入侵植物黄顶菊的繁殖生物学特性
2017-07-15魏珍岳彩鹏常岳旸李秋洁许
魏珍++岳彩鹏++常岳旸++李秋洁++许雪敏++朱世新

摘要:黄顶菊[Flaveria bidentis (L.) Kuntze]是近年来恶性入侵我国的一种菊科植物。本研究通过探讨黄顶菊的繁殖生物学特性,為科学预防和治理黄顶菊的入侵扩散提供理论基础。于2013年8月至2015年11月,对在郑州大学生命科学学院栽培的黄顶菊种群进行繁殖生物学特征研究,包括综合形态学、花序开花动态、花粉胚珠比和交配系统类型等,并对河南省郑州市东风渠附近的黄顶菊自然种群的访花昆虫进行观察研究。结果表明,黄顶菊具有灵活的交配系统,有性繁殖方式为兼性异交型,即自交和异交授粉均能产生可育瘦果,也可通过无融合生殖方式产生瘦果。瘦果体积小[千粒质量仅为(0.17±0.01) g]且数量庞大,发育成熟时间短,开花5 d后便可观察到成熟的瘦果。在开花盛期,观察到访花昆虫有蝶类、蝇类、蛾类和蜂类等。本研究从繁殖生物学的角度揭示黄顶菊的快速入侵机制,为控制和治理黄顶菊由河南省向南扩散蔓延提供了重要理论依据。
关键词:菊科;交配系统;黄顶菊;入侵物种;花粉胚珠比;繁殖生物学
中图分类号: S451;Q945文献标志码: A
文章编号:1002-1302(2017)09-0097-04
黄顶菊[Flaveria bidentis (L.) Kuntze]原产于南美洲,在分类上隶属菊科(Asteraceae)堆心菊族(Helenieae)黄顶菊属(Flaveria)[1]。刘全儒曾将黄顶菊定名为二齿黄菊[2],《中国植物志(英文版)》将它译为“黄顶菊”,并归属为向日葵族(Heliantheae)[3]。2001年,在南开大学附近路边首次发现黄顶菊,虽然入侵我国的时间较短,但表现出非常强的入侵性,在入侵等级上属于1级,即恶性入侵类[1,4]。目前黄顶菊在河南省的黄河以及沿岸地区(包括安阳市、濮阳市、新乡市、郑州市等)的入侵比较严重,已经对该地区造成了一定规模的生态破坏,而且蔓延形势相当严峻。有研究预测,黄顶菊在我国的潜在适生区为南方诸省[5],而河南省是黄顶菊从北向南蔓延的必经区域和关键区域。如果黄顶菊的蔓延得不到控制,不仅会对入侵地的粮食生产、生物多样性保护和生态环境等均造成严重的危害,也会加剧黄顶菊向南方入侵的速度。
菊科植物是我国入侵种类最多的类群之一,多数具有生长发育快,果实数量大且有适宜传播扩散的结构和机制等生物学特性,有些植物还能分泌特殊物质抑制其他植物的生长[6]。对黄色星蓟(Centaurea solstitialis L.)、窄叶千里光(Senecio inaequidens)和三叶鬼针草(Bidens pilosa L.)[7-9]等菊科入侵植物的繁殖生物学研究表明,菊科入侵物种呈现出多样化的繁殖类型,如营养繁殖、无融合生殖、自交不亲和、自交亲和以及不完全自交亲和等。与繁殖有关的特征,在外来入侵植物种群建群时将直接影响其归化或入侵是否成功[10],因此研究入侵物种的繁殖生物学特性对控制种群扩散分布具有重要意义。本研究通过探讨黄顶菊的综合形态学特征及花序开花动态、交配系统类型和访花昆虫等特征,鉴定黄顶菊的交配系统类型并评估其繁殖能力,为科学预测、预防和治理黄顶菊的入侵工作提供更多的理论基础。
1材料与方法
1.1试验材料
黄顶菊种子采集于河南省郑州市金水区东风渠畔的黄顶菊自然种群,来源于不同植株上的成熟种子经过干燥后保存在室温条件下。于2014年4月和2015年4月,随机选取收集的黄顶菊种子,在花盆中萌发,每个花盆保留1株黄顶菊植株,共计30株。栽培的黄顶菊植株置于郑州大学生命科学学院实验楼阳台,进行开放培育。
1.2试验方法
1.2.1黄顶菊综合形态学特征及花序开花动态观察
待黄顶菊植株进入花期,对形态特征如花序、小花、苞片、瘦果等性状进行观察记录,并对花序、小花、瘦果数目、瘦果质量及总苞片、瘦果长度进行测量。使用Nikon SMZ800体视显微镜对黄顶菊瘦果形态进行观察,瘦果的成熟以果体饱满,颜色变为黑褐色为标准。在Nikon ECLIPSE E200显微镜下观察黄顶菊小花柱头可授区的位置和花粉在柱头上的落置情况,并使用MiE 3.0图像处理软件进行拍照保存。
黄顶菊花序的开花动态研究参照郝建华等的方法[9,11]并加以修改:以黄顶菊整个聚伞花序为观察单位,以聚伞花序中央头状花序开放为开花的起始阶段,以最外一轮的头状花序开放作为开花期的结束,对黄顶菊从开花到结果期间的形态变化和发育所经历的时间进行观察和记录。
1.2.2人工控制套袋试验
在黄顶菊开花前期,在30株植株上各选取一定数量的形态发育一致的花序分别进行3种处理:(a)自然状况的花序作开放处理;(b)检测黄顶菊是否自交亲和,在开花前将整个花序用牛皮纸进行套袋处理;(c)参照Koltunow等的方法[12]将开花前的花序中的所有花药和柱头去除,仅保留部分花柱和子房,进行去雄处理,然后套袋。每个黄顶菊植株上作15组处理,3种处理各重复5次。等瘦果成熟后,观察各处理的单花序的结实现象,并对结实率进行统计分析。
1.2.3花粉胚珠比
Cruden认为,花粉胚珠比(pollen ∶ovule ratio,简称P/O)是植物繁育系统的重要评估指标[13]。从20个黄顶菊植株上随机选取40个花序,用FAA固定液固定。从每个固定过的花序上随机选取10枚花药未开裂的小花,参照郝建华等的方法[9,14],将所有的花药进行处理并制成压片在Nikon ECLIPSE E200显微镜下观察,并使用MiE 3.0图像处理软件进行拍照保存,统计总花粉量和每枚小花的P/O值。
1.2.4访花昆虫
在黄顶菊的开花盛期,对位于郑州市金水区东风渠畔的黄顶菊自然种群和盆栽黄顶菊的访花昆虫进行观察,并捕捉正在访花的昆虫,带回室内制作成标本,对标本进行鉴定并保存。
1.2.5数据分析
采用SPSS 21.0统计软件分析数据,并对不同处理下的结实率进行单因素ANOVA分析,不同处理间使用LSD法进行多重比较,显著性水平为0.05,最后用Excel软件绘制图表。
2结果与分析
2.1黄顶菊综合形态学特征及花序开花动态观察
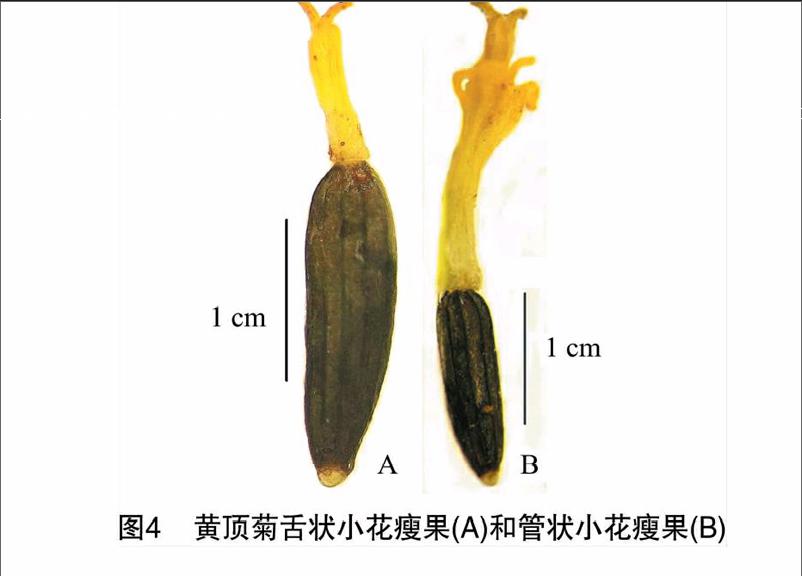
黄顶菊植株高10~120 cm,根系发达。茎直立,具纵棱,被微绒毛,呈紫红色或绿色,具主轴和多级分枝组成,每级侧枝两两对生。叶长椭圆形至披针状椭圆形,基生三出脉,交互对生,叶缘具浅锯齿,葉柄基部近于合生,长5~18 cm,宽 2.5~4.0 cm。在河南省黄顶菊的花期主要为6月下旬至10月中旬,头状花序密集成多歧聚伞花序(图1)或平顶形伞房状花序,具3~4级侧枝。每株具有26~70(43.5±13.38)个多歧聚伞花序,每一聚伞花序包含3~10(4.58±0.77)个蝎尾状花序,每一个蝎尾状花序由4~20(8.09±0.75)个头状花序组成,头状花序排列在同一平面上,每一个头状花序中有2~11枚小花。总苞长椭圆形,内凹,先端圆或钝,3~4枚总苞片,长约(3.90±0.49)mm,黄绿色,内凹,小苞片1~2枚。头状花序中边缘花舌状1~2枚,黄白色,舌片不突出或微突出于闭合的小苞片外。盘花花冠檐部5裂,辐射对称,鲜黄色,基部联合成筒状,花冠筒状基部周围有白色近透明冠毛。黄顶菊的柱头呈鸭舌状二裂(图2),柱头顶端有乳突分布,两柱头分叉处内侧光滑为花粉可授区(图3)。开花时头状花序中边缘舌状花先开花,然后中央管状花开花,但整个蝎尾状花序和多歧聚伞花序按从里向外顺序开花,每天约1~8个头状花序中有小花开放,开花盛期可达11个。1个蝎尾状花序全部开放的花期约为10~20 d。从第10天开始,小花从蝎尾状花序里侧开始逐渐干萎,花冠逐渐脱落,柱头随之脱落,整个聚伞花序全部干萎需要约14~20 d。开花第5天,便可以观察到成熟瘦果。自然开放状态下单个头状花序可以产生 (6.15±2.82)粒饱满的瘦果。瘦果呈黑色或深褐色,倒披针形或近棒状,长约(2.20±0.31) mm,无喙,具10纵肋,千粒质量为(0.17±0.01) g。边缘舌状小花的瘦果(图4-A) 成熟较早且大于中央管状小花的瘦果(图4-B)。
2.2人工控制套袋试验
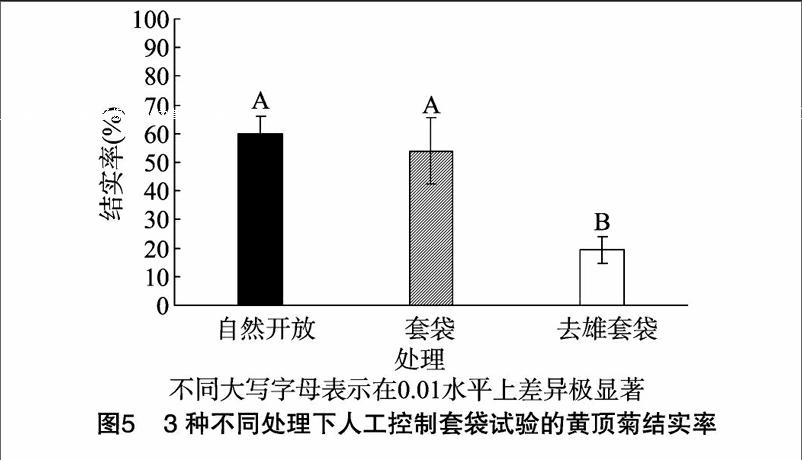
由图5可知,3种处理均可以正常结实,自然开放处理的单个花序结实率为(60.00±6.24)%;套袋处理的单个花序结实率为(54.00±11.64)%;去雄套袋单个花序结实率为(19.33±4.94)%。用SPSS 21.0软件对结实率进行单因素方差分析并进行多重比较。结果表明,3种处理之间存在极显著差异(α=0.05,df1=2,df2=12,F=36.35,P<0.01),且去雄套袋处理与其他2种处理之间差异极显著(P<0.01),而自然开放处理和套袋处理间无显著差异。
2.3花粉胚珠比
试验测得黄顶菊每枚小花的花粉数量平均为(1 105±38)粒。由于黄顶菊每个雌蕊仅含1枚胚珠,可得黄顶菊花粉胚珠比(P/O)为1 105±38。Cruden认为花粉胚珠比为介于244.7~2 588.0时,繁育系统为兼性异交[13]。因此黄顶菊的繁育系统为兼性异交型。
2.4访花昆虫
在黄顶菊的开花盛期,观察到访花昆虫有甜菜白带野螟(Spoladeare curvalis Fabricius)(图6-A)、灰蝶科(Lycaenidae)昆虫(图6-B)、蜜蜂(图6-C)、泥蜂(图6-D)、寄蝇科(Tachinidae)昆虫(图6-E)及丽蝇科(Calliphoridae)昆虫(图6-F)等蛾类、蝶类、蜂类、蝇类昆虫。
3讨论
3.1黄顶菊在入侵地的综合形态及开花动态变化
本研究对黄顶菊的综合形态学特征进行观察,结果表明,每株黄顶菊中包含的头状花序数量(以均值计算为43.5×4.58×8.09=1 611.77)以及每个头状花序中的小花数目(2~11枚)与原产地(2~8枚)相比增多[15]。小花数目的增加在我国其他地区黄顶菊自然种群也观察到一致的结果[16]。在河南省内,黄顶菊自然种群和试验室栽培种群在四月中旬就可以观察到开花现象,主要的花期集中在6月下旬至10月中旬,表现出很强的适应性。从黄顶菊开花第5天起,便可以观察到成熟的瘦果,千粒质量为(0.17±0.01) g,与天津地区的黄顶菊瘦果(0.204 2±0.005)g的质量相比较轻[16]。河南地区的黄顶菊种群与天津、河北等地的黄顶菊种群相比[16-18],不但开花较早、花期较长,并且瘦果成熟期短,仅需 5 d 便可成熟。除上述差异以外,本研究中其他地区的黄顶菊种群综合形态特征并无明显差异。
3.2黄顶菊的交配系统
花粉胚珠比(P/O)是植物繁育系统的重要评估指标[13]。本研究中的花粉胚珠比(P/O)值为1 105±38,表明黄顶菊的繁育系统为兼性异交型。在人工套袋试验中开放和套袋处理均能正常结实,说明黄顶菊能以自花授粉和异花授粉的有性生殖方式结实,验证了花粉胚珠比试验结果。自然开放和套袋2种处理的结实率之间没有显著差异,但均极显著高于去雄套袋处理的结实率。去雄套袋处理产生了可育种子,表明黄顶菊可以无融合生殖的方式结实。
黄顶菊是虫媒花,据报道访花昆虫包括蜂类、蝶类[如红珠灰蝶(Lycaeides argyrognomon Bergstrasser)]、蛾类(如甜菜白带野螟)和蝇类等[19]。本研究同样发现了这4类访花昆虫,如甜菜白带野螟、蜜蜂、泥蜂、灰蝶科、寄蝇科及丽蝇科等昆虫。访花昆虫帮助黄顶菊提高了异花授粉的结实率,对繁殖结实具有重要意义。
Baker认为,杂草的自交亲和性或具有很强的营养繁殖能力,在杂草的长距离扩散中起重要作用[20-22]。黄顶菊在存在异交的情况下,也可通过自交亲和(或)和无融合生殖的方式来增加种子产量,成为其成功入侵的重要因素。
3.3黄顶菊适应于入侵的繁殖生物学特征
首先,黄顶菊具有灵活的交配系统,既包括有性繁殖(自交和异交都能产生可育种子),也包括无融合生殖。这种包含了自花传粉、异花传粉、无融合生殖等多种繁殖方式的交配系统有利于黄顶菊种群的快速入侵及扩散。
其次,黄顶菊的瘦果发育成熟时间短。黄顶菊在开花5 d后便可观察到成熟的瘦果,花冠仍然饱满富有水分。黄顶菊瘦果的快速发育可保证在短时间内形成可育种子,有利于快速入侵。
再次,每株黄顶菊的瘦果数量庞大。以本研究的数据为例,每株黄顶菊最多产生70个多歧聚伞花序,每1个聚伞花序至多包含10个蝎尾状花序,每1个蝎尾状花序有多达20个头状花序组成,每1个头状花序中平均产生6枚饱满可育瘦果,那么仅1株黄顶菊便可产生8.4万枚瘦果。据报道,野外的黄顶菊自然种群中单株黄顶菊可产生高达30多万粒种子[23]。
此外,黄顶菊种子的发芽率很高,在不同温度、光照以及一定范围内的盐含量、pH值、重金属含量、除草剂胁迫下均有较高的萌发率[24-27]。
本研究主要探讨了一种外来恶性入侵物种黄顶菊的繁殖生物学特性,发现它具有自交亲和性、异交亲和性、无融合生殖、瘦果发育成熟迅速且数量极多等繁殖特性。研究内容从繁殖生物学角度揭示了黄顶菊快速入侵的机制,为防控黄顶菊从河南省继续向南繁殖扩散提供了重要的理论基础。
参考文献:
[1]高贤明,唐廷贵,梁宇,等. 外来植物黄顶菊的入侵警报及防控对策[J]. 生物多样性,2004,12(2):274-279.
[2]刘全儒. 中国菊科植物一新归化属——黄菊属[J]. 植物分类学报,2005,43(2):178-180.
[3]Chen Y S,Hind D J N. Heliantheae:Flora of China[M]. Beijing:Science Press,St. Louis:Missouri Botanical Garden Press,2011:852-878.
[4]閆小玲,刘全儒,寿海洋,等. 中国外来入侵植物的等级划分与地理分布格局分析[J]. 生物多样性,2014,22(5):667-676.
[5]白艺珍,曹向锋,陈晨,等. 黄顶菊在中国的潜在适生区[J]. 应用生态学报,2009,20(10):2377-2383.
[6]朱世新,覃海宁,陈艺林. 中国菊科植物外来种概述[J]. 广西植物,2005,25(1):69-76.
[7]Sun M,Ritland K. Mating system of yellow starthistle (Centaurea solstitialis),a successful colonizer in North America[J]. Heredity,1998,80(2):225-232.
[8]Lafuma L,Maurice S. Increase in mate availability without loss of self-incompatibility in the invasive species Senecio inaequidens(Asteraceae)[J]. Oikos,2007,116(2):201-208.
[9]郝建华,刘倩倩,强胜. 菊科入侵植物三叶鬼针草的繁殖特征及其与入侵性的关系[J]. 植物学报,2009,44(6):656-665.
[10]Pyek P,Richardson D M. Traits associated with invasiveness in alien plants:where do we stand?[M]. Biological Invasions:Springer Berlin Heidelberg,2007:97-125.
[11]Burtt B L. Compositae and the study of functional evolution[J]. Transactions of the Botanical Society of Edinburgh,1961,39(2):216-232.
[12]Koltunow A M. Apomixis:embryo sacs and embryos formed without meiosis or fertilization in ovules[J]. Plant Cell,1993,5(10):1425-1437.[HJ1.7mm]
[13]Cruden R W. Pollen-ovule ratios:a conservative indicator of breeding systems in flowering plants[J]. Evolution,1977,31(1):32-46.
[14]Kliber A,Eckert C G. Sequential decline in allocation among flowers within inflorescences:proximate mechanisums and adaptive significance[J]. Ecology,2004,85(6):1675-1687.
[15]Powell A M. Systematics of Flavaria[J]. Annals of the Missouri Botanical Canden,1978,65(2):590-636.[HJ1.76mm]
[16]李香菊,王贵启,张朝贤,等. 外来植物黄顶菊的分布、特征特性及化学防除[J]. 杂草科学,2006(4):58-61.
[17]任艳萍,江莎,古松,等. 外来植物黄顶菊(Flaveria bidentis)的研究进展[J]. 热带亚热带植物学报,2008,16(4):390-396.
[18]张旭,岳强,聂志英,等. 田间黄顶菊种子成熟进度研究[J]. 中国植保导刊,2013,33(3):58-60.
[19]杜喜翠,谭万忠,孙现超. 外来入侵植物黄顶菊上昆虫种类多样性研究[J]. 西南大学学报(自然科学版),2011,33(6):1-6.
[20]安瑞军,王永忠,田迅. 外来入侵植物——少花蒺藜草研究进展[J]. 杂草科学,2015,33(1):27-31.
[21]Baker H G. Self-compatibility and establishment after long-distance dispersal[J]. Evolution,1955,9(3):347-348.
[22]钟军弟,马生健,袁长春,等. 3种菊科入侵植物白花鬼针草、胜红蓟和假臭草的种子生物学特性比较研究[J]. 杂草学报,2016,34(2):7 -11.
[23]王贺军. 河北省黄顶菊疫情发生动态与防控工作进展[J]. 中国植保导刊,2008,28(8):37-39.
[24]任艳萍,古松,江莎,等. 温度、光照和盐分对外来植物黄顶菊种子萌发的影响[J]. 云南植物研究,2008,30(4):477-484.
[25]张风娟,李继泉,徐兴友,等. 环境因子对黄顶菊种子萌发的影响[J]. 生态学报,2009,29(4):1947-1953.
[26]岳强,李瑞军,陆秀君,等. 温度对黄顶菊生长发育影响的研究[J]. 中国植保导刊,2010,30(5):15-18.
[27]乔永旭,张永平. 黄顶菊种子的萌芽试验[J]. 吉林农业科学,2015,40(1):86-90,94.
