杆状病毒ctl基因的进化分析
2017-07-15朱姗颖康启鑫武亚楠陈奕蓄
朱姗颖++康启鑫++武亚楠++陈奕蓄++王文兵++何华纲


摘要:苜蓿尺蠖核型多角体病毒(AcMNPV)芋螺毒素类似多肽(CTL)具有酚氧化酶抑制剂活性,在农业害虫的生物防治领域具有潜在的应用价值。为了揭示杆状病毒ctl基因的进化特点,对杆状病毒CTLs进行多序列比对分析和分类研究。结果表明,杆状病毒CTLs可分为Ⅰ型、Ⅱ型,2种类型的CTLs都具有相同的二硫鍵骨架,但涉及昆虫酚氧化酶抑制剂活性的酪氨酸残基在2种类型CTLs中的数目、位置明显不同;另外,Ⅰ型CTLs、Ⅱ型CTLs的理化性质也存在明显差异,疏水性氨基酸平均含量分别为29.7%、21.3%,净电荷平均值分别为-3.56、+0.507,等电点平均值分别为3.86、7.32。由此推测,Ⅰ型CTLs、Ⅱ型CTLs在功能上可能存在着重要分化。
关键词:杆状病毒;ctl基因;分类;进化
中图分类号:Q753 文献标志码: A
文章编号:1002-1302(2017)09-0022-05
芋螺毒素(conotoxin)是由海洋软体动物芋螺分泌的一类用于自卫和捕食的活性多肽,一般由10~30个氨基酸残基组成,富含半胱氨酸(C)。芋螺毒素能特异性地作用于钾、钠、钙等离子通道、细胞膜上的各种神经递质、激素的受体,从而干扰细胞或神经中的信号传递,在治疗慢性疼痛、急性疼痛、神经保护等方面具有广泛的应用前景。根据保守的信号肽序列,芋螺毒素可分为A、M、O、I、P、S、T等超家族;进一步依据其半胱氨酸骨架及药理活性,又可分为α、μ、ω、δ、Ψ、σ、λ、κ、γ等家族[1]。
1992年,Eldridge等从苜蓿尺蠖核型多角体病毒(Autographa californica nucleopolyhedrovirus,简称AcMNPV)中鉴定了1个基因,该基因编码的蛋白质序列与ω-芋螺毒素具有一定的相似性且具有相同的二硫键骨架(C1—C2—C3C4—C5—C6),因此将其命名为类芋螺毒素基因(conotoxin-like,简称ctl)[2]。但Eldridge等通过体内、体外试验都未能证明杆状病毒ctl基因的任何功能[2-3]。曹青等发现,AcMNPV CTL不仅对金黄色葡萄球菌、微球菌等具有显著的抗菌活性,还可对酚氧化酶起到抑制作用,能阻止昆虫血液的黑化[4-5]。本研究从GenBank数据库中收集已知的杆状病毒ctl基因,对其编码的CTLs进行较全面系统的进化分析,在此基础上对杆状病毒ctl基因进行分类,以期为该基因的功能研究提供理论依据。
1材料与方法
1.1基因序列和蛋白质序列的获取
以AcMNPV ctl基因(GenBank登录号:L22858)作为种子序列,在GenBank数据库中进行BLAST,获取杆状病毒ctl基因序列,使用DNAStar软件进一步推导出对应的蛋白质序列。芋螺毒素ω-MⅦA、虎纹蜘蛛毒素HWTX-X、家蝇酚氧化酶抑制剂MdPOI等抑制剂半胱氨酸结(inhibitor cystine knot,简称ICK)家族(也称knottin家族)成员的氨基酸序列从KNOTTIN数据库(http://knottin.cbs.cnrs.fr)中获得。
1.2多序列比对分析
使用SignalP 3.0软件(http://www.cbs.dtu.dk/services/SignalP-3.0)预测杆状病毒CTL蛋白质的信号肽,进而根据信号肽的位置,推导CTL成熟肽。使用ClustalW软件(http://www.genome.jp/tools/clustalw)对杆状病毒CTL成熟肽进行多序列比对分析,并采用同样的方法对ICK家族进行多序列比对分析。
1.3系统进化树的构建
应用MEGA 4.0软件通过邻接(N-J)法构建系统进化树。
1.4蛋白质理化性质分析
用DNAStar软件统计杆状病毒CTL成熟肽、ICK家族成员的疏水性氨基酸含量,并计算净电荷、等电点(pI)。
2结果与分析
2.1杆状病毒ctl基因的分布
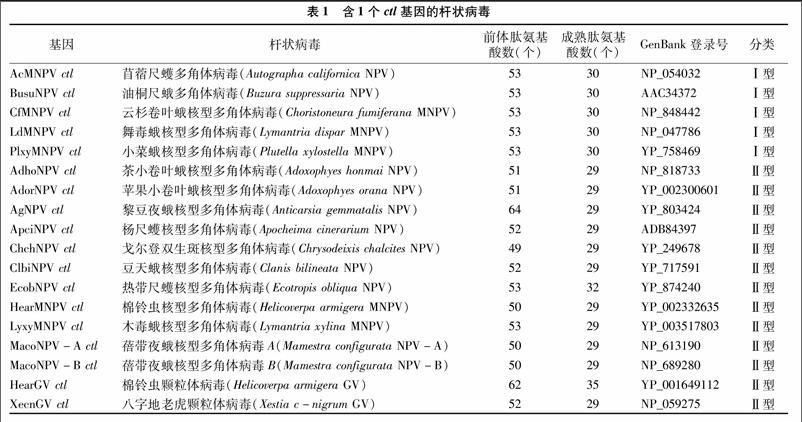
以AcMNPV ctl基因为种子,在GenBank数据库中进行BLAST分析。结果显示,在全基因组已测序的50多个杆状病毒中,有22个杆状病毒含有ctl基因,约占40%。在这些杆状病毒中,ctl基因的数目一般为1~2个。例如,AcMNPV、豆天蛾核型多角体病毒(Clanis bilineata NPV,简称ClbiNPV)等16种NPVs和棉铃虫颗粒体病毒(Helicoverpa armigera granulosisvirus,简称HearGV)、八字地老虎颗粒体病毒(Xestia c-nigrum GV,简称XecnGV)等2种GVs都只有1个ctl基因(表1);蓖麻蚕核型多角体病毒(Samia cynthia NPV,简称SacyNPV)、柞蚕核型多角体病毒(Antheraea pernyi NPV,简称AnpeNPV)等6种NPVs均有2个ctl基因(表2),而家蚕核型多角体病毒(Bombyx mori NPV,简称BmNPV)等杆状病毒中则没有该基因。在杆状病毒中,ctl基因序列一致性在74%~100%之间,推导的蛋白质CTLs的氨基酸序列一致性为 40%~78%。对不同杆状病毒CTLs进行信号肽的预测分析,发现所有CTLs的N端都有1段信号肽序列,其氨基酸组成的保守性较低,而成熟肽的氨基酸序列保守性较高。
2.2杆状病毒CTLs的分类
对预测的杆状病毒CTLs成熟肽序列进行多序列比对分析,可将杆状病毒CTLs分为2种类型,分别命名为Ⅰ型CTLs、Ⅱ型CTLs(图1)。Ⅰ型CTLs的前体肽都含有53个氨基酸残基(aa),预测的成熟肽均为30个氨基酸,成熟肽的氨基酸组成相当保守,为ACA(/S)E(/D)TGAVCV(/I)HN(/S/D)DECCSGACSPV(/I)FNYCLPQ(/E/D),包括AcMNPV等4种NPVs编码的CTLs,及SacyNPV、AnpeNPV等含有2个ctl基因的NPVs所编码的CTL-1(表1、表2)。
Ⅱ型CTLs的前体肽含有48~68个氨基酸,长度变异主要位于信号肽部分,而成熟肽大部分为29个氨基酸,个别为35、32个氨基酸,成熟肽的氨基酸组成较为保守,为CTE(/D)T(/D)GR(/K)NCK(/Q)Y(/T)S(/N)Y(/A/D/N)ECCSG(/N)ACSAA(/V/L)FG(/K)F(/Y)CLH(/K/P)R,包括ClbiNPV等11种NPVs、HearGV等2种GVs编码的CTLs及含有2个ctl基因的NPVs所编码的CTL-2(表1、表2)。
采用CTLs成熟肽的氨基酸序列对杆状病毒进行系统发育分析,发现所有编码Ⅰ型CTLs的杆状病毒聚于1支,而所有编码Ⅱ型CTLs的杆状病毒聚于另1支(图2)。
2.3Ⅰ型、Ⅱ型CTLs成熟肽的保守位点
多序列比对分析表明,在Ⅰ型CTLs、Ⅱ型CTLs的成熟肽中均含有6个高度保守的半胱氨酸残基(C),二硫键骨架为C1—C2—C3C4—C5—C6。酪氨酸(Y)是涉及昆虫酚氧化酶抑制剂活性的关键位点,因此本研究对2类CTLs中的酪氨酸的分布进行统计分析。结果显示,Ⅰ型CTLs都有1个保守的酪氨酸残基,位于C5~C6之间;在Ⅱ型CTLs中,除了茶小卷叶蛾核型多角体病毒(Adoxophyes honmai NPV,简称 AdhoNPV)、棉褐带卷蛾核型多角体病毒(Adoxophyes orana NPV,简称AdorNPV)的CTL缺乏酪氨酸残基,ChchNPV CTL、OrleNPV CTL-2只有1个酪氨酸残基之外,其余CTLs在C2~C3之间都有2个保守的酪氨酸残基。除了这2个保守的酪氨酸残基,黎豆夜蛾核型多角体病毒(Anticarsia gemmatalis NPV,简称AgNPV)、HearGV、XecnGV的CTLs还有1个酪氨酸残基,其位置与Ⅰ型CTLs中的一致。此外,在HearGV CTL、黄杉毒蛾核型多角体病毒(Orgyia pseudotsugata MNPV,简称 OpMNPV)CTL-2的C1之前还有1个额外的酪氨酸残基(图1)。
2.4Ⅰ型、Ⅱ型CTLs成熟肽的理化性质
本研究对Ⅰ型、Ⅱ型CTLs的净电荷、等电点、疏水性氨基酸含量等理化性质进行了比较分析,表3结果显示,Ⅰ型CTL在pH值7.0时净电荷范围为-4.10~-3.10(平均值为-3.56),等电点(pI)范围为3.69~3.96(平均值为 3.86),疏水性氨基酸占26.7%~30.0%(平均值为 29.7%);而Ⅱ型CTLs在pH值7.0时净电荷为-0.11~+1.89(平均值为+0.507),pI值为6.88~8.01(平均值为7.32),疏水性氨基酸占17.2%~28.6%(平均值为 21.3%)。本结果表明,杆状病毒Ⅰ型、Ⅱ型CTLs的理化性质存在明显差异,这暗示2类CTLs在功能上可能存在分化。
注:括号内数据为平均值。AcCTL为苜蓿尺蠖核型多角体病毒CTL;ClbiCTL为豆天蛾核型多角体病毒CTL;SacyCTL-1、SacyCTL-2为蓖麻蚕核型多角体病毒CTL-1、CTL-2;AhIAAI为千穗谷(Amaranthus hypochondriacus)α-淀粉酶抑制剂;GsGUR为匙羹藤(Gymnema sylvestre)甜味抑制蛋白gurmarin;MjTI1为紫茉莉(Mirabilis jalapa)胰蛋白酶抑制剂;AsAST-A为海洋生物Asteropus simplex抗菌肽;MVⅡA为芋螺毒素ω-MVⅡA;AlALO1为长臂天牛(Acrocinus longimanus)抗菌肽;MdPOI为家蝇(Musca domestica)酚氧化酶抑制剂;PaAMP1为美洲商陆(Phytolacca americana)抗菌肽;HWTX-X为虎纹蜘蛛(Ornithoctonus huwena)毒素Huwentoxin-X。
2.5杆状病毒CTLs与ICK家族的比较分析
杆状病毒CTLs属于抑制剂半胱氨酸结(inhibitor cystine knot,简称ICK)家族,多序列分析表明,杆状病毒CTLs与芋螺毒素ω-MⅦA、虎纹蜘蛛毒素HWTX-X、紫茉莉胰蛋白酶抑制剂MjTI1、匙羹藤甜味抑制蛋白GsGUR、美洲商陆抗菌肽PaAMP1、家蝇酚氧化酶抑制剂MdPOI等具有相同的二硫键骨架,但氨基酸序列一致性较低,仅为16.7%~33.3%(图3)。
在理化性质方面,AhIAAI、GsGUR、MjTI1、AsAST-A等都带负电荷,等电点分别为6.06、4.97、4.81、3.04, 疏水性氨
基酸含量分别为9.4%、25.7%、24.3%、44.4%;而ω-MⅦA、AlALO1、MdPOI、PaAMP1、HWTX-X等都帶正电荷,等电点为8.00~8.53,疏水性氨基酸含量分别为6.7%、11.8%、23.7%、21.1%、14.3%(表3)。由此可见,不同的ICK家族成员在理化性质上存在较大差异,不过杆状病毒Ⅰ型CTLs的净电荷(平均值为-3.56)、等电点(平均值为3.86)与 MjTI1、AsAST-A、GsGUR等比较接近。
3讨论与结论
抑制剂半胱氨酸结家族,又称knottin家族,该家族是由富含半胱氨酸残基的小分子蛋白质组成,广泛存在于各种动物、植物和微生物中[6]。研究发现,海洋生物芋螺中的芋螺毒素属于ICK家族。在ICK家族中,HWTX-X、Pa-AMP、MdPOI等大量成员都与芋螺毒素ω-MⅦA具有相同的二硫键骨架,即C1-C2-C3C4-C5-C6。这些类MⅦA(MⅦA-like)蛋白往往具有复杂的生物学功能,例如,HWTX-X是虎纹蜘蛛产生的一种神经毒素,在捕食过程中发挥作用[7]。PaAMP1是从美洲商陆种子中分离的抗菌肽,对多种真菌和革兰氏阳性菌具有明显的抑制活性[8]。MdPOI是从家蝇蛹中发现的酚氧化酶抑制剂(phenoloxidase inhibitor,简称POI),对家蝇幼虫的酚氧化酶具有显著的抑制作用[9-10]。从二硫键骨架看,杆状病毒CTLs也属于ICK家族中类MⅦA蛋白。杆状病毒CTLs很少带有正电荷,而带正电荷氨基酸对芋螺毒素的镇痛功能是必需的,因此,尽管最初杆状病毒CTLs因与芋螺毒素具一定的同源性而被命名,但是杆状病毒CTLs并不像芋螺毒素那样具有镇痛活性(笔者的相关研究数据未发表)。另外,曹青等已证实,AcMNPV CTL对金黄色葡萄球菌、微球菌、短小芽孢杆菌等多种临床致病菌的生长有显著抑制功能,目前尚不明确抑菌活性对于杆状病毒的生物学意义[4]。还有研究发现,AcMNPV CTL具有酚氧化酶抑制活性,能阻止昆虫血液的黑化[5]。
昆虫酚氧化酶不仅在昆虫发育过程中具有重要的生理功能,如参与表皮的鞣化、硬化、伤口愈合等,还在昆虫免疫防御反应中起着重要作用,它可使病原微生物表面产生黑色素沉淀,通过黑色素包囊反应来隔离和抵抗病原物,并在黑色素形成过程中产生醌及其衍生物,导致活性氧的积累,从而对病原物起到杀伤作用[11]。已有研究表明,昆虫酚氧化酶与黑化幼虫表现出的较强的杆状病毒抗性有关[12]。Trudeau等用 AcMNPV 感染美洲棉铃虫时发现,气管上皮细胞的感染位点上出现了酚氧化酶介导的黑化及包囊反应[13]。由此可见,昆虫酚氧化酶在宿主防御杆状病毒的过程中发挥着重要作用。昆虫体内存在内源性酚氧化酶抑制剂,能在纳摩尔(nmol)水平上高效地竞争性抑制酚氧化酶活性[9]。Cao等的研究表明,AcMNPV CTL是病毒来源的酚氧化酶抑制剂,它可能在一定程度上能抑制昆虫宿主对杆状病毒的防御反应[5]。
Daquinag等对家蝇MdPOI的氨基酸序列及蛋白质结构进行解析发现,MdPOI成熟肽由38个氨基酸残基组成,富含半胱氨酸、赖氨酸,其中6个半胱氨酸形成3对二硫键(C1—C2—C3C4—C5—C6),且在C5~C6之间的酪氨酸残基(Y)经羟化修饰为多巴残基,该位点与酚氧化酶抑制剂的竞争性抑制活性有重要关系[10]。本研究将杆状病毒CTLs分为2类,即Ⅰ型CTLs、Ⅱ型CTLs,其中Ⅰ型CTLs在C5~C6之间也都有1个保守的酪氨酸残基,目前尚不清楚该位点是否存在羟化修饰,而大部分Ⅱ型CTLs在C2~C3之间有2个保守的酪氨酸残基。另外,与Ⅰ型CTLs类似,3个Ⅱ型CTLs(AgNPV CTL、HearGV CTL和XecnrGV CTL)在C5~C6之间还有1个酪氨酸残基。目前尚不清楚Ⅱ型CTLs是否也具有酚氧化酶抑制活性。此外,在蛋白质疏水性、电荷特性等方面,Ⅱ型CTLs与Ⅰ型CTLs存在明显差异,由此推测,Ⅱ型CTLs的功能可能不同于Ⅰ型CTLs,这有待进一步研究。
参考文献:
[1]Puillandre N,Watkins M,Olivera B M. Evolution of Conus peptide genes:duplication and positive selection in the A-superfamily[J]. Journal of Molecular Evolution,2010,70(2):190-202.
[2]Eldridge R,Li Y,Miller L K. Characterization of a baculovirus gene encoding a small conotoxinlike polypeptide[J]. Journal of Virology,1992,66(11):6563-6571.
[3]盧玉蓉,冯枞棣,吴东,等. 含外源类蜗牛毒素基因的AcMNPV的虫体感染性研究[J]. 中国病毒学,2004,19(3):276-280.
[4]曹青,吴岩,刘永,等. AcMNPV类芋螺毒素多肽的抗菌活性分析[J]. 生物学杂志,2012,29(6):29-31.
[5]Cao Q,Zhu S Y,Wu Y,et al. The effect of a small conotoxin-like ctx gene from Autographa californica nuclear polyhedrosis virus (AcMNPV) on insect hemolymph melanization[J]. Polish Journal of Microbiology,2012,61(3):183-189.
[6]Gracy J,Le-Nguyen D,Gelly J C,et al. KNOTTIN:the knottin or inhibitor cystine knot scaffold in 2007[J]. Nucleic Acids Research,2008,36(SI):D314-D319.
[7]Liu Z,Dai J,Dai L,et al. Function and solution structure of Huwentoxin-X,a specific blocker of N-type calcium channels,from the Chinese bird spider Ornithoctonus huwena[J]. The Journal of Biological Chemistry,2006,281(13):8628-8635.
[8]Liu Y,Luo J,Xu C,et al. Purification,characterization,and molecular cloning of the gene of a seed-specific antimicrobial protein from pokeweed[J]. Plant Physiology,2000,122(4):1015-1024.
[9]Tsukamoto T,Ichimaru Y,Kanegae N,et al. Identification and isolation of endogenous insect phenoloxidase inhibitors[J]. Biochemical and Biophysical Research Communications,1992,184(1):86-92.
[10]Daquinag A C,Nakamura S,Takao T,et al. Primary structure of a potent endogenous dopa-containing inhibitor of phenol oxidase from Musca domestica[J]. Proceedings of the National Academy of Sciences of the United States of America,1995,92(7):2964-2968.
[11]Nappi A J,Christensen B M. Melanogenesis and associated cytotoxic reactions:applications to insect innate immunity[J]. Insect Biochem Mol Biol,2005,35(5):443-459.
[12]Reeson A F,Wilson K,Gunn A,et al. Baculovirus resistance in the noctuid Spodoptera exemptais phenotypically plastic and responds to population density[J]. Proceedings of the Royal Society B,1998,265:1787-1791.
[13]Trudeau D,Washburn J O,Volkman L E. Central role of hemocytes in Autographa californica M nucleopolyhedrovirus pathogenesis in Heliothis virescens and Helicoverpa zea[J]. Journal of Virology,2001,75(2):996-1003.
