草(图5C)则是植株中下部种子萌发形成的幼苗的叶片荧光信号强于植株上部种子萌发形成的幼苗叶片荧光信号,尤其表现在I~P相阶段差异明显。
3 讨论与结论
3.1 位置效应对植株不同部位种子萌发及幼苗存活的影响
众所周知,植物自然更新保证了种群的繁衍、延续及群落的组成与稳定,甘草属植物的自然更新主要有种子繁殖和无性繁殖2种方式。然而,在自然环境中野生甘草实生苗却很少见,主要是通过根茎上休眠芽来进行无性生殖(克隆生殖)[12],这种繁殖方式对提高物种的环境适应能力,维持群落的稳定有着极其重要的作用。但是,种子更新能够提高或维持种群的遗传多样性,对种群的扩散和进化十分重要[19]。本研究结果表明,胀果甘草和乌拉尔甘草植株中下部种子具有较高的出苗率,可能对种群扩散的贡献率更大。这与前期的繁殖资源分配模式研究结果一致(胀果甘草和乌拉尔甘草植株中下部具有较高繁殖资源投入,座果率和结实率也较高,且种子品质相对较好),与陈思羽等[20]对大豆(Glycinemax)植株不同冠层种子活力的研究结果也相似。而这2种甘草植株上部种子出苗率都较低,幼苗存活率却与植株中下部种子无差异,其原因可能与种子活力有关。在繁殖资源受限的情况下,为保证子代繁殖成功,在植株上部选择性的败育部分种子,使其余种子具有较高活性,是植物在种子生产过程中对种子数量和质量的权衡。相反,光果甘草却是植株中、上部种子比植株下部种子相比有较高的出苗率、但存活率无差异,认为其原因与繁殖资源分配策略有关。有关研究表明,植物体分配给营养生长和生殖生长的资源与其个体大小有关,植株大小不仅影响繁殖构件的数量和大小,还可影响果实与种子的败育水平以及后代的生存力[21-23]。光果甘草植株较高大,在生殖过程的初期营养生长和生殖生长竞争激烈,虽然在植株下部具有较高的座果率和结籽率,但受资源竞争的影响,植株下部种子储藏的初始能量也较少,种子活性低,出苗率和存活率也较低。而随着植株中、后期的生长,通过败育部分种子,减少种子数量,提高种子活力,缓解了资源竞争的压力,从而提高种子的出苗率和存活率,有利于增加子代的存活机会,以减小繁殖资源限制对植物繁殖成功的影响,提高了子代幼苗的成功建植率。
3.2 位置效应对幼苗生长和建植特性的影响
幼苗通过改变其自身生活史、形态及生理特征,提高环境因子限制时的竞争力和幼苗存活力[24],能使植物适应各种不利的生存环境,对幼苗的散布与拓殖具有重要意义[25]。比如在极端干旱的荒漠环境中,植物通常具有发达的根系,较大的根冠比,可以增强植物的抗旱和抗瘠薄能力[26]。在本研究中,光果甘草植株中上部种子萌发形成的幼苗比植株下部种子萌发形成的幼苗总生物量大,而胀果甘草和乌拉尔甘草植株中下部种子萌发形成的幼苗均比植株上部种子萌发形成的幼苗总生物量大,同时根冠比较高,根也较长。说明这些部位种子萌发后将较多资源分配到地下部分,不仅增大了幼苗对水分和养分的吸收量,增强了幼苗的抗旱和抗瘠薄能力,而且对无性繁殖能力较强的甘草来讲,发达的根系对种群的扩大和延续也具有重要意义[27],这与荒漠植物骆驼刺(Alhagisparsifolia)[26]的生殖策略一致。而出苗率低的种子萌发后将相对较多的资源投入到地上部分的生长,较高的株高和较多的叶片数可以较早地占领空间,抢先获得光源来维持生长,提高建植成功率。由此可见,3种甘草不同部位种子萌发形成的幼苗,采取了不同的生物量分配策略,来提高生存适合度和建植能力。
3.3 位置效应对幼苗叶绿素含量及荧光特性的影响
叶绿素作为光合作用的光敏催化剂,其含量是植物适应和利用环境因子的重要指标[28],能够反映植物适应环境变化所形成的生存对策[29],因此可用叶绿素含量、荧光参数作为判断植物生长和抗逆性强弱的指标[30]。本研究结果表明,光果甘草植株中上部,乌拉尔甘草植株中下部种子萌发形成的幼苗生物量与叶绿素相对含量呈显著的正相关关系,且相关性系数的大小也恰好反映了幼苗生物量的大小。说明光果甘草和乌拉尔甘草植株不同部位种子的差异是影响幼苗叶绿素的合成与生长速率的直接原因。但胀果甘草幼苗植株生物量与叶绿素相对含量无相关性,出现这种现象的原因可能与室内培养条件下光照强度没有达到胀果甘草生长所需,3种甘草中胀果甘草对光照要求最高[12],所以导致胀果甘草幼苗叶绿素荧光动力学曲线P点时叶绿素荧光强度也最弱(1762~1774 mV),低于光果甘草和乌拉尔甘草(1749~1878 mV),说明光照强度影响了叶绿素含量的合成。这也符合3种甘草对野外环境的适应策略,胀果甘草对光照要求较高,使其主要分布于光照充足的区域。乌拉尔甘草对光照要求相对较低,使其成为一个广布种。光果甘草虽光照要求较低,但对水分要求较高,多分布于水多的绿洲[31]。
叶绿素荧光参数的大小和变化往往用于判断和推测植物对环境因子的抗逆性和对环境的适应能力[32-33]。本研究结果表明,光果甘草植株中上部,胀果甘草和乌拉尔甘草植株中下部种子萌发形成的幼苗比植株其他部位种子萌发形成的幼苗具有更高的Fm、Fv、Fv/Fm和Fv/F0,说明这些部位种子萌发形成的幼苗具有更高的PSⅡ电子受体还原能力、荧光产量、光能转换效率和潜在的光化学活性。较高的PSⅡ光化学效率和PSⅡ潜在的光化学活性可增强幼苗对光的利用效率与碳同化能力,同时也增强了其耐盐、耐干旱以及与同类或者其他物种的竞争力[34]。
叶绿素荧光的动力学曲线变化在一定程度上反映了植物对环境的响应[35],通过对3种甘草植株不同部位种子萌发形成的幼苗快速叶绿素荧光诱导动力学曲线的分析,可深入了解植株不同部位种子萌发形成的幼苗对环境的响应。本研究中的幼苗的叶绿素荧光动力学曲线属于典型的OJIP相,说明幼苗没受到胁迫[36]。植物叶片在经一段时间的暗适应后,光系统Ⅱ(PSⅡ)的初级醌受体(QA)、次级醌受体(QB)及质体醌(PQ)库等均完全失去电子而被氧化,此时光系统Ⅱ(PSⅡ)的受体接受电子的能力最大,叶片受光照射后的荧光产量最小,即此时处于初始荧光相“O”点(Fo)。当对植物叶片以强光照射时后,光系统Ⅱ(PSⅡ)反应中心被激发后产生的电子经由光系统传给QA,将其还原生成QA-,而QB不能及时从QA-接受电子将它氧化,造成QA-大量积累,使荧光强度迅速上升至“J”点。此后,QB不断接受来自QA-的电子,形成QB2-,导致QA和光系统完全进入还原状态,此时光系统Ⅱ(PSⅡ)反应中心处于完全关闭状态,不再接受光量子,荧光产量达到最高,使荧光增强到P点[37]。因此,通过对动力学曲线的不同阶段分析和比较,可以推测不同植物的潜在光合能力。本研究结果表明,光果甘草植株中上部,胀果甘草和乌拉尔甘草植株中下部种子萌发形成的幼苗与其他部位种子萌发形成的幼苗相比,无论是在初级醌受体(QA)不断被还原阶段,还是在次级醌受体(QB)不断被还原阶段都具有较强的荧光产量。说明其具有相对较强的电子传递和捕获能力,而且具有较多的反应中心数目和较大的质体醌(PQ)库,利用光能的效率和潜力也高,在相同自然条件下其具有更高的光合能力,使幼苗的生长和建植更具有优势。
本实验结果表明,甘草自然种群的花与果实存在位置效应,并且植株不同部位种子的萌发和幼苗的建植特性不同,但这种差异是否会在以后的生长中消失还有待进一步研究。此外,已有研究表明种子的萌发特征、幼苗的生长和建成过程还与植物物种本身的遗传特性有关[38],不同遗传类型可能会导致生长特性的差异,如花粉来源与子代种子萌发与幼苗生长具有很大相关性[39]。甘草属植物由于开花时序的差异,可能造成植株不同部位的花粉来源不同,这也可能导致种子遗传特性的差异,从而影响种子萌发、幼苗生长和幼苗光合特性。因此,对植株不同结实位置的花粉流进行研究,可为甘草属植物植株不同部位的繁殖策略、种群的适应性进化研究提供更全面的资料。
References:
[1] Hill M J, Luck R. The effect of temperature on germination and seedling growth of temperate perennial pasture legumes. Australian Journal of Agricultural Research, 1991, 42: 175-189.
[2] Kubitzki K, Ziburski A. Seed dispersal in flood plain forest of Amazonia. Biotropica, 1994, 26: 30-43.
[3] Gordon D R, Rice K J. Competitive suppression ofQuercusdouglasii(Fagaceae) seedling emergence and growth. American Journal of Botany, 2000, 87: 986-994.
[4] Paz H, Martinez R M. Seed mass and seedling performance within eight species ofPsychotria(Rubiaceae). Ecology, 2003, 84: 439-450.
[5] Zhang S T, Du G Z, Chen J K. The present situation and prospect of studies on evolutionary ecology of seed size variation. Acta Ecology Sinica, 2003, (2): 353-364. 张世挺, 杜国祯, 陈家宽. 种子大小变异的进化生态学研究现状与展望. 生态学报, 2003, (2): 353-364.
[6] Bu H Y, Chen X L, Xu X L,etal. Seed size and germination in an alpine meadow on the eastern Qinghai-Tibet plateau. Plant Ecology, 2007, 191: 127-149.
[7] Wu G L, Du G Z, Shang Z H. Contribution of seed size and its fate to vegetation renewal: A review. Chinese Journal of Applied Ecology, 2006, 17(10): 1969-1972. 武高林, 杜国祯, 尚占环. 种子大小及其命运对植被更新贡献研究进展. 应用生态学报, 2006, 17(10): 1969-1972.
[8] Wulff R D. Seed size variation inDesmodiumpaniculatum. I. Factors affecting seed size. Journal of Ecology, 1986, 74: 87-97.[9] Buide M L. Pollination ecology ofSileneacutifolia(Caryophyllaceae) floral traits variation and pollinator attraction. Annals of Botany, 2004, 97: 289-297.
[10] Xie T P, Du G Z, Zhang G F,etal. Effects of inflorescence position on seed production and seedling establishment inLigulariavirgaurea. Chinese Journal of Plant Ecology, 2010, 34(4): 418-426. 谢田朋, 杜国祯, 张格非, 等. 黄帚橐吾种子生产的花序位置效应及其对幼苗建植的影响. 植物生态学报, 2010, 34(4): 418-426.
[11] Li X Y. A study of the system and new taxa of genusGlycyrrhizaL. Bulletin of Botanical Research, 1993, 13(1): 13-43. 李学禹. 甘草属分类系统与新分类群的研究. 植物研究, 1993, 13(1): 13-43.
[12] Li X Y, Lu J H. Taxonomy and Experimental Biology of the GenusGlycyrrhizaL.[M]. Shanghai: Fudan University Press, 2015: 15-28. 李学禹, 陆嘉惠. 甘草属(GlycyrrhizaL.)分类系统与实验生物学研究[M]. 上海: 复旦大学出版社, 2015: 15-28.
[13] Chinese Pharmacopoeia Commission. Pharmacopeia of the People’s Republic of China[M]. Beijing: Chemical Industry Press, 2005: 59-60. 国家药典委员会. 中华人民共和国药典(2005年)[M]. 北京: 化学工业出版社, 2005: 59-60.
[14] Tian R W, Lu J H, Xie L B,etal. Effect of flowering mode and pollination on reproductive success and the relationship betweenGlycyrrhizaglabraL. andGlycyrrhizauralensisFisch. Acta Botanica Boreali-Occidentalia Sinica, 2012, 10: 2004-2008. 田润炜, 陆嘉惠, 谢良碧, 等. 光果甘草与乌拉尔甘草开花与传粉方式对生殖及种间关系的影响. 西北植物学报, 2012, 10: 2004-2008.
[15] Fang Y Q. Preliminary Study on Sexual Reproduction of Licorice and Reproductive of the Abortion[D]. Beijing: Beijing University of Chinese Medicine, 2013. 方玉强. 甘草有性生殖过程及生殖败育的初步研究[D]. 北京: 北京中医药大学, 2013.
[16] Wang D, Wang J J. Analyses on seed-setting characteristics and seed yield factors of wildGlycyrrhizauralensis. Journal of Inner Mongolia Agricultural University, 2015, (1): 59-64. 王丹, 王俊杰. 野生乌拉尔甘草结实特性及种子产量影响因子分析. 内蒙古农业大学学报, 2015, (1): 59-64.
[17] Han C, Lv J H, Chen X C,etal. Preliminary study on position effect of inflorescence and seed production, and reproductive resource allocation model of five species inGlycyrrhizaLinn. Journal of Plant Resources and Environment, 2016, 25(3): 72-79. 韩春, 陆嘉惠, 陈晓翠, 等. 5种甘草属植物花序和种子生产的位置效应及繁殖资源分配模式初步研究. 植物资源与环境学报, 2016, 25(3): 72-79.
[18] Lu J H, Lv X, Liang Y C,etal. Salt tolerance ofGlycyrrhizainflataseedlings in Xinjiang and its ion response to salt stress. Chinese Journal of Plant Ecology, 2013, (9): 839-850. 陆嘉惠, 吕新, 梁永超, 等. 新疆胀果甘草幼苗耐盐性及对NaCl胁迫的离子响应. 植物生态学报, 2013, (9): 839-850.
[19] Li X S, Peng M C, Dang C L. Research progress on natural regeneration of plants. Chinese Journal of Ecology, 2007, (12): 2081-2088. 李小双, 彭明春, 党承林. 植物自然更新研究进展. 生态学杂志, 2007, (12): 2081-2088.
[20] Chen S Y, Liu P, Zhu M,etal. Seed vigor and antioxidant enzyme activities during germination in different canopies of soybean. Chinese Bulletin of Botany, 2016, (51): 24-30. 陈思羽, 刘鹏, 朱末, 等. 大豆植株不同冠层种子活力及其萌发中抗氧化酶活性. 植物学报, 2016, (51): 24-30.
[21] Susko D J, Lovett-Doust L. Plant-size and fruit-position effects on reproductive allocation inAlliariapetiolata(Brassicaceae). Canadian Journal of Botany, 2000, 78: 1398-1407.
[22] Liu Z J, Du G Z, Chen J K. Size-dependent reproductive allocation ofLigulariavirgaureain different habitats. Chinese Journal of Plant Ecology, 2002, (1): 44-50. 刘左军, 杜国祯, 陈家宽. 不同生境下黄帚橐吾(Ligulariavirgaurea)个体大小依赖的繁殖分配. 植物生态学报, 2002, (1): 44-50.
[23] Han B, Wang X F, Huang S Q. Production of male flowers does not decrease with plant size in insect-pollinatedSagittariatrifolia, contrary to predictions of size-dependent sex allocation. Journal of Systematics and Evolution, 2011, 49: 379-386.
[24] Zhang G F. Studies on Reproductive Trait and Seedling Recruitment of Typical Clonal PlantLigulariavirgaureain Qinghai-Tibet Plateau[D]. Lanzhou: Lanzhou University, 2012. 张格非. 青藏高原典型克隆植物黄帚橐吾的繁殖特征与幼苗更新研究[D]. 兰州: 兰州大学, 2012.
[25] Lu Y, Huang S Q. Adaptive advantages of gynomonoecious species. Acta Physiologica Sinica, 2006, 44(2): 231-239.
[26] Zeng F J, Liu B, He J X,etal. Effect of irrigation on nitrogen characteristics ofAlhagisparsifoliaseedlings. Acta Ecology Sinica, 2010, (8): 2118-2127. 曾凡江, 刘波, 贺俊霞, 等. 骆驼刺幼苗氮素特征对不同灌溉量的响应. 生态学报, 2010, (8): 2118-2127.
[27] Wang S F, Hu Y X, Sun H Q,etal. Effects of salt stress on growth and root development of two oak seedlings. Acta Ecology Sinica, 2014, 34(4): 1021-1029. 王树凤, 胡韵雪, 孙海菁, 等. 盐胁迫对2种栎树苗期生长和根系生长发育的影响. 生态学报, 2014, 34(4): 1021-1029.
[28] Liu Y Q, Sun X Y, Wang Y,etal. Effects of shades on the photosynthetic characteristics and chlorophyll fluorescence parameters of Urticadioica. Acta Ecologica Sinica, 2007, 27(8): 3457-3464.
[29] Xu L, Gao Z Q, An W,etal. Flag leaf photosynthetic characteristics, change in chlorophyll fluorescence parameters, and their relationships with yield of winter wheat sowed in spring. Chinese Journal of Applied Ecology, 2016, (1): 133-142. 徐澜, 高志强, 安伟, 等. 冬麦春播条件下旗叶光合特性、叶绿素荧光参数变化及其与产量的关系. 应用生态学报, 2016, (1): 133-142.
[30] Ye B, Wu Y B, Shao W,etal. Effects of combined stress of elevated temperature and drought and of re-watering on the photosynthetic characteristics and chlorophyll fluorescence parameters ofBroussonetiapapyriferaseedlings. Chinese Journal of Ecology, 2014, 33(9): 2343-2349. 叶波, 吴永波, 邵维, 等. 高温干旱复合胁迫及复水对构树(Broussonetiapapyrifera)幼苗光合特性和叶绿素荧光参数的影响. 生态学杂志, 2014, 33(9): 2343-2349.
[31] Li X B, Chen L, Li G Q,etal. Ecological distribution and propagative technique research ofGlycyrrhizaresourcesin China. Ecology and Environmental Sciences, 2013, 22(4): 718-722. 李学斌, 陈林, 李国旗, 等. 中国甘草资源的生态分布及其繁殖技术研究. 生态环境学报, 2013, 22(4): 718-722.
[32] Qiu N W, Zhou F, Gu Z J,etal. Photosynthetic functions and chlorophyll fast fluorescence characteristics of fivePinusspecies. Chinese Journal of Applied Ecology, 2012, 23(5): 1181-1187. 邱念伟, 周峰, 顾祝军, 等. 5种松属树种光合功能及叶绿素快相荧光动力学特征比较. 应用生态学报, 2012, 23(5): 1181-1187.
[33] Chen C, He X D, Qin J Z,etal. Comparison of chlorophyll fluorescenceFv/Fmcharacteristics of four michelia trees. Journal of Anhui Agricultural University, 2013, 40(1): 32-37. 陈辰, 何小定, 秦金舟, 等. 4种含笑叶片叶绿素荧光参数Fv/Fm特性的比较. 安徽农业大学学报, 2013, 40(1): 32-37.
[34] Feng Q, Hu D, Li N. Comparative analysis of leaf gas exchange and chlorophyll fluorescence ofEuonymusjaponicasThunb by field experiment in the typical urban and suburban areas in Beijing. Acta Ecology Sinica, 2009, (7): 3477-3484. 冯强, 胡聃, 李娜. 典型城区与郊区环境大叶黄杨气体交换及叶绿素荧光特性比较. 生态学报, 2009, (7): 3477-3484.
[35] Jiang C D, Gao H Y, Zou Q. Changes of donor and accepter side in photosystem II complex induced by iron deficiency in attached soybean and maize leaves. Photosynthetica, 2003, 41: 267-271.
[36] Qiu N W, Zhou F, Wang Y,etal. Comparison on characteristics of the fast chlorophyll fluorescence induction kinetics betweenPinusspeciesandPopulusspecies. Scientia Silvae Sinicae, 2013, (3): 136-143. 邱念伟, 周峰, 王颖, 等. 松树与杨树叶片叶绿素快相荧光动力学特征比较. 林业科学, 2013, (3): 136-143.
[37] Li P M, Gao H Y, Reto J,etal. Application of the fast chlorophyll fluorescence induction dynamics analysis in photosynthesis study. Journal of Plant Physiology and Molecular Biology, 2005, (6): 559-566. 李鹏民, 高辉远, Reto J, 等. 快速叶绿素荧光诱导动力学分析在光合作用研究中的应用. 植物生理与分子生物学学报, 2005, (6): 559-566.
[38] Moles A T, Westoby M. What do seedling die from, and what are the implications for evolution of seed size. Oikos, 2004, 106: 193-199.
[39] Ou J L, Guo J J, Zhao Z G,etal. Preliminary report on offspring fitness in relation to patch size and genetic diversity ofBetulaalnoidesin a heterogenous landscape. Seed, 2012, (5): 15-18, 22. 欧景莉, 郭俊杰, 赵志刚, 等. 西南桦自由授粉子代适应性与斑块大小、遗传多样性间的相关性. 种子, 2012, (5): 15-18, 22.
Effects of seed position on seedling establishment in three medicinal licorice species
HAN Chun1, LU Jia-Hui1,2,3*, ZHOU Ling-Ling1, NIU Qing-Dong1, SONG Feng1, WANG Yue1, YANG Xu1
1.CollegeofLifeSciencesofShiheziUniversity,Shihezi832003,China; 2.TheKeyLaboratoryOasisEco-agriculture,XinjiangProductionandConstructionGroup,Shihezi832003,China; 3.InstituteofLicoriceofShiheziUniversity,Shihezi832003,China
The position effect of seeds within a plant is a common phenomenon, but its influence on seed germination and seedling establishment is not well known. The objective of this study was to determine whether there were differences in seed germination strategies and seedling growth among seeds obtained from different (upper, middle, and lower) positions of the plant for three medicinal licorice species (Glycyrrhizaglabra,Glycyrrhizainflata,Glycyrrhizauralensis).Seeds were collected from different positions on the plant, and then seed germination, seedling growth, seedling chlorophyll content, and chlorophyll fluorescence were measured. The main findings can be summarized as follows: 1) InG.glabra, the seedling emergence rates were higher for seeds from upper and middle positions than for seeds from the lower position. ForG.inflateandG.uralensis, the seedling emergence rates were higher for seeds from middle and lower positions than for those from the upper position. The seedling survival rate did not differ significantly among the different collection positions. 2) The seedlings from the seeds with a high seedling emergence rate had higher total biomass, root length, and root∶shoot ratio than those of other seedlings, and a larger proportion of their biomass was allocated to below-ground parts, which can enhance seedling resistance and environmental adaptability. The seedling chlorophyll content and chlorophyll fluorescence parameters (Fv,Fm,Fv/Fm,Fv/F0, electron transmission speed in the light reaction) were also higher in seedlings from seeds with a high seedling emergence rate. In addition, the fluorescence kinetics curve showed that these seedlings had a strong fluorescence intensity and high electron and light capture rates, which can enhance seedling establishment. In contrast, the seedlings from seeds with a low seedling emergence rate had greater plant height and more leaves to occupy space and intercept light. These results indicate that the seeds from different positions on the plant have different germination and seedling growth strategies, which are of great ecological significance for seedling establishment and population regeneration of wild licorice plants.
medicinal licorice; seed; positional effect; seedling establishment
10.11686/cyxb2016344
2016-09-14;改回日期:2016-11-14
国家自然科学基金(31260042)资助。
韩春(1991-),男,甘肃武威人,在读硕士。E-mail:wwhanchun@163.com
*通信作者Corresponding author. E-mail:jiahuil@shzu.edu.cn
http://cyxb.lzu.edu.cn
韩春, 陆嘉惠, 周玲玲, 牛清东, 宋凤, 王跃, 杨旭. 三种药用甘草种子位置效应对幼苗建植特性的影响. 草业学报, 2017, 26(7): 128-137.
HAN Chun, LU Jia-Hui, ZHOU Ling-Ling, NIU Qing-Dong, SONG Feng, WANG Yue, YANG Xu. Effects of seed position on seedling establishment in three medicinal licorice species. Acta Prataculturae Sinica, 2017, 26(7): 128-137.
