基于种间竞争模型的毒杂草入侵空间分布模拟研究
2017-07-20魏玉梅
刘 华, 金 鑫, 石 磊, 蒋 芮, 魏玉梅
1 西北民族大学数学与计算机科学学院,兰州 730030 2 西北民族大学实验中心,兰州 730030
基于种间竞争模型的毒杂草入侵空间分布模拟研究
刘 华1,*, 金 鑫1, 石 磊1, 蒋 芮1, 魏玉梅2
1 西北民族大学数学与计算机科学学院,兰州 730030 2 西北民族大学实验中心,兰州 730030
以天然草场中毒杂草与可食牧草为研究对象,在种间竞争的生态数学模型的基础上引入入侵扩散因素建立毒杂草入侵扩散模型。采用元胞自动机理论将竞争模型扩展到空间网络进行模拟研究,分析毒杂草属的空间分布类型,为毒杂草的控制提供数据支持。研究表明:(1)在入侵扩散作用下,毒杂草与可食牧草的共存平衡点由一个增加为两个,增加了共存的可能性;(2)入侵扩散作用影响了毒杂草种群的空间分布特征,减少了种群空间分布的聚集程度。
元胞自动机;空间分布;计算机模拟
生物入侵是指生物由原分布区域经由自然的或人为的途径扩展到一个新的区域,繁殖并维持下去[1]。近年来, 随着全球经济一体化、国内外贸易的迅速发展, 世界各地生物入侵现象愈加严重[2]。我国因外来物种入侵造成的损失相当惊人,每年仅几种外来物种造成的经济损失就达到574亿元人民币[3]。据世界自然保护联盟报道,全球范围内入侵物种对生物多样性的威胁高居前5位, 外来生物入侵是导致原生物种衰竭、生物多样性减少的重要原因之一[4]。如今, 生物入侵对我国生物多样性的影响越来越突出, 在一定程度上威胁我国生物多样性和生态系统稳定性[4]。
我国西北地区因其独特的地形条件和地理位置,加之资源过度开发和不合理利用,导致长期以来生物灾害严重、生态环境脆弱,而外来物种入侵加剧了这一矛盾。外来入侵植物可降低土壤的营养水平, 这主要是由于竞争、落叶的营养贫乏或难分解, 积累盐分改变土壤pH 值等造成的[2]。在天然草场中,入侵的毒杂草不仅与可食牧草争夺水、肥料、光照等资源抑制可食牧草的生长、降低可食牧草的产量,而且影响可食牧草的品质,更有甚者,有的杂草本身就是毒草,家畜误食,造成中毒、死亡。原产于中美洲的紫茎泽兰(Eupatoriumodoratum),约在20世纪50 年代初从中缅、中越边境传入云南南部,至今已占据了四川凉山州1/5 的草场,正以每年约30 km 的速度向北、向东扩散蔓延,当地畜牧业遭受了空前打击,每年经济损失达数千万元[3]。因此,牧民都希望草场中的毒杂草越少越好,可食牧草越多越好,而要想控制草场中毒杂草的数量,就首先需要了解毒杂草与可食牧草在草场中的空间分布情况。
本文假设毒杂草与可食牧草存在对环境资源的竞争关系,采用元胞自动机理论对种间竞争模型进行空间模拟。元胞自动机的基本思想就是将系统分解为多个元胞,每个元胞看成一个单元,系统中各个相邻单元之间存在一定的交互相互关系,而每个单元的状态都是整个系统在一些简单的规则作用下不断进化的结果[5- 7]。
在对种间竞争模型进行模拟的时候,本文引入了空间扩散因素,从而得到了毒杂草入侵的空间分布情况。
1 毒杂草与可食牧草的种间竞争模型
假设毒杂草与可食牧草种间存在相互作用,同时假设单一物种独自在同一个草场中生存时,数量的演变均遵从Logistic规律。于是得到毒杂草与可食牧草的种间竞争模型为[8- 10]:
(1)
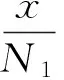

表1 种群竞争模型的平衡点及稳定性
2 毒杂草入侵空间扩散模型
毒杂草和可食牧草作为两个物种在单个斑块中将产生竞争,而且毒杂草是外来物种,在生长过程中会从密度高的斑块向密度低的斑块发生迁移,考虑到以上因素建立单个斑块毒杂草入侵空间扩散模型
(2)

2.1 具有空间扩散的毒杂草入侵空间扩散模型的稳定性分析
讨论模型(2)的稳定性,首先要计算平衡点,令:
(3)
则平衡点为:
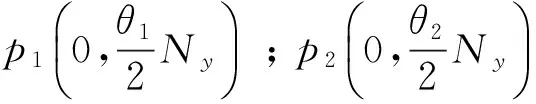
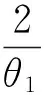
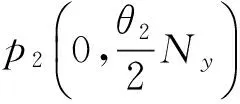
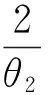
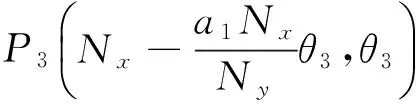
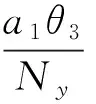
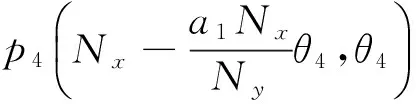
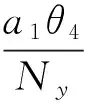
其中:
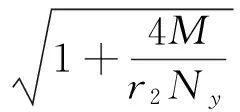
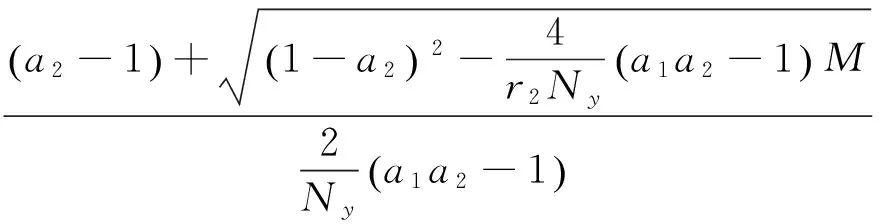
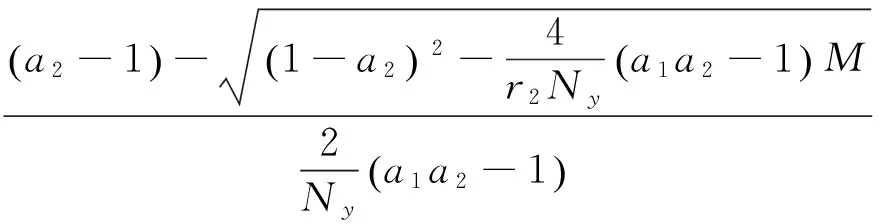
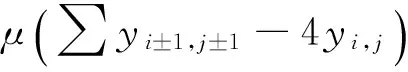
在以上两个模型中,模型(1)是毒杂草与可食牧草的种间竞争模型,该模型只考虑了当两个种群在同一个自然环境中生存时它们的相互竞争关系。而模型(2)是毒杂草入侵的空间扩散模型,该模型考虑到了毒杂草和可食牧草作为两个物种在单个斑块中将产生竞争,而且毒杂草是外来物种,在生长过程中会从密度高的斑块向密度低的斑块发生迁移这一重要因素。可以说模型(2)是模型(1)的改进。所得到的结果中:没有加入扩散项时,模型(1)的四个平衡点包括零平衡点和毒杂草灭绝的平衡点以及一个共存的非零平衡点;加入扩散项之后,模型的零平衡点和毒杂草灭绝的平衡点消失,而且毒杂草与可食牧草的共存的非零平衡点由一个变为两个。说明在扩散作用下,两种群的可能出现两种稳定情况,而且毒杂草续存的可能性增加。
3 毒杂草入侵的空间分布模拟
将单一斑块中的毒杂草与可食牧草的相互竞争扩展到空间上,在100×100的二维网格平面模拟两物种(毒杂草与可食牧草)的种群动态,时间标度t为单位时间步长。初始时刻可食牧草与毒杂草的数量都是随机的。在二维网格中,网格之间的物种会发生迁移,假定物种在冯诺依曼邻体(即只考虑相邻的每个斑块边的4个斑块)由高密度向低密度发生迁移(也可推广为摩尔邻体即与斑块边和顶点相邻的八个斑块发生迁移)。迁移率为0.25倍的密度差,即μ=0.25。
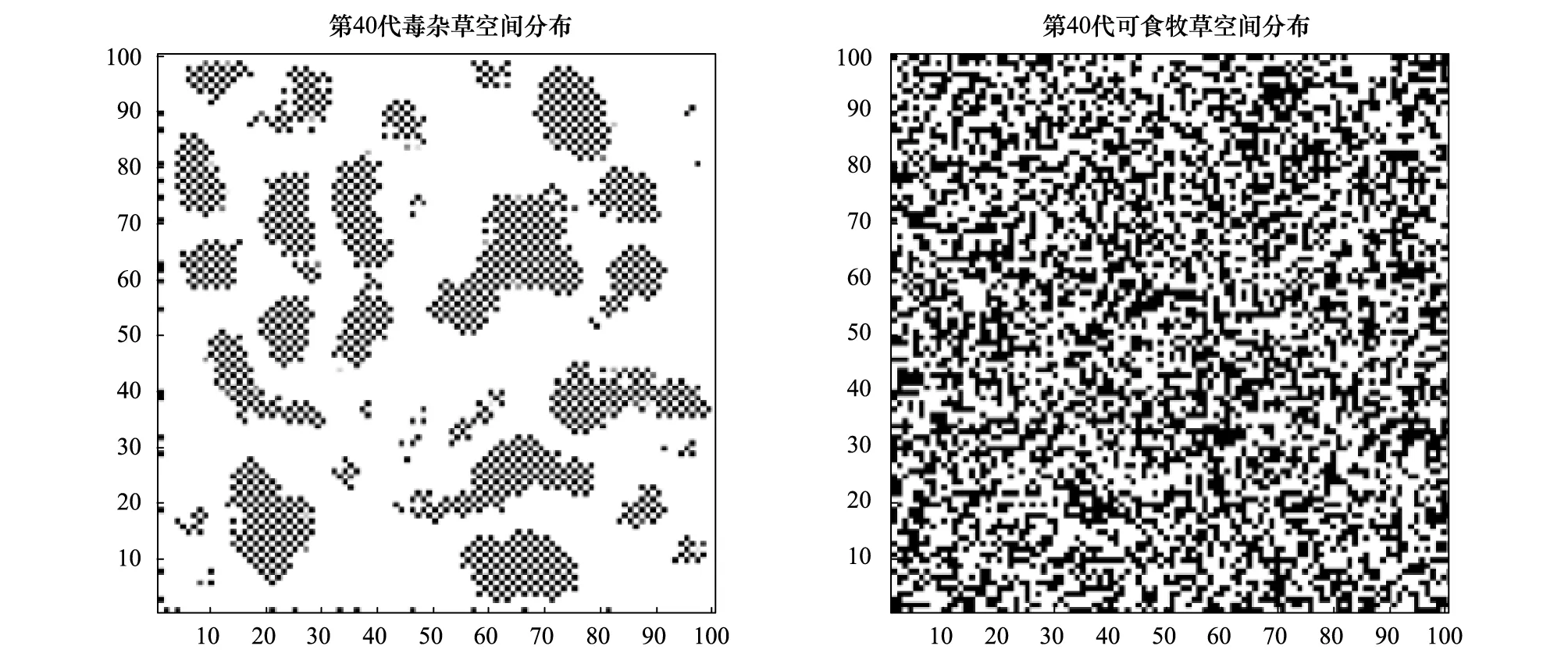
图1 第40代可食牧草与毒杂草空间分布Fig.1 Spatial distribution of fortieth generations of edible grass and poisonous weeds

图2 第60代可食牧草与毒杂草空间分布Fig.2 Spatial distribution of sixtieth generations of edible grass and poisonous weeds
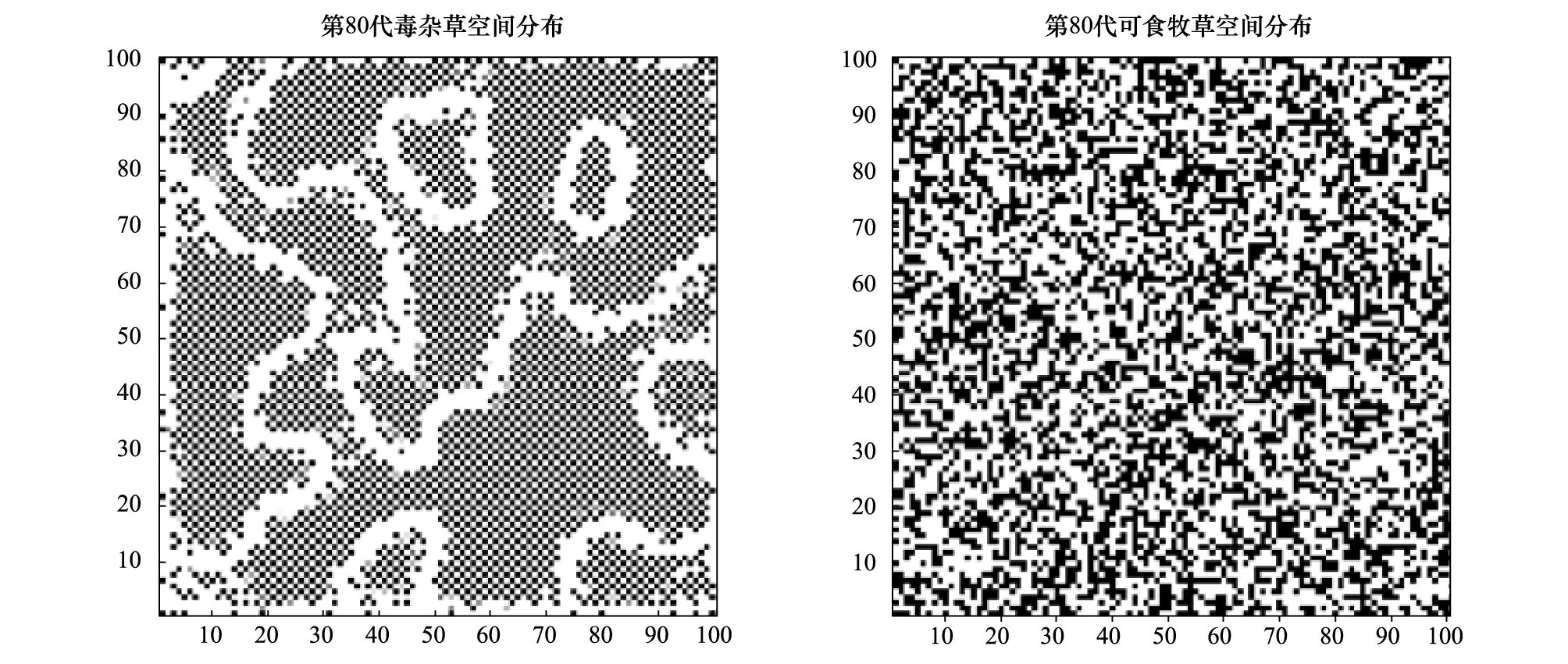
图3 第80代可食牧草与毒杂草空间分布Fig.3 Spatial distribution of eightieth generations of edible grass and poisonous weeds
现假定r1=0.45,r2=0.45,a1=0.5,a2=0.7,最大容量N1为40,N2为60,考虑到毒杂草为外来物种,因此认为该物种在入侵过程中具有扩散过程,而作为本地物种的可食牧草在空间上是没有扩散作用的,通过Matlab进行编程模拟。图1、图2及图3分别以第40,60,80代可食牧草与毒杂草空间分布情况为例给出空间分布的模拟结果,其中白色代表生存,黑色代表已灭绝。图4和图5显示:可食牧草生物量大概在18代之后趋于不变,而毒杂草的生物量大概在18代之后在小范围内波动。
4 毒杂草入侵的空间分布假设检验

(1)利用分散指标C对数据进行检验
根据数据计算得:
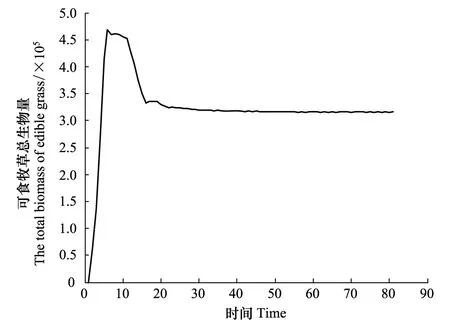
图4 可食牧草总生物量随时间变化Fig.4 The total biomass of edible grass changed with time
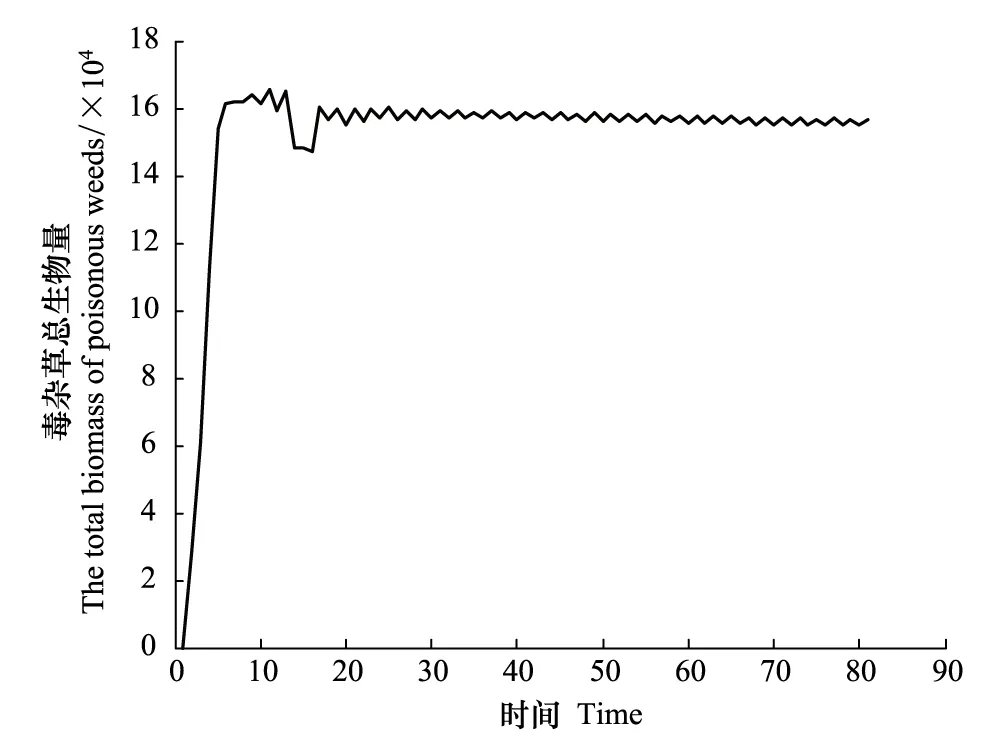
图5 毒杂草总生物量随时间变化Fig.5 The total biomass of poisonous weeds changed with time
又因为
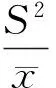
所以
C的概率为95%的置信区间为
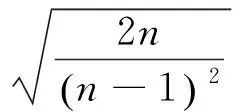
7.23890084 >1.028287,7.23890084> 0.9717129
显然C落入区间外,故模拟数据属于聚集型分布。
(2)利用David&Moore(1954)方法对数据进行检验
首先假设模拟数据属于随机分布,则有
由上可知
求得

取自由度
i=10000-1=9999
经计算得:
所以
显然不接受假设,所以模拟数据属于聚集型分布。
(3)利用平均拥挤度指标Lloyd,M. (1967)对数据进行检验
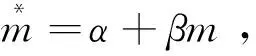
(5)计算Morisita′sindex对数据进行检验
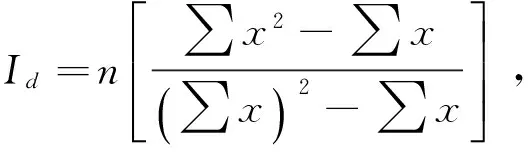
计算临界点
Uniformindex
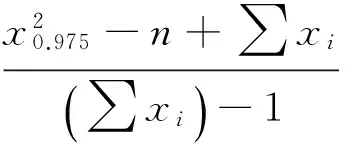
Clumpedindex
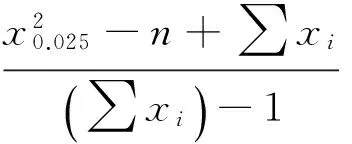
所以
Id≥Mc>1
计算StandardizedMorisitaindex
因为IP=0.50001535,得Ip>0,所以模拟数据属于聚集分布型。
采取以上方法,通过计算机模拟数据并对所有80代数据进行检验,得到的结果如下所示,其中字母“U”代表均匀分布,字母“A”代表聚集分布。充分说明毒杂草入侵可食牧草时,以较强的竞争能力,空间分布型从开始的均匀分布逐渐演变成最终的聚集分布。
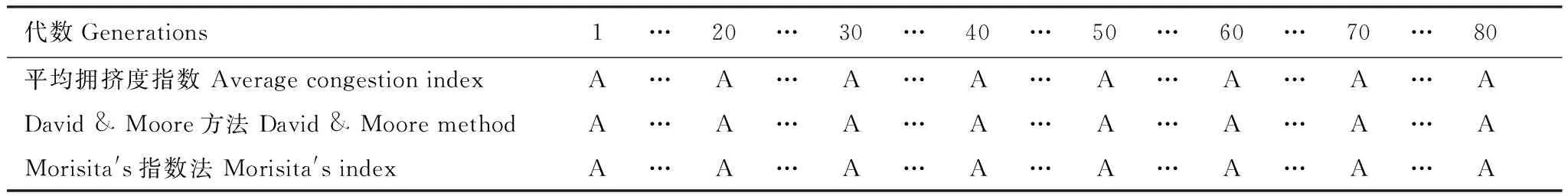
表2 加入扩散项前毒杂草空间分布结构(1到80代均为A)
由表2可以看出,在加入扩散项之前,即扩散系数为0时,毒杂草在第1代到第80代一直为聚集分布。
由表3可以看出,在加入扩散项之后,且扩散系数取0.25时,毒杂草在第1代至第24代时为均匀分布,从第25代以后开始变为聚集分布。
计算1至80代加入扩散项的植物入侵模型的空间分布相关参数,毒杂草从第1代开始到第24代为均匀分布,从第25代开始其空间分布已经由均匀分布向聚集分布发生转变,此后一直是聚集分布。综合两个表可以发现,外来植物物种的入侵及空间扩散可以影响植物物种的空间分布的格局,减少了种群空间分布的聚集程度。
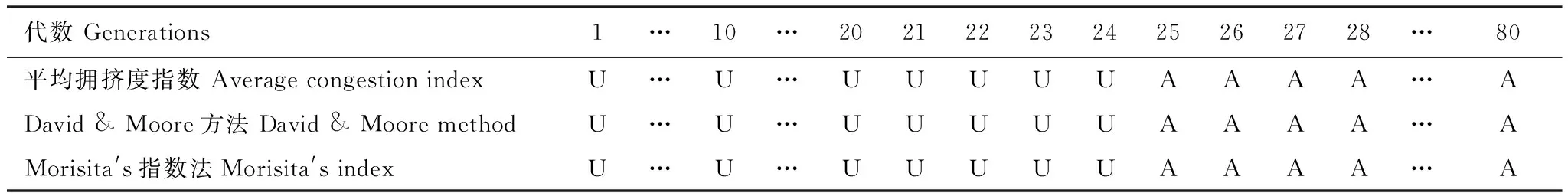
表3 加入扩散项后毒杂草空间分布结构(1到24代为U,25代以后为A)
在通过Matlab进行编程模拟的过程中,我们观察到:当扩散系数、固有增长率、可食牧草与毒杂草的生存能力等因素变化时,毒杂草与可食牧草的空间分布发生显著的变化,同时,毒杂草与可食牧草的总数量随时间的变化也很明显。当取扩散系数为0,r1=0.45,r2=0.45,a1=0.5,a2=0.7时,由于没有受到扩散因素的影响,毒杂草从第1代开始到第80代都是聚集分布;当取扩散系数为0.1,其余参数不变时,由于受到扩散因素的影响,可食牧草与毒杂草的分布结构发生了变化,毒杂草在第1代至第14代时为均匀分布,从第15代以后变为聚集分布;当取扩散系数增加到0.2时,毒杂草从第一代至第80代全部为聚集分布;当再取扩散系数为0.25时,可食牧草在第1至24代为均匀分布,从第25代开始为聚集分布;而当取扩散系数继续增加到0.3时,由于扩散系数过大,且可食牧草与毒杂草竞争激烈,可食牧草在很短的时间内灭绝,而毒杂草在第1至9代为均匀分布,从第10代以后为聚集分布;当扩散系数达到0.4,由于扩散系数太大,可食牧草很快就灭绝了,而毒杂草在第1代时为均匀分布,从第2代开始便属于聚集分布。
从以上的模拟过程中我们观察到:当扩散系数从0逐渐增大至0.4的过程中,可食牧草的灭绝速度越来越快。同时,毒杂草在受到扩散因素影响的时候,其均匀分布保持的时间长度也有很大变化,当扩散因素从0增加到0.25时,毒杂草均匀分布保持的时间逐渐变长,而当扩散因素从0.25增加到0.4时,毒杂草均匀分布保持的时间逐渐变短。
5 结果讨论
本文在假设参数时,认为毒杂草的生存能力略大于牧草,故在模拟中参数取值只考虑这一类情况。由于数据是模拟出来的,可能并不能很贴切的描述实际的情况,但这也可以从一个方面来了解研究毒杂草空间分布,进而进行有关的毒杂草问题的处理。本文元胞自动机的规则是自己设定的,并且采用的是四邻域,这个是可以使用八邻域、二十四邻域,得到进一步推广延伸,也就是说本文使用的元胞自动机为冯诺依曼型,可推广为摩尔型以及扩展摩尔型[15- 19]。
[1] Anderson R M, May R M, Joysey K, Mollison D, Conway G R, Cartwell R, Thompson H V, Dixon B. The invasion, persistence and spread of infectious diseases within animal and plant communities. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 1986, 314(1167): 533- 570.
[2] 安志兰, 郭笃发, 褚栋, 刘国霞, 范仲学. 生物入侵对我国生态环境的影响及其控制策略. 山东农业科学, 2007, (1): 87- 91.
[3] 宋晓丰, 叶桂峰. 我国外来生物入侵的现状、危害及防治研究. 生物学通报, 2008, 43(7): 24- 26.
[4] 林培群, 余雪标. 生物入侵的现状及其危害与防治. 华南热带农业大学学报, 2006, 12(2): 61- 65.
[5] 刘华, 刘志广, 高猛, 张丰盘, 李自珍. 有色环境噪音下空间种群的一致性与灭绝. 兰州大学学报: 自然科学版, 2008, 44(4): 99- 102.
[6] 潘士虎. 用元胞自动机模型模拟教学楼疏散过程[D]. 桂林: 广西师范大学, 2010.
[7] 吕凯. 元胞自动机的研究及模型的建立[D]. 哈尔滨: 哈尔滨理工大学, 2007.
[8] 刘华, 刘志广, 苏敏, 李自珍. 聚集效应对宿主—寄生物种群模型动态行为的影响. 山东大学学报: 理学版, 2008, 43(8): 31- 34, 37- 37.
[9] 付军, 朱宏. 两种群相互作用模型的定性分析及应用. 吉林师范大学学报: 自然科学版, 2006, 27(3): 7- 8.
[10] 杨正清. 两种群相互作用模型的全局渐近稳定性. 生物数学学报, 1994, (3): 33- 36.
[11] 姜启源, 谢金星, 叶俊. 数学模型(第四版). 北京: 高等教育出版社, 2011: 223- 246.
[12] Matis J H, Zheng Q. Describing the spread of biological populations using stochastic compartmental models with birth. Mathematical Biosciences, 1995, 126(2): 215- 247.
[13] 王德好. 毒麦空间分布型及其抽样面积的研究. 杂草科学, 1990, (4): 28- 30.
[14] 郭树江, 杨自辉, 王多泽, 李得禄, 李爱德, 詹科杰, 王强强. 民勤绿洲一荒漠过渡带植物物种多样性及其优势种群空间分布格局研究. 水土保持研究, 2011, 18(3): 92- 96.
[15] 贾斌, 高自友, 李克平, 李新刚. 基于元胞自动机的交通系统建模与模拟.北京: 科学出版社, 2007: 9- 18.
[16] 刘永欣. 基于元胞自动机的交通流研究[D]. 成都: 西南交通大学, 2003: 9- 27.
[17] 李群, 雷永林, 侯洪涛, 朱一凡. 仿真模型设计与执行. 北京: 电子工业出版社, 2010: 355- 362.
[18] Gustafsson L, Sternad M. Bringing consistency to simulation of population models-Poisson simulation as a bridge between micro and macro simulation. Mathematical Biosciences, 2007, 209(2): 361- 385.
[19] Engen S. Stochastic growth and extinction in a spatial geometric Brownian population model with migration and correlated noise. Mathematical Biosciences, 2007, 209(1): 240- 255.
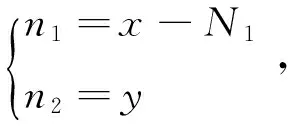
取得线性近似系统为:
当a1<1,a2>1时,特征根有负实部,p1(N1,0)稳定,否则不稳定(其中a1<1是根据相轨线分析的结果添加的[11],这也与生态背景相符合,当a1<1时,毒杂草的竞争力弱于可食牧草,从而灭绝)。
2)同理可得[11],当a1>1,a2<1时,p2(0,N2)稳定,否则不稳定。
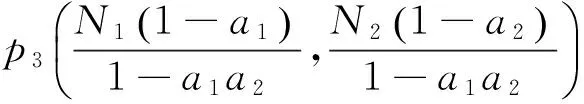
取得线性近似系统为:
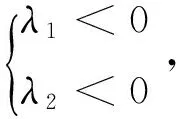
附录2:证明:为简化符号,证明中xi,j均记为X,yi,j均记为Y。
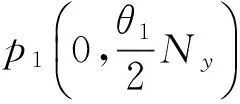
取得线性近似系统:
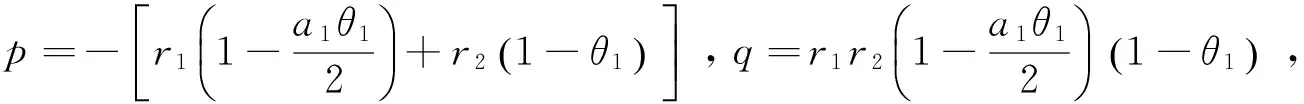
取得线性近似系统:
其特征方程为:
同理若p>0且q>0,则平衡点p3稳定[11],

由p>0得:
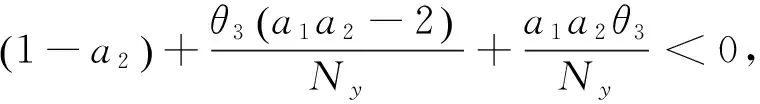
Spatial distribution of poisonous weed invasion based on inter-species competition models
LIU Hua1,*, JIN Xin1, SHI Lei1, JIANG Rui1, WEI Yumei2
1SchoolofMathematicsandComputerscience,NorthwestUniversityforNationalities,Lanzhou730030,China2ExperimentalCenter,NorthwestUniversityforNationalities,Lanzhou730030,China
In the present study, a spatial diffusion model of poisonous weed invasion by using inter-species competition model was developed based on poisonous weeds and edible forage in native pastures. Using the cellular automata theory, the competition model was extended to the spatial network to simulate and study the spatial distribution patterns of poisonous weeds, which will provide data supporting their control. The results showed that (1) under the influence of invasion and diffusion, the coexistence equilibrium point of the poisonous weeds and the edible grass could be increased by an increase of two, which increased the possibility of coexistence; and (2) the spatial distribution of the weed population was affected by invasion and diffusion, and the aggregation of the spatial distribution of the population was reduced.
cellular automata; space distribution; computer simulation
国家自然科学基金资助项目(31260098,11361049,31560127);甘肃省自然科学基金(145RJZA158,1506RJZA274);国家民委中青年英才计划资助(〔2014〕121号);西北民族大学研究生科研创新项目(YXM2014172, YXM2015178);2013年西北民族大学创新团队项目资助
2016- 04- 13; 网络出版日期:2017- 02- 22
10.5846/stxb201604130679
*通讯作者Corresponding author.E-mail: liuhuaemail@foxmail.com
刘华, 金鑫, 石磊, 蒋芮, 魏玉梅.基于种间竞争模型的毒杂草入侵空间分布模拟研究.生态学报,2017,37(11):3765- 3775.
Liu H, Jin X, Shi L, Jiang R, Wei Y M.Spatial distribution of poisonous weed invasion based on inter-species competition models.Acta Ecologica Sinica,2017,37(11):3765- 3775.
