辽宁地区两种菊科入侵植物与本地植物光合特性比较*
2017-07-18魏子上陈新微杨殿林皇甫超河
魏子上,陈新微,杨殿林,皇甫超河**
(1.农业部环境保护科研监测所 天津 300191;2.沈阳农业大学植物保护学院 沈阳 110866)
辽宁地区两种菊科入侵植物与本地植物光合特性比较*
魏子上1,2,陈新微1,2,杨殿林1,皇甫超河1,2**
(1.农业部环境保护科研监测所 天津 300191;2.沈阳农业大学植物保护学院 沈阳 110866)
为明确外来植物生理生态学性状及其与其强入侵性的关系,本研究采用野外原位气体交换参数测定的方法,分析了菊科入侵植物豚草、三裂叶豚草与其共生本地植物紫菀光合特性和叶片性状的差异。结果表明,豚草、三裂叶豚草、紫菀3种植物的光饱和点(LSP)均高于800 μmol·m-2·s-1,且两种入侵植物的LSP显著高于紫菀,表观量子效率(AQY)则显著低于紫菀;3种植物的最大净光合速率(Pnmax)大小顺序为豚草>三裂叶豚草>紫菀,其中,豚草和三裂叶豚草的Pnmax分别比紫菀高出151.28%、82.80%,且差异显著。两种入侵植物的比叶面积(SLA)、叶片的单位质量氮含量(Nmass)、叶片单位质量磷含量(Pmass)、光合能量利用效率(PEUE)、光合氮利用效率(PNUE)均显著高于紫菀,但3者叶片单位质量建成成本(CCmass)差异不显著。综上所述,辽宁的两种菊科入侵植物相对于本地共生种来说有着较高的气体交换参数和叶片特性指标,且其光合特性和叶片特性也存在着密切的联系,表现在这些菊科入侵植物有着更高的Pnmax、PNUE、PEUE和水分利用效率(WUE)等光合特性指标和能量利用指标,使得入侵植物能够更有效地捕获和利用环境资源,成为其得以成功入侵的原因之一。
菊科入侵植物;光合特性;最大净光合速率;叶片特性;资源利用效率
在经济全球变化日益加速的背景下,生物入侵已对入侵区的生态环境、农林业经济和人民健康造成了严重的威胁和损害,成为21世纪最瞩目的全球性环境问题之一[1-3]。入侵植物通常生长快速,且具有生态适应能力强、繁殖能力强和传播能力强的特点[4],并以此排挤本地植物。入侵种入侵到某一个地区后开始扩散建立自己的种群,然而并不是所有的本地植物都被竞争或排挤下去,对比入侵种和当地优势伴生种群的生理生态特性差异可以作为探求入侵种入侵机制的突破口。除了生长速度,与入侵植物强大竞争能力相关的指标还包括高光合速率,低建成成本(construction costs,CC)以及高总叶面积[5]。比叶面积(specific leaf area,SLA)又是表征叶片建成成本的间接指标[5-6]。较低的建成成本(也即高比叶面积)意味着较高的植物碳同化产物利用效率以及较高的光合氮素利用效率[7-8](photosynthetic nitrogen-use efficiency,PNUE),而这又有助于植物快速生长[6],使得入侵种能够获得生长优势[9-10]。例如,Baruch和Goldstein[11]在夏威夷州采集了30种入侵植物和34种本地植物,通过对比其叶片重要的功能特性例如比叶面积、叶片建成成本、叶片氮含量等和光合特性如叶片同化速率,研究发现入侵种较本地种都有着比较高的比叶面积、同化速率,但有着比较低的叶片建成成本。Alpert等[12]、王睿芳等[13]提出类似研究应该比较亲缘关系比较近的入侵种和本地种,例如同科或是同属的植物。王俊峰等[14]研究发现飞机草(Chromolaena odorata)和其同科兰花菊三七(Gynura japonica)两种植物适应光环境的策略有明显不同,相比本地植物飞机草有着较高的光能利用效率。王晓红等[15]通过研究入侵植物小飞蓬(Conyza canadensis)及其主要伴生植物山苦荬(Ixeris chinensis)和鸭跖草(Commelina communis)的光合特性发现,小飞蓬具有较高的光合速率和物质积累能力,较高的生产力是其成功入侵的重要因素之一。
我国最常见的入侵植物来自菊科、禾本科和十字花科[1],其中菊科植物是入侵种类最多的类群之一[16]。豚草(Ambrosia artemisiifolia)是原产于北美的一年生恶性杂草,属于菊科向日葵族豚草属,是世界公认的恶性入侵杂草[17],其花粉是过敏性鼻炎、枯草热和季节性哮喘等疾病的重要过敏原,严重威胁人类健康[18]。三裂叶豚草(Ambrosia trifida)同样也是原产于北美的一年生恶性杂草,于20世纪40年代传入我国东北,现在已成为我国北方最主要的恶性入侵杂草之一[19]。该入侵种能在入侵地形成单一优势种群,造成农作物减产甚至绝产,威胁自然生态系统生物多样性,且易引发易感人群的枯草热[20-21]。目前,基于某一类群(科、属)内入侵植物与非入侵植物之间的生理生态特性比较研究很少,由于同科属植物间有着相同的进化背景和相近的生物学特性,开展此类研究更有助于揭示入侵种得以扩展危害的特有生理生态学机制。鉴于此,本试验比较了不同菊科入侵植物与本地菊科共生种之间的光合特性和叶片性状,以期为探讨菊科入侵植物对环境光强变化的响应机制,并对潜在入侵植物风险评估和制定有效阻控措施提供科学依据。
1 材料与方法
1.1 研究区域概况
采样区位于辽宁省沈阳市棋盘山(42°1′58″N,123°51′43″E,海拔192 m),为豚草和三裂叶豚草重发生区。该区属于中温带大陆性季风气候,冬寒多雪,时间较长;夏热多雨,时间较短。全年平均气温7.41℃,无霜期148 d,年平均降水量为750 mm,平均相对湿度为65%~70%。样地为豚草和三裂叶豚草两种菊科入侵植物典型共同发生区,主要植被构成为入侵植物与本地植物自然混生群落,各入侵种呈斑块状分布。植物紫菀(Aster tataricus)为同科伴生优势非入侵植物,伴生杂草还有葎草(Humulusscandens)、苦荬菜(Lactuca indica)、狗尾草(Setaria viridis)等。土壤类型为棕壤土,土壤pH 6.4,有机质14.62 g·kg-1,全氮 1.53 g·kg-1,全磷 0.99 g·kg-1。
于2015年8月27日开展试验的测定和样品采集,此时供试菊科入侵植物与本地植物已经完成叶片形态建成,生育时期为营养生长盛期至初花期。
1.2 试验方法
每一种植物设置1个试验区组,面积为10 m×10 m,每区组内随机设置5个1 m×1 m的小样方,每个小样方内随机选取生长相对一致的具有代表性的植株10株,并在其对应位置分别随机采集30片健康完整的成熟叶片,因此每种植物采集叶片数5×30=150。5个小样方样品采集均设5次重复。采用Li-3100A叶面积仪(Li-Cor,Lincoln,Nebraska,USA)测定叶面积,随后将叶片放入自封袋,置于冰盒中低温保存,用于测定叶绿素含量、叶片N、P含量等理化指标。同时用土壤采样器取各植物根围0~10 cm的土壤样品,带回实验室供理化分析。
光合-光强(Pn-PAR)响应测定:于9:00—11:00在各小样方中选取菊科入侵植物和本地种处于营养生长期的植株。采用LI-6400便携式光合测定仪(Li-Cor Inc.,Lincoln,NE,USA)对两种菊科入侵植物和本地植物功能叶的净光合速率(net photosynthetic rate,Pn)、蒸腾速率(transpiration rate,Tr)、气孔导度(stomatal conductance,Gs)等指标进行测定[22]。叶室温度设为25℃,CO2浓度控制在400 μmol·mol-1,设置有效光合辐射(μmol·m-2·s-1)梯度为2 000、1 500、1 000、800、500、300、100、50 和0,利用自动测量程序进行光合-光强响应的测定[23]。测定时,均选取完全展开的成熟叶片,测定时保持叶片自然生长角度不变,每个叶片测3次,每区组重复测定5次。
光合-光强响应曲线采用Ye[24]、叶子飘等[25]提出的直角双曲线修正模型进行拟合:

式中:Pn为净光合速率(μmol·m-2·s-1);I为光合有效辐射(μmol·m-2·s-1);α为AQY(apparent quantum yield,表观量子效率),即植物叶片光响应曲线在I=0时的斜率,也被称为光响应曲线的初始斜率或初始量子效率;β和γ分别为独立于I的系数(m2·s·μmol-1);Rd为暗呼吸速率(mg·h-1·g-1)。
水分利用率( water use efficiency,WUE)的计算公式[26]:

式中:Pr为光合速率(μmol·m-2·s-1);Tr为蒸腾速率(mmol·m-2·s-1)。
叶绿素含量的测定:把采集的新鲜植物叶片用去离子水清洗干净,用打孔器切取干净的叶片,然后切成长细丝,用80%的丙酮浸提细丝至其完全变为白色为止,于663 nm和645 nm处读取光密度(记为OD663、OD645),按照下列公式计算叶绿素含量[27]:
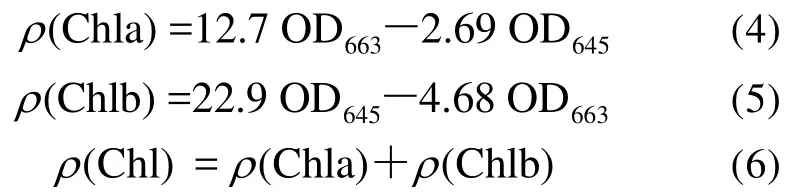
式中:ρ(Chla)、ρ(Chlb)分别为叶绿素a、叶绿素b的含量(mg·L-1);ρ(Chl)为叶绿素总量(mg·L-1)。
叶片元素含量的测定:植物样品于105℃下杀青30 min,然后65℃烘干至恒质量,叶片粉碎过筛,混匀后保存在塑封袋中备用。植物的全N采用开氏消煮法测定;全P采用钼锑抗比色法测定[28-30]。
叶片灰分含量的测定:植物样品在马弗炉中500℃灼烧6 h,称量剩余残渣,记录重量。干重热值采用氧弹式热量计(HWR-15E,上海上立检测仪器厂)测定:取0.5 g左右植物样品粉末,经完全燃烧测定热值,每个样品分别测定5个重复,取其平均值作为干重热值[31]。
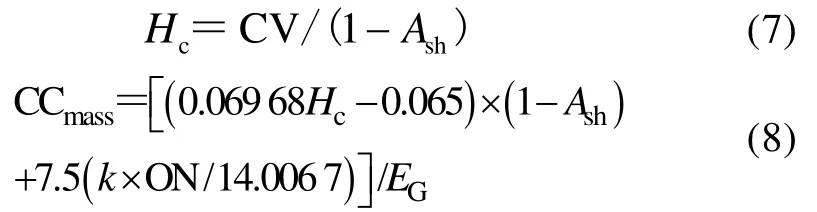
式中:Hc为去灰分热值(kJ·g-1);Ash为灰分含量(%);CV为干重热值(kJ·g-1);CCmass为叶片单位质量建成成本[g(glucose)·g-1];ON为有机氮含量,%;EG为生长效率;k为N的氧化态形式[31-32]。不同植物的平均叶片生长效率为0.87[33],对于每个样品,先以铵态氮(-3价)和硝态氮(+5价)作为N的氧化态形式分别计算,再按照土壤中的比例,求加权平均值作为叶片的CCmass。

式中:SLA为叶片比叶面积(cm2·g-1);ULA为单位叶片面积(cm2);DW为叶片干质量(g);CCarea为叶片单位面积建成成本[g(glucose)·m-2];Nmass为叶片单位质量N含量(mg·g-1);Narea为叶片单位面积N含量(g·m-2);PEUE为光合能量利用效率[µmol(CO2)·g-1(glucose)·s-1];PNUE为光合氮利用效率[µmol(CO2)·g-1·s-1]。
1.3 数据分析
参考陈新微等[23]的方法计算光饱和点(light saturation point,LSP)、光补偿点(light compensation point,LCP)、表观量子效率(apparent quantum yield,AQY)和最大净光合速率(maximum net photosynthetic rate,Pnmax)。所有数据应用SPSS 16.0对测定数据进行统计分析,应用Excel 2007对分析结果进行作图。各统计数据以平均值及标准误差表示,采用单因素方差分析(one-way ANOVA)和最小显著差数法(LSD法)进行不同物种间均值的方差分析和差异显著性比较(P=0.05)。
2 结果与分析
2.1 两种菊科入侵植物与本地植物对有效光合辐射的响应
由图1可见,随着有效光合辐射的增强,入侵植物豚草、三裂叶豚草及其共生植物紫菀的Pn均呈先增加后降低的趋势,其中,两种入侵植物的增幅均高于紫菀。当达到一定的有效光合辐射时,3种植物的Pn均达到最大值,即为光饱和点。豚草(R2=0.997,P<0.001)、三裂叶豚草(R2=0.999,P<0.001)、紫菀(R2=0.995,P<0.001)的Pn与有效光合辐射的相关性均达极显著水平(P<0.01),由直角双曲线修正模型可以计算出3种植物的光合响应参数(表1)。由表1可见,3种植物的LSP均高于800 μmol·m-2·s-1,其大小顺序为三裂叶豚草>豚草>紫菀,且差异显著;LCP的大小顺序为紫菀>三裂叶豚草>豚草,其中,本地种紫菀的LCP为30.736 μmol·m-2·s-1,显著高于豚草;而两种入侵植物的AQY均显著低于紫菀(P<0.05);3种植物的Pnmax大小顺序为豚草>三裂叶豚草>紫菀,入侵种豚草和三裂叶豚草的Pnmax分别比紫菀高出151.28%和82.80%,差异显著,显示两种菊科入侵植物对光能的利用率较高,在相同的有效光合辐射下,具备更强的光合同化能力。
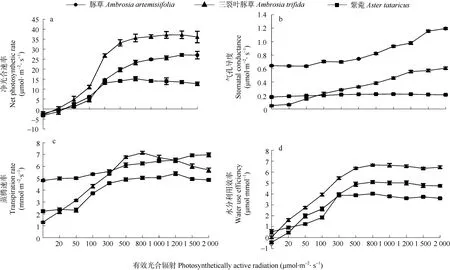
图1 辽宁菊科入侵植物豚草和三裂叶豚草与本地植物紫菀的光合-光响应曲线的比较Fig.1 Comparison ofPn-PAR curves of invasive plant species,Ambrosia artemisiifoliaandAmbrosia trifida,with native plant species,Aster tataricusin Liaoning

表1 辽宁菊科入侵植物豚草和三裂叶豚草与本地植物紫菀的光合响应特征参数Table 1 Photosynthetic parameters of invasive plant species,Ambrosia artemisiifoliaandAmbrosia trifida,and native plant species,Aster tataricusin Liaoning
随着有效光合辐射的增加,3种植物的Gs、Tr和WUE均有一定程度的增加,但在有效光合辐射大于500 μmol·m-2·s-1时,紫菀的Gs、Tr和WUE并未随着有效光合辐射的增强而增加,反而是稳定在一个恒定值附近,而其他两种入侵植物的Gs和Tr却一直表现为增加。由图1d可知,两种入侵植物的WUE随着有效光合辐射增加而增加的幅度明显高于本地种紫菀,显示两种入侵植物更高的水分利用效率。
2.2 两种菊科入侵植物与本地植物叶片特性指标的比较
由表2可见,豚草、三裂叶豚草的叶绿素含量均显著高于紫菀(P<0.05),从高到低依次为豚草>三裂叶豚草>紫菀。两种入侵植物的SLA、Nmass、叶片单位质量磷含量(leaf P content per unit mass,Pmass)、PEUE、PNUE均显著高于其本地种紫菀(P<0.05),但3种植物的CCmass差异不显著。

表2 辽宁菊科入侵植物豚草和三裂叶豚草与本地植物紫菀叶片特性的比较Table 2 Comparison of leaf traits of invasive plant species,Ambrosia artemisiifoliaandAmbrosia trifida,with native plant species,Aster tataricusin Liaoning
3 讨论
入侵植物可以改变自然群落生态系统基本的生物学特征[34-35]。对入侵植物而言,成功入侵的因素是多方面的,而其自身的生理特性对于其生存、建立种群入侵和扩展至关重要,有些外来植物具有比本地种更强的光能利用率、光合响应机制以及生理生态特性,从而使它们具有很强的入侵潜力[36]。环境和植物本身特性的相互作用共同决定植物的入侵能力,在繁殖能力和天敌压力相同的条件下,入侵种能够迅速扩散并取代当地植物,通常认为外来入侵植物能更有效地利用有限的环境资源,或在时间上可以利用本地种不能利用的资源[37]。
3.1 两种菊科入侵植物与本地植物气体交换参数之间的差异
植物对光的吸收能力和光合作用对生境的适应特征在很大程度上体现了植物生长与竞争的能力[38]。LSP和LCP反映了植物对强光和弱光环境的适应能力[39],AQY和Pn则是植物光合机构功能效率和初级生产力高低的指标[40]。本研究中,在一定光强范围内入侵植物豚草和三裂叶豚草Pn、Gs、WUE的增幅明显高于本地种紫苑,说明与共生本地植物相比,两种入侵植物的气孔伸展范围和灵活性更大,可以根据环境条件的变化进行调节,从而保证在水分损失较少的情况下能够同化更多的CO2。WUE是植物水分生理的一个重要指标,高WUE对处在干旱环境中的植物非常重要[26]。随着有效光合辐射的增加,入侵植物的Gs也随之增加,然而并没有导致较低的WUE,可能因为Pnmax的增加弥补了由于Gs的增加而导致的水分散失[41],并且豚草和三裂叶豚草WUE的最大值均高于紫苑,也显示出两者有更高的水分利用效率。与本研究结果相同,Feng等[42]和梁作盼等[43]研究表明,入侵植物紫茎泽兰与本地种相比有更高的WUE;王晓红等[15]的研究也表明入侵植物小飞蓬水分的利用效率显著高于其伴生种山苦荬和鸭跖草。
3.2 两种菊科入侵植物与本地种叶片特性指标之间的差异
植物吸收光能同化CO2主要是在叶绿体内完成的,所以叶绿素含量直接影响植物光合作用[44]。本研究中,豚草和三裂叶豚草叶绿素含量均显著高于本地种紫苑。SLA与氮素的分配紧密相关[45],王满莲等[46]研究表明,高氮促进了入侵植物光合能力的提高,利于碳积累;磷素又可促进植物开花结实,在细胞膜结构、物质代谢、酶活性调节以及信号传导等方面都起着极为重要的作用,在植物的光合作用中同样起着极其重要的作用[47],较高的叶片氮磷含量不仅可以促进植物生长,而且还能增强植物光合能力[45]。本研究中,豚草和三裂叶豚草叶片N、P含量均显著高于本地植物紫苑,则可能显示其对土壤中N、P养分利用能力更强[37]。
Feng等[42]也指出,入侵植物相比原产地种群有着更高的Nmass和SLA,这可能是因为入侵种为了逃避自然天敌而出现的基因定向选择,当外来种入侵到新的地区时,自然天敌相对于原始产地减少,于是入侵种就会将较少的氮素分配到植物细胞壁中(合成防御物质),进而将较多的氮素分配到光合作用中(促进植物生长),从而增强自身的竞争力。本研究中,豚草和三裂叶豚草SLA均显著高于本地植物紫苑;同时,豚草和三裂叶豚草叶片的Nmass、Pmass均高于紫苑,且差异显著;CCmass、PEUE和PNUE作为植物能量利用效率的重要指标,反映了植物适应环境的能量策略[33]。其中,CCmass是衡量植物叶片建成所需能量的重要指标,反映了植物叶片的能量利用策略,较低的CCmass可能会增加植物的竞争优势[32]。本研究中,豚草和三裂叶豚草较高的Pnmax并没有导致较低的CCmass,原因可能是植物叶片较高的含氮量以及合成蛋白质、氨基酸等大分子物质的高消耗,会导致叶片CCmass的增加[42],这与Geng等[41]的研究结果也一致。PNUE是植物氮素生理利用效率的特征之一[48]。PEUE则是反映植物能量利用效率最直接的指标,入侵植物的PNUE和PEUE均显著高于本地种,与Funk等[49]的研究结果相一致。
光合作用过程中最重要的指标是Pn,其大小决定着物质积累能力的高低,在一定程度上也决定着植物生长的快慢和在群落中竞争能力的强弱[50]。本研究中,豚草和三裂叶豚草较高的植物Pn并没有导致较低的CCmass,这与Penning等[33]、Feng等[42]的结果一致,这可能是因为入侵植物的PEUE和PNUE显著高于本地种,而PEUE和PNUE是表征植物叶片能量利用效率的重要指标[51],导致入侵植物较高的CCmass,从而前者在表征植物资源获取能力方面更具参考价值。
本研究表明,入侵植物豚草、三裂叶豚草及当地紫菀的净光合速率均与有效光合辐射极显著相关。3种植物的光饱和点均高于800 μmol·m-2·s-1,且表现为三裂叶豚草>豚草>紫菀;光补偿点顺序为紫菀>三裂叶豚草>豚草,本地种紫菀显著高于豚草;两种入侵植物的表观量子效率均显著低于紫菀;3种植物的Pnmax顺序为豚草>三裂叶豚草>紫菀。两种入侵植物的水分利用效率随有效光合辐射增加而增加幅度明显高于本地种紫菀。总之,相同条件下,两种入侵植物较当地植物显示出了较高光合能力、光能利用率和水分利用效率。
References
[1]鞠瑞亭,李慧,石正人,等.近十年中国生物入侵研究进展[J].生物多样性,2012,20(5):581–611 Ju R T,Li H,Shi Z R,et al.Progress of biological invasions research in China over the last decade[J].Biodiversity Science, 2012,20(5):581–611
[2]Westpha M I,Browne M,MacKinnon K,et al.The link between international trade and the global distribution of invasive alien species[J].Biological Invasions,2008,10(4):391–398
[3]徐高峰,申时才,张付斗,等.外来入侵植物小子虉草研究进展与展望[J].中国生态农业学报,2015,23(9):1083–1092 Xu G F,Shen S C,Zhang F D,et al.Research progress and prospect about exotic invasive speciesPhalaris minorRetz[J].Chinese Journal of Eco-Agriculture,2015,23(9):1083–1092
[4]Pattison R R,Goldstein G,Ares A.Growth,biomass allocation and photosynthesis of invasive and native Hawaiian rainforest species[J].Oecologia,1998,117(4):449–459
[5]McDowell S C L.Photosynthetic characteristics of invasive and noninvasive species ofRubus(Rosaceae)[J].American Journal of Botany,2002,89(9):1431–1438
[6]Durand L Z,Goldstein G.Photosynthesis,photoinhibition,and nitrogen use efficiency in native and invasive tree ferns in Hawaii[J].Oecologia,2001,126(3):345–354
[7]Feng Y L,Fu G L.Nitrogen allocation,partitioning and use efficiency in three invasive plant species in comparison with their native congeners[J].Biological Invasions,2008,10(6):891–902
[8]Matzek V.Superior performance and nutrient-use efficiency of invasive plants over non-invasive congeners in a resource-limited environment[J].Biological Invasions,2011,13(12):3005–3014
[9]Smith M D,Knapp A K.Physiological and morphological traits of exotic,invasive exotic,and native plant species in tallgrass prairie[J].International Journal of Plant Sciences,2001,162(4):785–792
[10]Daehler C C.Performance comparisons of co-occurring native and alien invasive plants:Implications for conservation and restoration[J].Annual Review of Ecology,Evolution,and Systematics,2003,34:183–211
[11]Baruch Z,Goldstein G.Leaf construction cost,nutrient concentration,and net CO2assimilation of native and invasive species in Hawaii[J].Oecologia,1999,121(2):183–192
[12]Alpert P,Bone E,Holzapfel C.Invasiveness,invasibility and the role of environmental stress in the spread of non-native plants[J].Perspectives in Plant Ecology,Evolution and Systematics,2000,3(1):52–66
[13]王睿芳,冯玉龙.叶物候、构建消耗和偿还时间对入侵植物碳积累的影响[J].生态学报,2009,29(5):2568–2577 Wang R F,Feng Y L.The effects of leaf phenology,construction cost and payback time on carbon accumulation in invasive plants[J].Acta Ecologica Sinica,2009,29(5):2568–2577
[14]王俊峰,冯玉龙,李志.飞机草和兰花菊三七光合作用对生长光强的适应[J].植物生理与分子生物学学报,2003,29(6):542–548 Wang J F,Feng Y L,Li Z.Acclimation of photosynthesis to growth light intensity inChromolaena odorataL.andGynurasp.[J].Journal of Plant Physiology and Molecular Biology,2003,29(6):542–548
[15]王晓红,纪明山.入侵植物小飞蓬及其伴生植物的光合特性[J].应用生态学报,2013,24(1):71–77 Wang X H,Ji M S.Photosynthetic characteristics of an invasive plantConyza canadensisand its associated plants[J].Chinese Journal of Applied Ecology,2013,24(1):71–77
[16]万方浩,郭建英,张峰,等.中国生物入侵研究[M].北京:科学出版社,2009 Wan F H,Guo J Y,Zhang F,et al.Research on Biological Invasions in China[M].Beijing:Science Press,2009
[17]Friedman J,Barrett S C H.High outcrossing in the annual colonizing speciesAmbrosia artemisiifolia(Asteraceae)[J].Annals of Botany,2008,101(9):1303–1309
[18]Mandrioli P,Di Cecco M,Andina G.Ragweed pollen:The aeroallergen is spreading in Italy[J].Aerobiologia,1998,14(1):13–20
[19]王大力,祝心如.三裂叶豚草的化感作用研究[J].植物生态学报,1996,20(4):330–337 Wang D L,Zhu X R.Allelopathic research ofAmbrosia tri-fida[J].Acta Phytoecologica Sinica,1996,20(4):330–337
[20]沙伟,周福军,祖元刚.不同生境三裂叶豚草种群的遗传结构[J].植物研究,2000,20(1):94–98 Sha W,Zhou F J,Zu Y G.The population genetic structure ofAmbrosia trifidain different environment[J].Bulletin of Botanical Research,2000,20(1):94–98
[21]魏守辉,曲哲,张朝贤,等.外来入侵物种三裂叶豚草(Ambrosia trifidaL.)及其风险分析[J].植物保护,2006,32(4):14–19 Wei S H,Qu Z,Zhang C X,et al.Invasive alien species giant ragweed (Ambrosia trifidaL.) and its risk assessment[J].Plant Protection,2006,32(4):14–19
[22]吴彦琼,胡玉佳.外来植物南美蟛蜞菊、裂叶牵牛和五爪金龙的光合特性[J].生态学报,2004,24(10):2334–2339 Wu Y Q,Hu Y J.Researches on photosynthetic characteristics of exotic plantsWedelia trilobata,Pharbitis nilandIpomoea cairica[J].Acta Ecologica Sinica,2004,24(10):2334–2339
[23]陈新微,魏子上,刘红梅,等.云南菊科入侵物种与本地共生物种光合特性比较[J].环境科学研究,2016,29(4):538–546 Chen X W,Wei Z S,Liu H M,et al.Comparison of photosynthetic characteristics between invasive and co-occuring native Asteraceae plants in Yunnan Province,China[J].Research of Environmental Sciences,2016,29(4):538–546
[24]Ye Z P.A new model for relationship between irradiance and the rate of photosynthesis inOryza sativa[J].Photosynthetica,2007,45(4):637–640
[25]叶子飘,于强.光合作用光响应模型的比较[J].植物生态学报,2008,32(6):1356–1361 Ye Z P,Yu Q.Comparison of new and several classical models of photosynthesis in response to irradiance[J].Journal of Plant Ecology,2008,32(6):1356–1361
[26]高丽,杨劼,刘瑞香.不同土壤水分条件下中国沙棘雌雄株光合作用、蒸腾作用及水分利用效率特征[J].生态学报,2009,29(11):6025–6034 Gao L,Yang J,Liu R X.Effects of soil moisture levels on photosynthesis,transpiration,and moisture use efficiency of female and male plants ofHippophae rhamnoidesssp.sinensis[J].Acta Ecologica Sinica,2009,29(11):6025–6034
[27]舒展,张晓素,陈娟,等.叶绿素含量测定的简化[J].植物生理学通讯,2010,46(4):399–402 Shu Z,Zhang X S,Chen J,et al.The simplification of chlorophyll content measurement[J].Plant Physiology Communications,2010,46(4):399–402
[28]顾大形,陈双林,黄玉清.土壤氮磷对四季竹叶片氮磷化学计量特征和叶绿素含量的影响[J].植物生态学报,2011,35(12):1219–1225 Gu D X,Chen S L,Huang Y Q.Effects of soil nitrogen and phosphonium on leaf nitrogen and phosphonium stoichiometric characteristics and chlorophyll content ofOligostachyum lubricum[J].Chinese Journal of Plant Ecology,2011,35(12):1219–1225
[29]Sparks D L,Page A L,Helmke P A,et al.Methods of Soil Analysis,Part 3:Chemical Methods[M].Madison:Soil Science Society of America,American Society of Agronomy,1996
[30]鲁如坤.土壤农业化学分析方法[M].北京:中国农业科学出版社,2000:296–338 Lu R K.Methods of Soil Agricultural Chemistry Analysis[M].Beijing:Chinese Agricultural Science and Technology Press,2000:296–338
[31]Williams K,Percival F,Merino J,et al.Estimation of tissue construction cost from heat of combustion and organic nitrogen content[J].Plant,Cell &Environment,1987,10(9):725–734
[32]屠臣阳,皇甫超河,姜娜,等.入侵植物黄顶菊与5种共生植物叶片建成成本的比较[J].生态学杂志,2013,32(11):2985–2991 Tu C Y,Huangfu C H,Jiang N,et al.Comparison of leaf construction cost between invasive plantFlaveria bidentisand its five co-occuring plants[J].Chinese Journal of Ecology,2013,32(11):2985–2991
[33]Penning De Vries F W T,Brunsting A H M,Van Laar H H.Products,requirements and efficiency of biosynthesis:A quantitative approach[J].Journal of Theoretical Biology,1974,45(2):339–377
[34]Jäger H,Alencastro M J,Kaupenjohann M,et al.Ecosystem changes in Galápagos highlands by the invasive treeCinchona pubescens[J].Plant and Soil,2013,371(1/2):629–640
[35]Evans R D,Rimer R,Sperry L,et al.Exotic plant invasion alters nitrogen dynamics in an arid grassland[J].Ecological Applications,2001,11(5):1301–1310
[36]Poorter L.Light-dependent changes in biomass allocation and their importance for growth of rain forest tree species[J].Functional Ecology,2001,15(1):113–123
[37]Ehrenfeld J G.Effects of exotic plant invasions on soil nutrient cycling processes[J].Ecosystems,2003,6(6):503–523
[38]张教林,曹坤芳.光照对两种热带雨林树种幼苗光合能力、热耗散和抗氧化系统的影响[J].植物生态学报,2002,26(6):639–646 Zhang J L,Cao K F.The effect of irradiance on photosynthetic capacity,heat dissipation,and antioxidants of seedlings of two tropical rain forest tree species[J].Acta Phytoecologica Sinica,2002,26(6):639–646
[39]Chown S L,Slabber S,McGeoch M A,et al.Phenotypic plasticity mediates climate change responses among invasive and indigenous arthropods[J].Proceedings of the Royal Society B:Biological Sciences,2007,274(1625):2531–2537
[40]冯玉龙,曹坤芳,冯志立,等.四种热带雨林树种幼苗比叶重,光合特性和暗呼吸对生长光环境的适应[J].生态学报,2002,22(6):901–910 Feng Y L,Cao K F,Feng Z L,et al.Acclimation of lamina mass per unit area,photosynthetic characteristics and dark respiration to growth light regimes in four tropical rainforest species[J].Acta Ecologica Sinica,2002,22(6):901–910
[41]Geng X Y,Jiang S,Li B,et al.Do higher resource capture ability and utilization efficiency facilitate the successful invasion of exotic plant? A case study ofAlternanthera philoxeroides[J].American Journal of Plant Sciences,2013,4(9):1839–1845
[42]Feng Y L,Li Y P,Wang R F,et al.A quicker return energy-use strategy by populations of a subtropical invader in the non-native range:A potential mechanism for the evolution of increased competitive ability[J].Journal of Ecology,2011,99(5):1116–1123
[43]梁作盼,李立青,万方浩,等.土壤微生物对紫茎泽兰生长与竞争的反馈:不同灭菌方法的比较[J].中国生态农业学报,2016,24(9):1223–1230 Liang Z P,Li L Q,Wan F H,et al.Feedback of soil biota onAgeratina adenophoragrowth and competitiveness with native plant:A comparison of different sterilization methods[J].Chinese Journal of Eco-Agriculture,2016,24(9):1223–1230
[44]叶子飘,赵则海.遮光对三叶鬼针草光合作用和叶绿素含量的影响[J].生态学杂志,2009,28(1):19–22 Ye Z P,Zhao Z H.Effects of shading on the photosynthesis and chlorophyll content ofBidens pilosa[J].Chinese Journal of Ecology,2009,28(1):19–22
[45]FengY L,Fu G L,Zheng Y L.Specific leaf area relates to the differences in leaf construction cost,photosynthesis,nitrogen allocation,and use efficiencies between invasive and noninvasive alien congeners[J].Planta,2008,228(3):383–390
[46]王满莲,冯玉龙.紫茎泽兰和飞机草的形态、生物量分配和光合特性对氮营养的响应[J].植物生态学报,2005,29(5):697–705 Wang M L,Feng Y L.Effects of soil nitrogen levels on morphology,biomass allocation and photosynthesis inAgeratina adenophoraandChromoleana odorata[J].Acta Phytoecologica Sinica,2005,29(5):697–705
[47]李志刚,谢甫绨,张玉铃,等.磷胁迫对大豆不同磷素基因型光合作用的影响[J].内蒙古民族大学学报:自然科学版,2004,19(3):297–299 Li Z G,Xie P D,Zhang Y L,et al.Influence to photosynthesis of different genotype soybean with phosphorus stress[J].Journal of Inner Mongolia University for Nationalities,2004,19(3):297–299
[48]李勇.氮素营养对水稻光合作用与光合氮素利用率的影响机制研究[D].南京:南京农业大学,2011 Li Y.Studies on mechanisms of effects of different nitrogen supplies on photosynthesis and photosynthetic nitrogen use efficiency of rice plants[D].Nanjing:Nanjing Agricultural University,2011
[49]Funk J L,Vitousek P M.Resource-use efficiency and plant invasion in low-resource systems[J].Nature,2007,446(7139):1079–1081
[50]王康满,侯元同.山东归化植物一新记录属——银胶菊属[J].曲阜师范大学学报,2004,30(1):83–84 Wang K M,Hou Y T.PartheniumL (Asteraceae) a newly naturalized record genus in Shandong Province[J].Journal of Qufu Normal University,2004,30(1):83–84
[51]Meziane D,Shipley B.Direct and indirect relationships between specific leaf area,leaf nitrogen and leaf gas exchange:Effects of irradiance and nutrient supply[J].Annals of Botany,2001,88(5):915–927
Comparison of gas exchange characteristics between two invasive plants and a co-occurring native plant in Liaoning Province*
WEI Zishang1,2,CHEN Xinwei1,2,YANG Dianlin1,HUANGFU Chaohe1,2**
(1.Agro-Environmental Protection Institute,Ministry of Agriculture,Tianjin 300191,China;2.College of Plant Protection,Shenyang Agriculture University,Shenyang 110866,China)
Biological invasion is one of the century’s most serious environmental problems.A comparative study of invaders and co-occurring natives is an effective approach to gaining insights into the invasiveness of exotic plants.Ambrosia artemisiifoliaL.andA.trifidaL.are two aggressive North American annual weeds in China,whereasA.trifidaoccurs mainly in Northeast China.They usually invade cultivated fields and reduce crop productivity,causing significant ecological and socio-economic impacts in introduced areas.Besides its economic impact on crop yield,A.artemisiifoliaalso represents a major health problem since its pollen is a strong allergen and one of the main causes of hay fever.To explore the traits contributing to invasiveness ofA.artemisiifoliaandA.trifidaand to test the relationship between plasticity of these traits and invasiveness,we conducted a comparative study of two invasive species and their co-occurring native Asteraceae plant,Aster tataricus,intataricus,in terms of gas exchange and efficiencies in resource utilization.We tested the hypothesis thatA.artemisiifoliaandA.trifidahad growth-related ecophysiological advantages over the natives in its non-native range,which resulted in its rapid growth and enhanced its invasiveness.Photosynthesis,specific leaf area (SLA),and the efficiency of resource use (light,phosphorus,and nitrogen) were examined in field.Leaf construction cost (CC) is considered as a quantifiable measure of energy demand for biomass production and is related to energy use efficiency as it includes component of both morphological and physiological traits.The results showed that the light saturation point (LSP) ofA.artemisiifolia,A.trifidawere significantly higher than that of the native speciesA.tataricus.The order ofPnmaxwas thatA.artemisiifolia>A.trifida>A.tataricus,and this parameter of two invasive species were significantly higher than that ofA.tataricusby 151.28%,82.80%,respectively.The specific leaf area (SLA),leaf N content per unit mass (Nmass),leaf P content per unit mass (Pmass),photosynthetic energy use efficiency (PEUE),photosynthetic nitrogen use efficiency (PNUE) of two invasive plants were significantly higher than those of the native,but the difference of leaf construction cost per unit mass (CCmass) between the three plant species was not significant.In summary,the two Asteraceae invasive plant species has higher gas exchange characteristics and leaf traits as compared with co-family non-invasive plant species,and their photosynthetic characteristics and leaf characteristics were closely linked.The two Asteraceae invasive plant species had higher photosynthetic characteristics and energy use parameters,includingPnmax,PNUE,PEUE,water use efficiency (WUE),leading to more effectively resources capturing and utilization efficiency,which may be one of the reasons for successful invasion of these Asteraceae invasive plants in the studied area.
Asteraceae invasive plants;Gas exchange characteristics;Maximum net photosynthetic rate;Leaf traits;Resources utilization efficiency
Dec.30,2016;accepted Mar.22,2017
X171.1
:A
:1671-3990(2017)07-0975-08
10.13930/j.cnki.cjea.161190
魏子上,陈新微,杨殿林,皇甫超河.辽宁地区两种菊科入侵植物与本地植物光合特性比较[J].中国生态农业学报,2017,25(7):975-982
Wei Z S,Chen X W,Yang D L,Huangfu C H.Comparison of gas exchange characteristics between two invasive plants and a co-occurring native plant in Liaoning Province[J].Chinese Journal of Eco-Agriculture,2017,25(7):975-982
*天津市自然科学基金项目(12JCQNJC09800)资助
** 通讯作者:皇甫超河,主要从事入侵生态学研究。E-mail:huangfu24@163.com
魏子上,研究方向为入侵生态学。E-mail:602796418@qq.com
2016-12-30 接受日期:2017-03-22
*This study was supported by the Natural Science Foundation of Tianjin (12JCQNJC09800).
** Corresponding author,E-mail:huangfu24@163.com
