新疆达坂城盐湖不同植物群落土壤呼吸研究①
2017-07-21李典鹏姚美思韩东亮王丽萍胡保安王宁宁蒋大勇朱新萍贾宏涛
李典鹏,姚美思,韩东亮,王丽萍,胡保安,王宁宁,蒋大勇,张 凯,朱新萍,贾宏涛
新疆达坂城盐湖不同植物群落土壤呼吸研究①
李典鹏,姚美思,韩东亮,王丽萍,胡保安,王宁宁,蒋大勇,张 凯,朱新萍,贾宏涛*
(新疆农业大学草业与环境科学学院,乌鲁木齐 830052)
新疆盐湖分布地域广泛而不均匀。盐湖生态系统碳循环是干旱区生态系统碳循环的重要组分,其动态变化对干旱区碳平衡有着重要的影响,然而目前对干旱区盐湖土壤呼吸的研究相对薄弱。为探讨盐湖沿岸不同植物群落土壤呼吸及其影响因子,以新疆达坂城盐湖为典型研究区,选取沿岸小獐毛()、鸢尾()、芨芨草()、黑果枸杞()植物群落土壤为主要研究对象,以周边农田撂荒地为对照,利用LI-COR 8100对达坂城盐湖沿岸4种植物群落和撂荒地土壤呼吸进行了监测,结果表明:盐湖沿岸土壤呼吸具有明显的日变化。监测时段内土壤呼吸作用均为单峰曲线,排放通量在13:00左右最高,07:00左右出现最低值。不同群落土壤呼吸速率变化范围在0.89 ~ 4.34 µmol/(m2·s)之间,撂荒地为1.77 ~ 3.48 µmol/(m2·s),表现为黑果枸杞>小獐毛>芨芨草>撂荒地>鸢尾。不同植物群落地下生物量影响土壤CO2排放,研究区土壤呼吸日平均累计CO2排放通量为209.28 mg/(m2·d)。土壤呼吸速率与土壤5 cm温度呈现显著正相关(<0.01),可以解释土壤呼吸日变化的69% ~ 78%,是决定达坂城盐湖土壤呼吸变化的主要因子。利用土壤5 cm处温度得到小獐毛、鸢尾、芨芨草、黑果枸杞、撂荒地的10值分别为2.30、1.24、1.20、1.57、1.28,小獐毛样地土壤呼吸对温度更敏感。除撂荒地外各群落土壤呼吸与土壤湿度呈显著负相关(2= 0.38 ~ 0.51)。该研究对进一步明确干旱区盐湖生态系统土壤呼吸变化规律和影响因素,以及对估算区域碳平衡及“碳汇”功能具有重要意义。
干旱区;盐湖;碳循环;土壤呼吸;影响因子
呼吸作用是生物能学的一个重要组成部分,由于土壤生物过程十分复杂,故现阶段的土壤呼吸主要集中在研究土壤CO2释放量[1]。土壤呼吸作为全球生态系统碳循环的重要环节,也是全球气候变化的关键生态过程。不同时间尺度上探讨影响土壤呼吸特征及其敏感环境因素,对准确评估生态系统碳收支有着重要的意义。作为温室气体的主要组成部分,CO2在限制地表红外辐射向外传送起到重要的作用,更多的热量被存储在了大气中,从而导致气候变暖。特定条件下土壤作为“碳汇”,对缓解区域气候变化起到了一定的作用;据估计,在全球尺度上,土壤每年向大气中排放77 Pg C[2],土壤呼吸作为“碳源”释放碳的速率比人类活动释放碳的速率要大一个数量级。因此,在气候变暖与全球碳循环之间的正反馈中,土壤呼吸是一个关键的过程。为了正确评价气候变化中的不确定性,需要了解不同类型土壤及特定条件下的呼吸特征及其影响因素。
盐湖是湿地生态系统的重要组成部分,具有盐分和水分双重胁迫的特点,干旱区盐湖湿地土壤呼吸的研究相对薄弱。在极端干旱的盐生荒漠系统,盐碱土对大气中的CO2有很好的吸收能力,吸收CO2量可达62 ~ 622 g/(m2·a)[3-4],其吸收量相当于植物的2 ~ 15倍,且湿润年份日累积最大CO2吸收量是干旱年份的4倍[5]。在一定程度上说明水分能够影响到盐碱土的“碳汇”功能,盐湖作为干旱区特殊的湿地,无论是从潮湿土壤有机固碳还是盐渍化土壤无机固碳的角度来看,均应该是典型“碳汇”。但其中的核心问题取决于盐湖周边土壤的呼吸,即土壤CO2排放特征。
通过对盐湖周边土壤呼吸的研究,明确驱动盐湖土壤呼吸的关键因素,可为湿地生态系统在全球气候变化中作用的准确评估提供参考数据。本研究拟以新疆达坂城盐湖为典型研究区域,以其周边生长不同主要植物群落立地土壤为研究对象,对土壤呼吸速率及其影响因子进行原位监测,明确达坂城盐湖沿岸不同植物群落土壤呼吸速率日变化特征,为进一步明确内陆盐湖湿地在干旱区陆地碳循环中的地位提供科学依据。
1 材料与方法
1.1 研究区概况
达坂城盐湖又名盐湖、破城子湖、乌鲁木齐湖等[6],位于新疆维吾尔自治区乌鲁木齐市区东南45 km达坂城区西部(88°03′53″ ~ 88°12′15″E,43°21′00″ ~43°25′25″N)(图1)。湖盆为封闭内流盆地,周围无常年性地表河流,靠大气降水和地下水补给,面积35 km2,湖面海拔1 066 m。湖盆在柴窝堡山间盆地东南部,南北高山夹持,气候干旱,年平均气温 6.1℃(月平均气温–22.1℃,7月平均气温 35.8℃,年较差 31.1℃),日较差12.1℃;年降水量71.8 mm,年蒸发量2 648 mm,相对湿度38%;冰冻期5个月,年平均降(积)雪150 d,最大积雪深度30 ~ 40 cm;年日照数2 500 ~ 2 600 h,全年多刮西北风,最大风力10 级,最大风速30 m/s,属于干旱大陆性气候。盐湖沿岸主要植被类型为小獐毛()、鸢尾()、芨芨草()、黑果枸杞()、苔草()等。
1.2 研究方案
试验于2015年7—9月进行,以盐湖为核心,根据试验区植物群落特点,由湖心向外依次选取小獐毛()、鸢尾()、芨芨草()、黑果枸杞()群落和撂荒地(Abandoned land)土壤作为监测对象,研究样地基本性质见表1。在每个群落内随机选取3块样地,每块样地10 m × 10 m,沿样方对角线设置2个长期固定的底座,每月中下旬选择2 ~ 3个相对晴朗的天气进行土壤呼吸的监测,从上午07:00至晚上19:00,每2小时监测1次,监测土壤呼吸的同时监测环境因素指标,包括近地面气温,土壤5、10、15、20和25 cm处土温,表土层含水量(0 ~ 10 cm)、气压、风速、光照强度等。
1.3 研究方法
1.3.1 土壤呼吸测定 利用土壤碳通量测量系统LI-COR 8100(Li-COR,Lincoln,NE,USA)测定土壤呼吸速率。每块样地内设两个内径为20 cm、高10 cm 的PVC管基座,将基座嵌入土中,使上端高出地表约5 cm,剪除基座环内的植物。为减少由于嵌入基座对土壤扰动而造成短期内土壤呼吸速率的波动,经过24 h 的平衡后,土壤呼吸速率恢复到正常水平才进行相关指标的测定。监测时每个基座测定1 次,即每个处理6个监测数据。
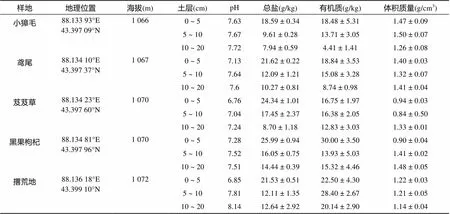
表1 达坂城盐湖研究样地概况
1.3.2 土壤温度、湿度测定 与土壤呼吸监测同步,监测土壤湿度和土壤温度;土壤湿度采用野外采样-室内烘干法进行,以基座为圆心在半径为1 m的圆形范围内均为土壤湿度的采样范围;5 cm土壤温度采用LI-COR 8100 自带的温度探头,监测前一天在PVC管基座附近安置地温计,同步对10、15、20、25 cm处土壤地温进行监测。
1.3.3 植被调查方法 植被调查采用常规方法,由于不同植物群落的差异,所以调查方法也略有不同。小獐毛群落和撂荒地植被调查方法如下:在所选样地附近,按一定方向设置50 m 样线,每隔10 m 布设一个1 m×1 m 的样方。鸢尾、芨芨草、黑果枸杞群落生物量测定,按一定方向设置50 m 样线,则为每隔10 m 布设一个5 m×5 m 的样方,对样方内地表活体植物和凋落物进行采集和分析,估计样方内植被密度、盖度,用称重法测定生物量。根系样品采集采用根钻法,选择剪完草的5个样方,采取0 ~ 5、5 ~ 10、10 ~ 20、20 ~ 30、30 ~ 50、50 ~ 70、70 ~ 100 cm 分层根系样品。
1.3.4 数据处理 采用单因素方差分析(ANOVA)检验不同群落土壤呼吸速率的显著性。对所得数据采用 Excel 2016、SPSS 20.0 进行数据处理分析和制图,绘图工具为MapInfo 8.5和Adobe Photoshop CS5。采用 Van’t Hoff 模型分析土壤温度与土壤呼吸速率的关系:
= aebT(1)
式中:表示土壤呼吸(µmol/(m2·s),下同),表示土壤温度(℃,下同),a、b均为参数(下同)。
采用线性回归分析的方法分析土壤湿度与土壤呼吸速率的关系:
= a+ c (2)
式中:表示土壤湿度(%),c为参数(下同)。
采用多性回归分析法分析土壤水分和温度与土壤呼吸速率的关系:
= a+ b+ c (3)
采用公式(4)计算土壤呼吸对温度变化敏感程度:
10= e10b(4)
式中:10为土壤呼吸的温度敏感性,即土壤温度升高10℃时土壤呼吸速率增加或减少的倍数;b为Van’t Hoff 指数模型 (= aebT) 中的温度反应常数。
2 结果与分析
2.1 不同植被群落土壤呼吸日动态
图2为达坂城盐湖不同植被群落土壤呼吸速率的日动态,可以看出,各土壤呼吸速率的日变化均为先增后降的单峰曲线。不同植被类型土壤呼吸的日变化特征存在差异,大小为黑果枸杞>小獐毛>芨芨草>鸢尾,其中鸢尾群落土壤呼吸速率明显低于其他群落。曲线的最高峰值均出现在13:00左右,小獐毛、鸢尾、芨芨草、黑果枸杞和撂荒地曲线峰值分别为4.02、2.26、3.37、4.94、3.48 µmol/(m2·s)。除撂荒地外,各个群落土壤呼吸速率最低值出现在19:00;撂荒地土壤呼吸速率在17:00时降低到2.05 µmol/(m2·s),随后其呼吸速率又呈现增加趋势。
2.2 植被生物量对土壤CO2排放通量的影响
从图3可以看出,小獐毛、鸢尾、芨芨草、黑果枸杞群落和撂荒地地上活体生物量分别为144.56、32.70、55.21、51.47、21.9 g/m2,小獐毛群落土壤地上活体生物量与其他4个处理差异达到极显著水平(<0.01)。小獐毛、鸢尾、芨芨草、黑果枸杞和撂荒地地表凋落物生物量分别为84.81、14.84、33.36、32.13、9.43 g/m2,小獐毛群落土壤地表凋落物与其他群落的凋落物差异达显著水平(<0.05);鸢尾与芨芨草、芨芨草与黑果枸杞地表凋落物差异性未达到显著水平(>0.05)。小獐毛、鸢尾、芨芨草、黑果枸杞和撂荒地土壤CO2排放总量分别为5.35、3.55、5.16、6.23、4.84 g/(m2·d);各群落和撂荒地之间土壤CO2日平均通量的差异性达到极显著水平(<0.01)。总体来看,活体生物量大、凋落物高的小獐毛群落土壤,其CO2排放通量大于生物量小、凋落物低的鸢尾、芨芨草群落和撂荒地,低于黑果枸杞群落。
本文利用土壤碳通量测量系统LI-COR 8100测定土壤呼吸,而土壤呼吸包括了微生物呼吸和根呼吸以及无机过程,微生物呼吸与土壤中微生物数量密切相关;根呼吸与植物地下根系生物量有关;在盐生荒漠系统中土壤温度的变化改变了CO2在土壤溶液中的溶解度,催生出一个日出夜进的“无机呼吸”过程,此过程与环境温度的变化关系密切。通过分析不同植物群落土壤0 ~ 100 cm 根系与土壤CO2日平均通量的关系,结果表明不同植物群落土壤根系生物量差异性达到极显著水平,且各群落土壤CO2排放通量也存在显著性差异,通过对根系生物量与CO2通量拟合的结果表明,土壤呼吸与根系生物量呈线性相关:= 0.111 7+ 3.385 8(² = 0.64,<0.05),说明土壤呼吸也受地下根系生物量的影响。
2.3 土壤呼吸与土壤温度、湿度的关系
在监测时段内,各群落和撂荒地土壤5 cm温度呈现出先增加后下降的趋势(图4),但不同群落土壤表层温度峰值出现的时间存在差异,小獐毛、鸢尾、芨芨草群落土壤峰值出现在17:00,分别为32.3℃、39.2℃、42.3℃,撂荒地的峰值出现在15:00,为41.4℃,黑果枸杞群落土壤最高温度出现在13:00,为40.8℃。除小獐毛群落10 cm土壤温度日变化为单增曲线之外,其他监测点10 cm土壤温度变化均为单峰曲线;小獐毛群落10 cm土壤温度最高值较表层峰值滞后1 h,为27.6℃。鸢尾、芨芨草群落和撂荒地10 cm土壤温度峰值与土壤表层峰值出现时刻一致,黑果枸杞群落10 cm土壤温度最高值较土壤表层温度峰值滞后4 h出现。且各群落土壤10 cm以下土层温度变幅不大。
通过 Van’t Hoff 指数方程分别拟合土壤呼吸速率与土壤5、10、15、20、25 cm 温度的回归关系,并用决定系数(2) 来解释土壤呼吸速率的变异量,结果表明土壤呼吸速率与5 cm地温的拟合关系最为理想,即Van’t Hoff指数方程能很好地描述土壤呼吸对土壤5 cm 温度的响应(表2)。不同群落及撂荒地土壤5 cm温度与土壤呼吸速率具有指数关系(图4),在> 99% 的置信区间内,土壤5 cm温度的变化能够解释土壤呼吸变化的69% ~ 78%,而10 cm 土层土壤温度的变化可以解释土壤呼吸变化的24% ~ 44%。然而,在对15、20、25 cm土层温度与土壤呼吸速率进行拟合时,三者决定系数与5、10 cm决定系数相比有明显减弱,因此15 cm 以及15 cm 以下土层温度对土壤呼吸的贡献率可以不用考虑。5 cm 土层土壤温度对土壤呼吸速率的决定系数高于10 cm土层,说明在盐湖沿岸生态系统中,5 cm 土层土壤温度可以作为影响土壤呼吸急剧变化的变量。
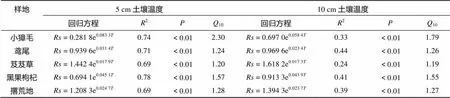
表2 土壤呼吸的温度敏感系数及土壤呼吸日变化与土壤温度指数回归关系
土壤呼吸温度敏感系数(10)大小顺序依次为小獐毛群落>黑枸杞群落>撂荒地>鸢尾群落>芨芨草群落(表2),小獐毛群落土壤的10值为2.30,明显高于其他4个处理,说明小獐毛群落土壤呼吸对温度变化的响应程度大于其他群落土壤。但是土壤呼吸和土壤温度、土壤湿度等生物环境因素有着多重关系,所以如果仅用10解释盐湖不同群落和撂荒地土壤呼吸的变化规律具有一定的不确定性。
各群落土壤湿度与土壤呼吸的关系如图6所示。不同群落土壤呼吸与土壤湿度进行拟合分析可知(表3),各群落土壤湿度与土壤呼吸速率呈显著负相关,土壤呼吸曲线决定系数在0.38 ~ 0.51之间;撂荒地土壤湿度与土壤呼吸速率呈显著正相关(2= 0.38)。各群落和撂荒地土壤湿度和土壤呼吸之间线性关系达到显著水平,说明土壤湿度也是影响土壤呼吸的因子。4种植物群落土壤湿度与土壤呼吸相关性决定系数大小顺序为鸢尾>小獐毛>芨芨草>黑果枸杞,决定系数的大小在一定程度上反映不同群落土壤呼吸速率对水分的依赖程度,出现此现象的原因可能与植物群落与湖边的距离有关,距离不同,水分的胁迫程度不同。
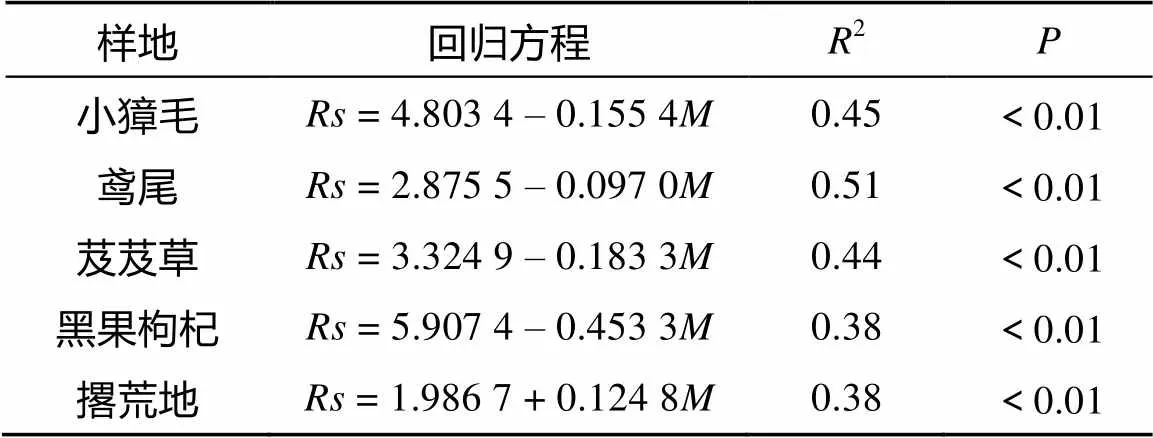
表3 土壤呼吸与土壤湿度的线性回归关系
2.4 土壤呼吸与土壤温度及土壤湿度的线性回归分析
不同群落土壤呼吸与土壤表层温度、湿度的线性回归分析见表4。各群落和撂荒地土壤呼吸与温度的相关系数分别为0.85、0.84、0.73、0.77、0.76,相关性显著。在各群落和撂荒地土壤回归方程中,土壤湿度和表层温度对土壤呼吸的影响均存在较小的交互作用,其中芨芨草群落土壤湿度与表层温度对土壤呼吸的交互作用大于小獐毛和鸢尾群落,在回归方程中,土壤湿度和表层温度均进入回归方程(95% 置信区间),以温度-水分-土壤呼吸双因子模型可以解释土壤呼吸的变化。从各群落土壤呼吸与湿度、温度的相关系数可以看出,沿湖心向外土壤呼吸对温度的响应呈先降低再增加的变化趋势;不同群落土壤呼吸与土壤湿度呈显著的负相关,由芨芨草群落到小獐毛群落土壤呼吸和含水量的负相关依次增强,即鸢尾>小獐毛>芨芨草>黑果枸杞。撂荒地土壤呼吸与土壤温度、湿度呈正相关。
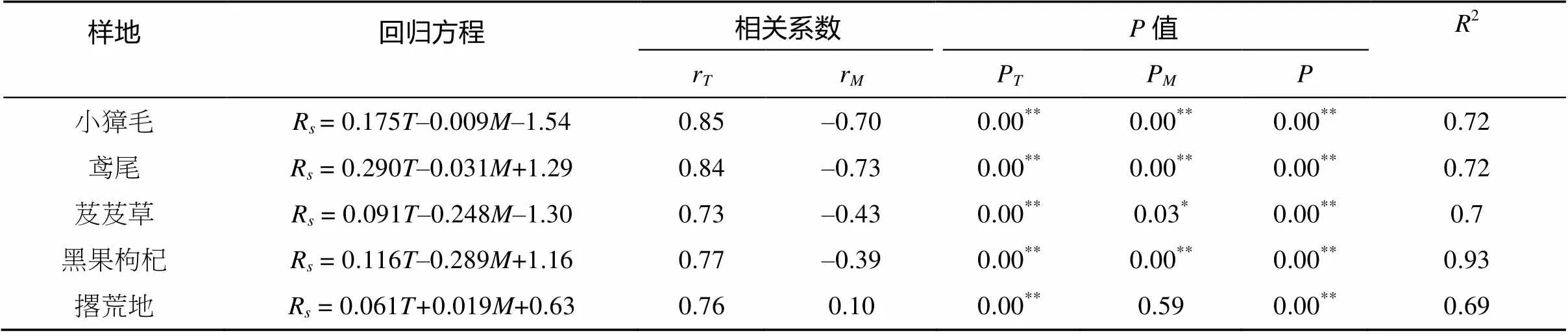
表4 土壤呼吸与土壤温度和土壤湿度的函数模拟
注:R表示土壤呼吸速率,表示温度,表示土壤湿度;** 表示达到<0.01显著水平,* 表示达到<0.05显著水平。
3 讨论
3.1 不同植物群落土壤呼吸日变化特征
盐湖生态系统不同群落条件下土壤呼吸日变化呈现明显单峰曲线,此结果与国内外大部分学者研究相同[7-9],峰值均出现在13:00,这可能与土壤温度和湿度有关,凌晨土壤温度较低,且土壤湿度较高,二者同时限制土壤呼吸,导致土壤呼吸较弱。随着气温的上升,土壤温度与土壤湿度也随之变化,同时也促使土壤呼吸速率的增加,当土壤温度和土壤湿度在13:00达到适宜条件时,土壤酶活性较高,促进了植物根系和微生物的代谢速率[10-11],故土壤呼吸在此时出现监测时段内的峰值。13:00后土壤温度持续上升,由于温度升高和蒸发速率加强导致土壤湿度下降,湿度与温度交互作用从而使土壤呼吸减弱,故13:00后土壤呼吸速率呈下降趋势。在干旱区盐生荒漠,马杰等[12]对原始盐生荒漠多枝柽柳群落土壤CO2通量观测表明,盐生荒漠土壤CO2排放通量呈现出单峰的日变化过程。相关研究结果表明,土壤呼吸日变化相对于土壤温度的变化存在不定的提前效应[10]或滞后[13]现象。在本研究中,土壤呼吸速率最大值出现在13:00,相对于土壤表层温度最高值出现提前2 h,这与Jassal等[14]的研究结果一致,其原因可能是土壤温度和含水量的变化所导致,除此之外底物供应也可能是调节土壤呼吸日变化的一个重要因子[15]。
3.2 植物类型对土壤呼吸的影响
植物是生态系统的重要组成部分,通过植物的相关指标可以间接地反映生态系统的稳定性和环境状况。植物类型和生物量的大小是影响土壤和大气之间进行CO2交换的因素,不同群落有不同植物类型和覆盖度,凋落物、生物量的数量方面也有较大差异,这些都有可能对土壤的呼吸速率造成一定的影响。通过对盐湖生态系统的研究发现,建群种分别为小獐毛、鸢尾、芨芨草、黑果枸杞,单位面积的生物量沿湖心向外有逐步递减的趋势,小獐毛样地地上生物量与鸢尾、芨芨草地上生物量存在显著性差异,且CO2通量也存在显著性差异。黑果枸杞地上生物量和凋落物均小于小獐毛,但其呼吸速率大于其他3个群落和撂荒地。地表凋落物分解速率也是影响土壤与大气之间CO2交换速率的因素之一,本研究表明,除撂荒地外,其他群落都呈现土壤呼吸速率随凋落物增加而增加的趋势。董自红等[16]、王忆慧等[17]通过对高山草甸草原植物群落的研究结果表明土壤呼吸与凋落物存在线性正相关,本研究结果与此相同。与同处于干旱区的艾比湖相比,达坂城盐湖沿岸不同群落土壤呼吸速率要高于艾比湖流域[18],究其原因,是因为艾比湖流域植被类型主要以胡杨、梭梭等木本植物为主,植物覆盖度低、植物残体较少,导致土壤微生物活动较弱、有机质的矿化速率缓慢,从而降低了土壤CO2的排放。不同土地利用方式对土壤呼吸也有不同影响,达坂城盐湖沿岸土壤呼吸速率与橡胶林土壤相近[19],低于种植玉米农田土壤[20]。在不考虑环境条件的情况下,此类结果均有可能是由于不同植被类型所导致。
3.3 环境因素对土壤呼吸的影响
从本质上来说,土壤呼吸是一个生物化学过程,土壤温度、湿度几乎影响土壤呼吸的各个方面。在生物化学水平,温度通过影响酶活性从而影响根和根际微生物的呼吸速率;在物理水平,温度和水分相互作用影响土壤孔隙结构与外界的气体交换。就大部分情况而言,土壤表层及亚表层温度在时间尺度上变化较为明显,通过分析可知土壤温度函数都可以用于解释土壤呼吸的动态变化。微生物呼吸作为土壤CO2排放的来源之一,其释放速率受温度的影响较大,并且与土壤温度有着密切的联系;温度通过影响土壤中各类酶活性、微生物代谢速率、土壤动物呼吸速率和植物根系生长从而影响土壤CO2的排放[21-22]。土壤呼吸与土壤温度的拟合方式有幂函数、线性、抛物线和指数函数等,不同研究结果拟合模型也有所不同,但指数函数拟合的方法较多[23],原因可能与温度敏感系数的计算有关。本研究中土壤呼吸与土壤温度呈显著的指数相关(2分别为0.74、0.71、0.69、0.78、0.69;<0.01),且土壤呼吸与土壤5 cm温度的指数拟合相关性大于10 cm,这与陈宝玉等[24]研究结果不一致,这可能是由于盐湖沿岸土壤5 cm温度在时间尺度上的变化更为明显以及土壤有机质水平较高造成的。相关研究表明全球各生态系统的10的变化范围在1.3 ~ 3.3之间波动,中值为2.40[2]。本研究中小獐毛、鸢尾、芨芨草、黑果枸杞和撂荒地样地的10分别为2.30、1.24、1.20、1.57、1.28,中值为1.28。与前人研究成果[25]相比,盐湖沿岸群落土壤呼吸对温度的敏感程度更低,这可能是由环境因素导致的,盐湖夏季的日较差较低,温度小范围的变化降低了土壤呼吸的温度敏感程度,从而降低了10值。
土壤湿度是影响土壤呼吸的另一个重要因子。土壤湿度对土壤呼吸的直接影响是通过影响根和微生物的生理过程,对土壤呼吸的间接影响是通过影响底物和氧气的扩散。相关研究结果表明,土壤呼吸速率和土壤湿度之间存在正相关、负相关或是没有相关性[26]。本研究中,达坂城盐湖沿岸不同群落土壤呼吸与土壤湿度呈显著负相关,这与高艳红等[27]在腾格里沙漠得到的结论相同,这可能是因为土壤含水量超过了田间持水量的50%,限制了微生物活性,从而降低了微生物的异养呼吸。撂荒地土壤呼吸速率与土壤湿度呈正相关,能够解释土壤呼吸速率变化的38%,这与方晰等[28]对杉木人工林地土壤呼吸研究的结果和黄湘等[29]在塔里木河的研究结果一致。当土壤湿度在一个很小的范围内变化,土壤呼吸与土壤湿度之间将不会存在相关性,这是因为痕量般的水分变化可能会被其他因子所掩盖[30]。本研究中,撂荒地地表植被稀疏、地上地下生物量较低、砾石含量高、土壤水分状况较差,这都是限制土壤微生物活性和根呼吸的重要因子。当土壤含水量未超土壤水分阈值时,土壤微生物活性、根系呼吸也会出现正反馈。
土壤CO2通量与湿度的关系比较复杂,包括了生物、物理、化学机制,并且随着区域和时间尺度的不同而改变。土壤湿度的改变通常伴随着土壤微生物活性、根以及温度的变化,在干旱区还会影响到可溶性盐分的分布。在极端条件下的盐生荒漠系统中土壤经常会出现“碳汇”和无机CO2交换的现象[31-32],目前对土壤无机CO2通量形成的机制仍没有统一的定论,主要观点有碳酸盐系统的溶解/沉淀和CO2的溶解化学等。本研究区域位于达坂城盐湖沿岸,土壤类型属于盐渍土,在监测时段内,土壤未出现吸收CO2现象。
4 结论
在监测时段内,不同植物群落和撂荒地土壤呼吸日变化均呈单峰曲线,土壤呼吸峰值出现在13:00左右。各群落间及撂荒地土壤CO2日排放总量存在显著性差异,黑果枸杞群落土壤CO2日排放总量大于其他群落及撂荒地土壤;地上、地下生物量是影响土壤CO2排放总量的重要因子。土壤呼吸速率与土壤表层温度存在显著指数相关(2= 0.69 ~ 0.78),表明土壤温度是影响土壤呼吸的环境因子;小獐毛群落土壤对温度敏感强度大于其他土壤。不同样地土壤呼吸与土壤湿度相关性显著(2= 0.38 ~ 0.51),土壤湿度是决定达坂城盐湖土壤呼吸速率的重要因子之一。
盐湖沿岸土壤与盐生荒漠的土壤均为盐渍化土壤,而二者的生态功能却截然不同。我们的研究表明,在监测时段内土壤CO2未出现负通量,所以盐湖周边不同群落立地土壤表现为CO2的源,这可能与土壤盐分类型以及土壤水分条件有关。要进一步准确评估达坂城盐湖的生态系统碳收支状况,仍需对各植物群落的固碳潜力进行研究,从而为盐湖生态系统碳循环过程的系统研究提供科学数据。
[1] 杨涛, 姜文波, 孙希春, 等. 不同植被类型高山草甸土呼吸作用的研究[J]. 土壤学报, 1989, 26(2): 186–192
[2] Raich J W, Schlesinger W H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate[J]. Ellus, 1992, 44(B): 81–99
[3] 李彦, 王勤学, 马健, 等. 盐生荒漠地表水、热CO2输送的实验研究[J]. 地理学报, 2004, 59(1): 33–39
[4] 刘冉, 王勤学, 唐立松, 等. 盐生荒漠地表水热与二氧化碳通量的季节变化及驱动因素[J]. 生态学报, 2009, 29(1): 67–75
[5] 刘冉, 李彦, 刘燕. 盐生荒漠CO2通量对两个极端降水年份响应[J]. 生态学杂志, 2013, 32(10): 2545–2551
[6] 郑喜玉, 张明刚, 徐昶, 等. 中国盐湖志[M]. 北京: 科学出版社, 2002
[7] 杨磊, 贾宏涛, 胡毅, 等. 水分条件对巴音布鲁克高寒湿地CO2排放的影响[J].草业科学, 2015, 32(2): 168–173
[8] 刘跃辉, 艾力·买买提依明, 杨帆, 等. 塔克拉玛干沙漠腹地冬季土壤呼吸及其驱动因子[J]. 生态学报, 2015, 35(20): 6711–6719
[9] 张增信, 施政, 何容, 等. 北亚热带次生栎林和人工松林土壤呼吸日变化[J]. 南京林业大学学报(自然科学版), 2010, 34(1): 19–23
[10] Bontti E E, Decant J P, Munson S M, et al. Litter decomposition in grasslands of Central North America[J]. Global Change Biology, 2009, 15(5): 1356–1363
[11] Raich J W, Tufekcioglu A. Vegetation and soil respiration:Correlations and controls[J]. Biogechemistry, 2000, 48(1): 71–90
[12] 马杰, 于丹丹, 郑新军. 盐生荒漠土壤CO2通量及其环境影响因素[J]. 生态学杂志, 2013, 32(10): 2532–2538
[13] Xu M, Qi Y. Soil-surfer CO2efflux and its spatial temporal variations in a young ponderosa pine plantation in northern California[J]. Global Change Biology, 2001, 7: 667-677
[14] Jassal R, Black A, Novak M, et al. Relationship between soil CO2concentrations and forest-floor CO2effluxes[J]. Agricultural and Forest Meteorology, 2005, 130(3): 176–192
[15] Tang J, Baldocchi D D. Spatial-temporal variation in soil respiration in an oak-grass savanna ecosystem in California and its partitioning into autotrophic and heterotrophic components[J]. Biogeochemistry, 2005, 73(1):183–207
[16] 董自红, 蒋平安, 贾宏涛, 等. 新疆高山草甸草原植物群落夏季CO2排放日变化分析[J].新疆农业科学, 2007, 44(1): 1–5
[17] 王忆慧, 龚吉蕊, 刘敏, 等. 草地利用方式对土壤呼吸和凋落物分解的影响[J]. 植物生态学报, 2015, 39(3): 239–248
[18] 杨建军, 吕光辉, 张燕, 等. 艾比湖流域土壤呼吸日变化及水热因子影响[J]. 新疆农业科学, 2009, 46(2): 223–231
[19] 王春燕, 陈秋波, 袁坤, 等. 橡胶林土壤呼吸速率及其与土壤温湿度的关系[J]. 土壤学报, 2013, 50(5):119–128
[20] 张赛, 王龙昌, 张晓雨, 等. 紫色土丘陵区保护性耕作下旱地土壤呼吸及影响因素[J]. 土壤学报, 2014, 51(3):520–530
[21] 王忠媛, 谢江波, 王玉刚, 等. 温度对盐土和碱土土壤无机CO2通量的影响[J]. 生态学杂志, 2013, 32(10): 2525–2531
[22] 全权, 张震, 何念鹏, 等. 短期氮添加对东灵山三种森林土壤呼吸的影响[J]. 生态学杂志, 2015, 34(3): 797–804
[23] Luo Y Q, Wan S Q, Hui D F, et al. Acclimatization of soil respiration to warming in a tall grass prairie[J]. Nature, 2001, 413: 622–625
[24] 陈宝玉, 刘世荣, 葛建平, 等. 川西高山针叶林土壤呼吸速率与不同土层温度的关系[J]. 应用生态学报, 2007,18(6):1219–1224
[25] 张盼弟, 王旭, 陈宝瑞, 等. 呼伦贝尔地区贝加尔针茅草甸草原土壤CO2排放特征[J]. 应用生态学报, 2014, 25(2): 387–393
[26] 孔雨光, 张金池, 王因花, 等. 苏北淤泥质海岸典型防护林地土壤呼吸及其温度敏感性[J]. 生态学报, 2009, 29(8): 4084–4093
[27] 高艳红, 张志山, 刘立超, 等. 腾格里沙漠植被重建对土壤呼吸的影响. 土壤学报, 2009, 46(4):627–633
[28] 方晰, 田大伦, 项文化. 杉木人工林林地土壤CO2释放量及其影响因子的研究. 林业科学, 2005, 41(2): 1–7
[29] 黄湘, 李卫红, 陈亚宁, 等. 塔里木河下游荒漠河岸林群落土壤呼吸及其影响因子[J]. 生态学报, 2007, 27(5): 1952–1959
[30] 李虎, 邱建军, 王立刚. 农田土壤呼吸特征及根呼吸贡献的模拟分析[J]. 农业工程学报, 2008, 24(4): 14–20
[31] Liu J B, Feng W, Zhang Y Q. Abiotic CO2exchange between soil and atmosphere and its response to temperature[J]. Environmental Earth Sciences, 2015, 73: 2463–2471
[32] Emma L, Angela M, Detweiler L T. Assessing the role of alkaline soils on the carbon cycle at a playa site[J]. Environmental Earth Sciences, 2013, 70(3): 1047–1056
Soil Respirations Under Different Plant Communities in Dabancheng Salt Lake of Xinjiang
LI Dianpeng, YAO Meisi, HAN Dongliang, WANG Liping, HU Bao’an, WANG Ningning, JIANG Dayong, ZHANG Kai, ZHU Xinping, JIA Hongtao*
(College of Grassland and Environment Sciences, Xinjiang Agricultural University, Urumqi 830052, China)
The distribution of salt lakes in Xinjiang is broad but uneven. The carbon cycle of the salt lake ecosystem is an important component and has an important impact on carbon equilibrium in the ecosystem of the arid region. However, the research on soil respiration in salt lakes in the arid region is relative weak. A study was conducted in order to explore soil respiration and its influential factors of different plant communities along the Salt Lake in Dabancheng of Xinjiang.,,,were taken as the main plant objects with the abandoned farmland around as the control, soil respiration were measured by LI-COR 8100 (Li-COR, Lincoln, NE, USA). The results showed soil respiration of Salt Lake coastal had obvious diurnal variation and appeared in the pattern of single peak, the highest point occurred at 13:00 while the lowest occurred at around 07:00. The rates of soil respiration ranged from 1.77 to 3.48 µmol/(m2·s) in CK and from 0.89 to 4.34 µmol/(m2·s) under different communities, with an order of. Underground biomass influenced soil CO2emissions, the daily average CO2emission flux of soil respiration in the study region was 209.28 mg/(m2·d). Soil temperature at 5 cm depth had significant correlation with soil respiration rate, which is the dominant factor in determining the diurnal variation of soil respiration and could explain 69%-78% of it. The10values calculated based on soil temperature at 5 cm depth were 2.30, 1.24, 1.20, 1.57, and 1.28, soil respiration was most sensitive to soil temperature under. Except for abandoned land, there were significantly negative correlation (2= 0.38-0.51) between soil respiration and soil moisture. The above results disclosed further soil respiration variation and influential factors of Slat Lake ecosystem in the arid region, which is helpful in estimating carbon equilibrium and sequestration.
Arid region; Salt take; Carbon cycle; Soil respiration; Influential factors
10.13758/j.cnki.tr.2017.03.028
S154.1
A
国家自然科学基金项目(31560171)和国家大学生创新训练计划项目(201510758004)资助。
(hongtaojia@126.com)
李典鹏(1992—),男,湖南新华人,在读本科生,主要研究领域为土壤生态系统物质循环。E-mail: LLdp05120@126.com
