茶园异质性栖境对狼蛛科蜘蛛群落结构的影响
2017-07-12黎健龙唐颢周波方华春
黎健龙, 唐颢, 周波, 方华春
广东省农业科学院茶叶研究所/广东省茶树资源创新利用重点实验室, 广州 510640
茶园异质性栖境对狼蛛科蜘蛛群落结构的影响
黎健龙, 唐颢, 周波, 方华春*
广东省农业科学院茶叶研究所/广东省茶树资源创新利用重点实验室, 广州 510640
茶园生态系统中蜘蛛既是重要的天敌类群, 又是改善生态系统服务的指示标志。为更好地保护与利用天敌蜘蛛进行生态防治, 对亚热带丘陵地区不同栖境茶园的狼蛛科蜘蛛进行调查, 并对其种类、年龄结构、性别比例等进行分析。结果表明:复杂栖境茶园(JZ)与简单栖境茶园(CK)间狼蛛科蜘蛛数量特征组成存在显著差异 (P<0.001); 狼蛛科蜘蛛优势种 JZ茶园有 3种, CK茶园有4种; JZ茶园雌蛛、雄蛛和幼蛛个体数分别是CK茶园的4.36倍、3.63倍和7.91倍。JZ茶园狼蛛科的年龄结构、性别比例等数量特征影响较大,总个体数量发生动态显著高于CK茶园(P<0.05)。主成分分析表明, JZ茶园明显向家福穴狼蛛Houa jiafui)、幼豹蛛(Pardosa pusiola)和旋囊脉水狼蛛(Venonia spirocysta)等群落数量分布较多的方向偏移, 而CK茶园则向数量减少方向偏移。研究表明, 基于茶园异质性栖境管理, 采用多样性种植方式来保持生态平衡, 对保护和利用狼蛛科蜘蛛来增强害虫自然控制能力有深远意义, 还有利于成为一种可持续改善茶园生态系统的管理方式。
茶园; 栖境异质性; 狼蛛科; 群落结构
1 前言
蜘蛛是农业生产中的重要天敌, 狼蛛科等地表游猎型蜘蛛是天敌组成中重要类群之一, 对环境变化敏感, 已广泛用作环境对生物多样性影响的指示生物[1-2]。栖境特征的改变会对生物多样产生重要影响, 特别是害虫暴发、天敌群落改变等[3-4]。已有研究表明, 农田周边栖境单一种植会使生物多样性减少, 导致害虫自然调控失效, 不利于生态环境保护;而传统农业实施间作套种复合体系模式, 改变栖境特征结构复杂程度, 更有利于生物多样性的保护和应用[5-6]。因此, 从栖境异质性角度出发, 选择多样性种植等管理措施来改善农田生境, 促进环境友好,增加生物多样性, 减少害虫发生, 已成为近年来国内外生物农业研究的热点[7]。
生境变化对狼蛛科影响方面, 罗媛媛等[8]通过对千岛湖陆桥岛屿上黑腹狼蛛(Lycosa coelestris)遗传多样性及其受生境片段化的影响研究, 结果表明生境片段化初期岛屿面积和形状是影响黑腹狼蛛种群遗传多样性的主要原因。张征田等[9]开展了镉污染对不同生境狼蛛的影响, 研究表明狼蛛体内镉的累积是受不同生境影响。此外, 狼蛛生物学特性受不同生境土壤重金属含量的影响, 可以作为重金属污染指示生物[10]。生物防治应用方面, 王智等[11]通过对稻田蜘蛛的生态位研究, 确定蜘蛛的生态位宽度值随害虫生态位宽度值增减而增减, 其优势种为狼蛛科。张永强等[12]对玉米蜘蛛群落结构及动态研究, 表明玉米地里狼蛛科种群占优势。汪波等[13]分析狼蛛对黑腹果蝇捕食数量的PCR检测, 表明随着狼蛛捕食数量增多, 其体内含有的黑腹果蝇DNA残留就越多。综上所述, 表明狼蛛科蜘蛛作为捕食性天敌, 既容易受生境变化影响, 又在农业生态系统中实施生物防治具有重要作用。虽然生态环境干扰和农田复合间作种植与蜘蛛群落的关系有一些报道[14-15], 但在广东茶区不施化学农药下, 针对茶园异质性栖境对狼蛛科蜘蛛群落结构影响却报道较少。本研究重点调查两种不同栖境茶园狼蛛科蜘蛛种类, 分析其群落结构、年龄大小、性别比例以及动态规律等影响, 对生物多样性保护政策的制定和措施的实施具有一定指导意义。
2 材料与方法
2.1 试验茶园栽培管理
本试验选取2个实验点, 选择同一区域内茶园,地理位置(海拔、气候等)、茶龄等均一致。包括复杂栖境茶园和简单栖境茶园。每个试验点生境类型茶园具有相同的坡面, 相同茶树品种等。该地秋冬季雨量较少, 夏季雨量充足, 年降雨量 1906 mm。茶园土壤类型均为赤红壤, 属酸性土茶园, pH 为 4.20—4.38。各个茶园茶树本身的株行距是 0.3 m×1.5 m。纬度:24°18′09 N; 经度: 113°23′05 E; 海拔: 42—46 m。
复杂栖境茶园, 茶园实施多样性种植, 四周植物丰富, 茶园生境相对较好, 种植品种为英红9号。茶园间种有台湾相思树(Acacia confusa Merr.)、阴香树(Cinnamomum burmanni)等多种景观树。其中间种台湾相思树, 树龄为 30 多年, 树高 15—20 m, 种植规格为株距×行距=8 m×24 m; 茶园遮荫度较大, 地表周年覆盖厚度为1—3 cm的凋落物, 夏季温度为28—35 ℃, 湿度为60%—90%, 光照强度为15200—44300 lux。茶园不使用除草剂, 杂草每年修剪3—4次, 茶园工作道及四周自然生长有狗牙根(Cynodon dactylon)、鬼针草(Bidens bipinnataL.)等草本植物。
简单栖境茶园, 为纯茶园, 种植品种为英红9号。茶园中心以及工作道没有间作乔木、高灌木等植物;茶园四周生境相对较差, 缺少遮荫, 地表覆盖凋落物较少, 茶树生长容易受极端天气影响; 夏季温度为29—37 ℃, 湿度为58%—82%, 光照强度为19000—75000 lux。茶园不使用除草剂, 杂草每年修剪4—5次, 自然生长有狗牙根(Cynodon dactylon)、狗尾草(Setaria viridis)、鬼针草(Bidens bipinnataL.)等草本植物。此外, 茶园边界周围皆无其他乔木树种。
2.2 标本采集鉴定
采用陷阱法: 把口径10 cm的一次性纸杯埋入土中, 纸杯中加入福尔马林溶液, 上面盖上20 cm左右的一次性纸盘(防止溶液挥发和陷阱被破坏), 每块茶园选取40个采样点(4次重复, 每个重复10个点), 记录下陷阱的地点和收集时间, 最后进行鉴定分析。
采集时间: 分两年进行, 2010年和2011年, 每年5至10月每半个月采集一次, 11月至次年4月每1个月采集一次。每次将采集到的蜘蛛标本浸泡在75%的酒精中并进行种类鉴定。
2.3 数据分析方法
物种丰富度(S): 每个样地中出现的物种数。多度(N): 个体数。
优势种(d): d= Ni /N代表1个物种(或类群)的优势度, 即第i个物种(或类群)个体数(Ni)占群落总个体数(N)的比例, 当d≥0.1时为优势种[16]。
采用微软SAS统计软件(SAS 8.0 Software, SAS nstitute Inc.)和R(ADE-4)多元数据分析软件[17]对试验数据进行整理与分析。主成分分析(Principal Components Analysis)通过在R中导入ADE-4 软件包,比较不同茶园狼蛛科蜘蛛综合特征的差异; 多元数据分析结果利用二维空间载荷图和得分图直观而形象地进行反映。
3 结果与分析
3.1 茶园狼蛛科蜘蛛种类组成
对狼蛛科蜘蛛种类及数量进行调查和分析(图1和表1), 共采集鉴定的种类有7种, 主要包括有旋囊脉水狼蛛(Venonia spirocysta)、幼豹蛛(Pardosa pusiola)、家福穴狼蛛(Houa jiafui)、沟渠豹蛛Pardosa laura)、脉娲蛛(Wadicosa venatrix)、苏门答腊豹蛛(Pardosa sumatrana)、黑腹狼蛛(Lycosa coelestris Schenkel)。从种类上看, 复杂栖境茶园(JZ)有7种; 而简单栖境茶园(CK)有6种, 缺少的种类为脉娲蛛。复杂栖境茶园(JZ)有优势种3种, 分别为幼豹蛛、旋囊脉水狼蛛和苏门答腊豹蛛, 多度为242、149和76, 分别占49.09%、30.22%和15.42%。简单栖境茶园有优势种4种, 以幼豹蛛、旋囊脉水狼蛛个体数量最多, 多度为49和28, 分别占42.24%和24.14%。与复杂栖境茶园相比, 幼豹蛛和旋囊脉水狼蛛多度比简单栖境茶园(CK)增加3.94倍和4.32倍; 说明茶园栖境实行间作树木增加生物多样性管理, 可改变蜘蛛群落组成和结构并有利于狼蛛科蜘蛛的种类、多度以及优势种的不断变化。

图1 两种茶园狼蜘科种类基本组成Fig. 1 Species composition of wolf spider in two tea plantations
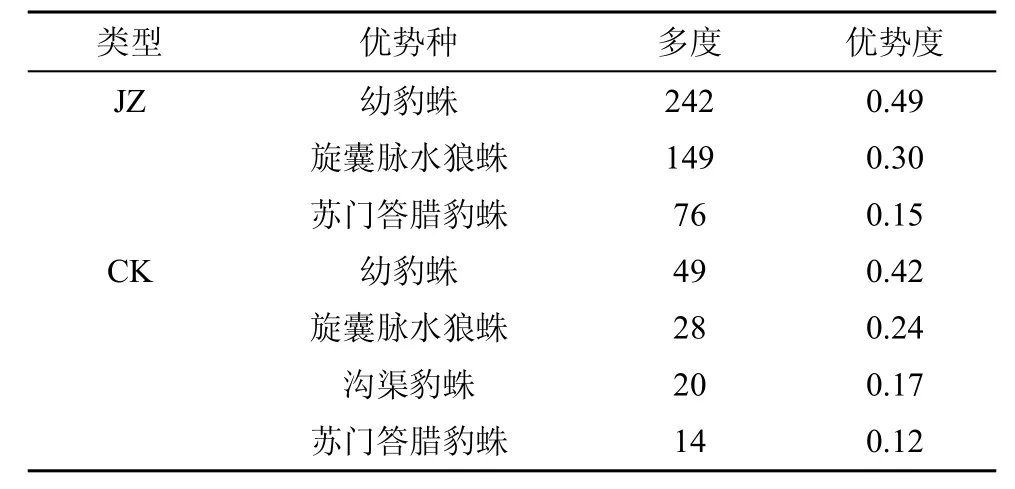
表 1 两种茶园狼蜘科蜘蛛优势种类Tab.1 Dominant species in two tea plantations
3.2 茶园狼蛛科蜘蛛性别和年龄特征
在蜘蛛群落数量特征上(如表2和表3), 不同种植模式茶园间的差异达到极显著水平(F=16.25,P<0.001) , 总体上, 复杂栖境茶园个体数量是简单栖境茶园个体数量的4.23倍; 可见, 茶园栖境复杂、间作树木对狼蛛科数量增长是有益的。两类茶园在年龄结构、性别比例方面存在显著差异(F=5.73, P<0.05)。从年龄结构上看, 复杂栖境茶园实行间作树木后幼蛛个体数增加是最多, 是简单栖境茶园个体数量的7.91倍。从性别比例上看, 雌蛛、雄蛛个体数量是简单栖境茶园个体数量的4.36倍和3.63倍。此外,雌蛛个体数中, 以旋囊脉水狼蛛的最多, 与简单栖境茶园(CK)的个体数比例为1: 0.21; 在雄蛛和幼蛛个体数中, 均以幼豹蛛的个体数最多; 幼豹蛛在复杂栖境茶园与简单栖境茶园(CK)的性比, 雄蛛比为1: 0.24, 幼蛛比为1: 0.04。说明复杂栖境茶园模式不仅影响蜘蛛群落结构数量大小, 还对狼蛛科年龄结构、性别比例等影响较大。
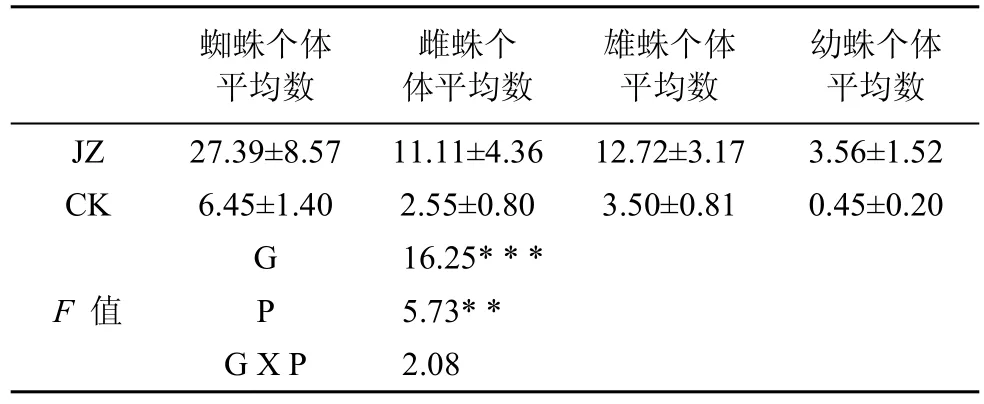
表2 两种茶园狼蛛科蜘蛛性别和年龄组成特征Tab.2 Quantitative characteristics of wolf spider guild in two tea plantations
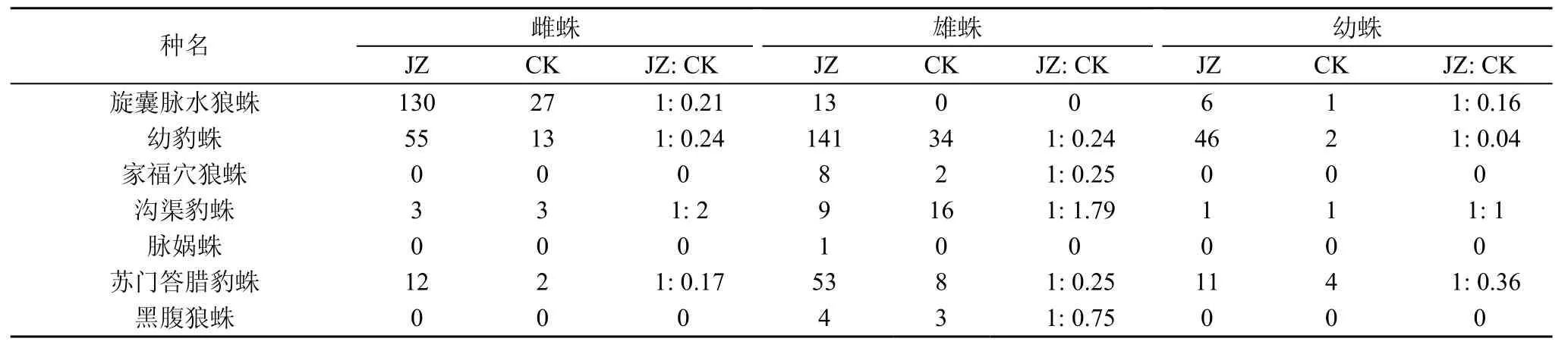
表3 两种茶园狼蛛科蜘蛛年龄性比Tab.3 Sex ratio of wolf spider guild in two tea plantations
3.3 茶园狼蛛科不同性别数量动态
狼蛛科蜘蛛群落动态个体数高峰时期从2010年夏季之后呈不断下降的趋势, 同时对两类茶园群落特征数量发生动态分别进行单因素方差分析。蜘蛛个体总数发生动态(图2), 从2010—2011年, 复杂栖境茶园蜘蛛总个体数量的活动密度, 大部分时期高于简单栖境茶园(F=5.83, P <0.05)。雄蛛个体数发生动态(图3), 复杂栖境茶园雄蛛个体数量的活动密度,除2010年的9月30日—12月29日和2011年的4月26日、5月11日外, 绝大部分时期均显著高于简单栖境茶园(F=7.39, 0.001< P <0.01)。此外, 雌蛛和幼蛛个体数发生动态(图4和图5), 两类茶园个体数量的活动密度差异不显著(F=3.77 ns 和 F=4.11 ns)。
3.4 不同种植管理茶园对狼蛛科生物学特征总体影响
对狼蛛科蜘蛛群落数量结构分布的各变量进行多元统计分析结果显示(图6): 由于第一主成分和第二主成分的累计贡献率达57.66%, 第一主成分的贡献率为37.93%, 蜘蛛数量分布受狼蛛科的家福穴狼蛛、幼豹蛛和旋囊脉水狼蛛等影响最大。第二主成分贡献率为19.73%, 其主要与狼蛛科的黑腹狼蛛、苏门答腊豹蛛和沟渠豹蛛紧密联系。复杂栖境茶园与简单栖境茶园样点随第一主成分属性进行空间分布, 二者清晰的被划分在第二主成分坐标轴的两边,复杂栖境茶园位置明显向家福穴狼蛛、幼豹蛛和旋囊脉水狼蛛等地面狼蛛科蜘蛛群落数量分布较多的方向偏移, 而简单栖境茶园则数量减少方向偏移,这一现象是由茶园栖境异质性所造成的。
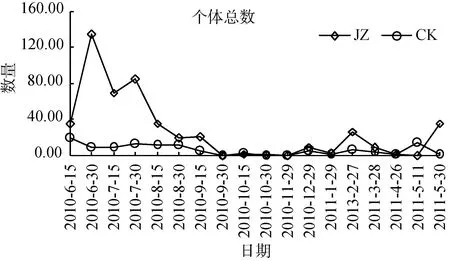
图2 两种茶园蜘蛛总个体数量发生动态变化Fig. 2 Dynamic changes of spider guild populations in two tea plantations
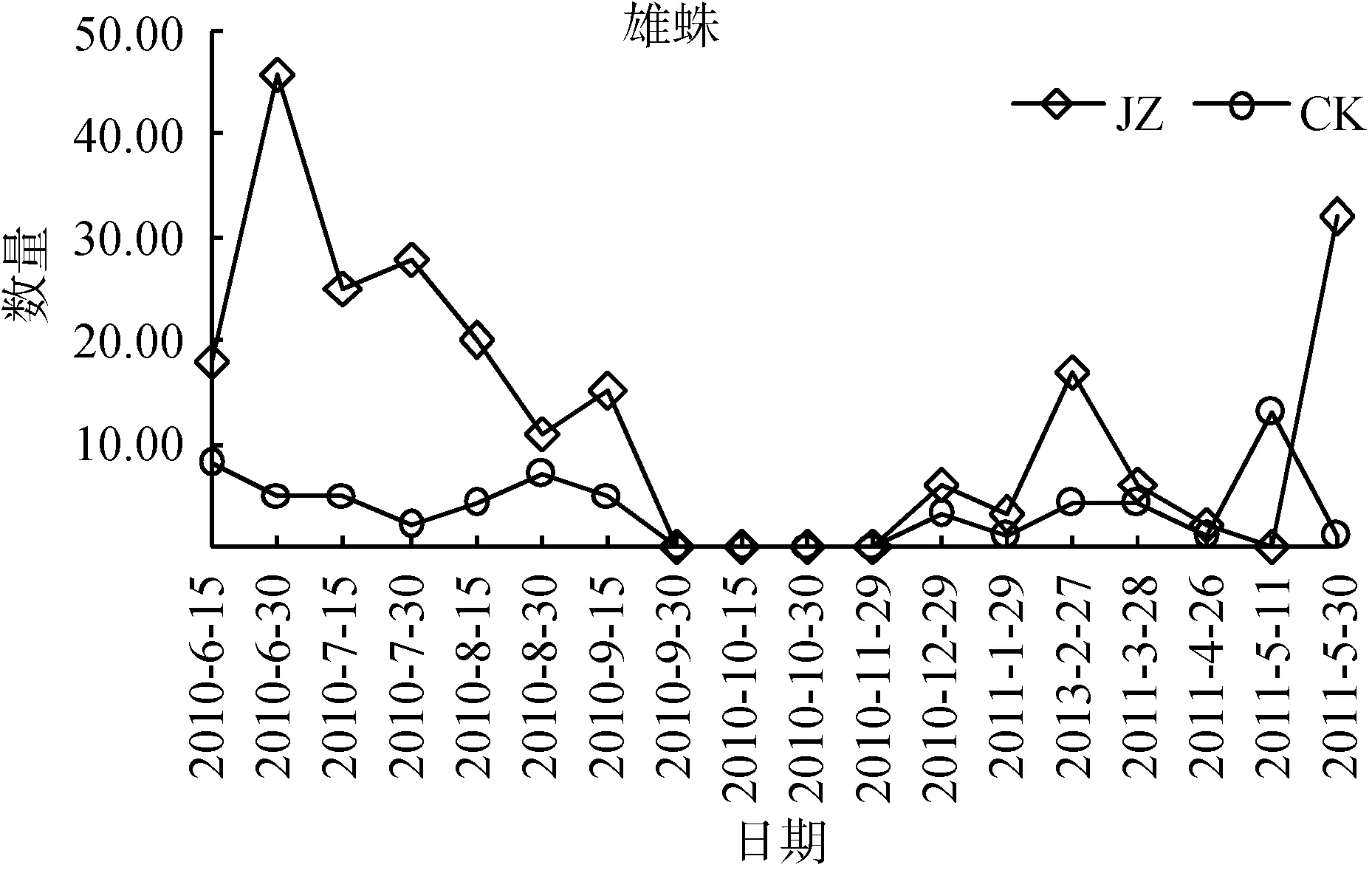
图3 两种茶园雄蛛数量发生动态变化Fig. 3 Dynamic changes of male spider populations in two tea plantations
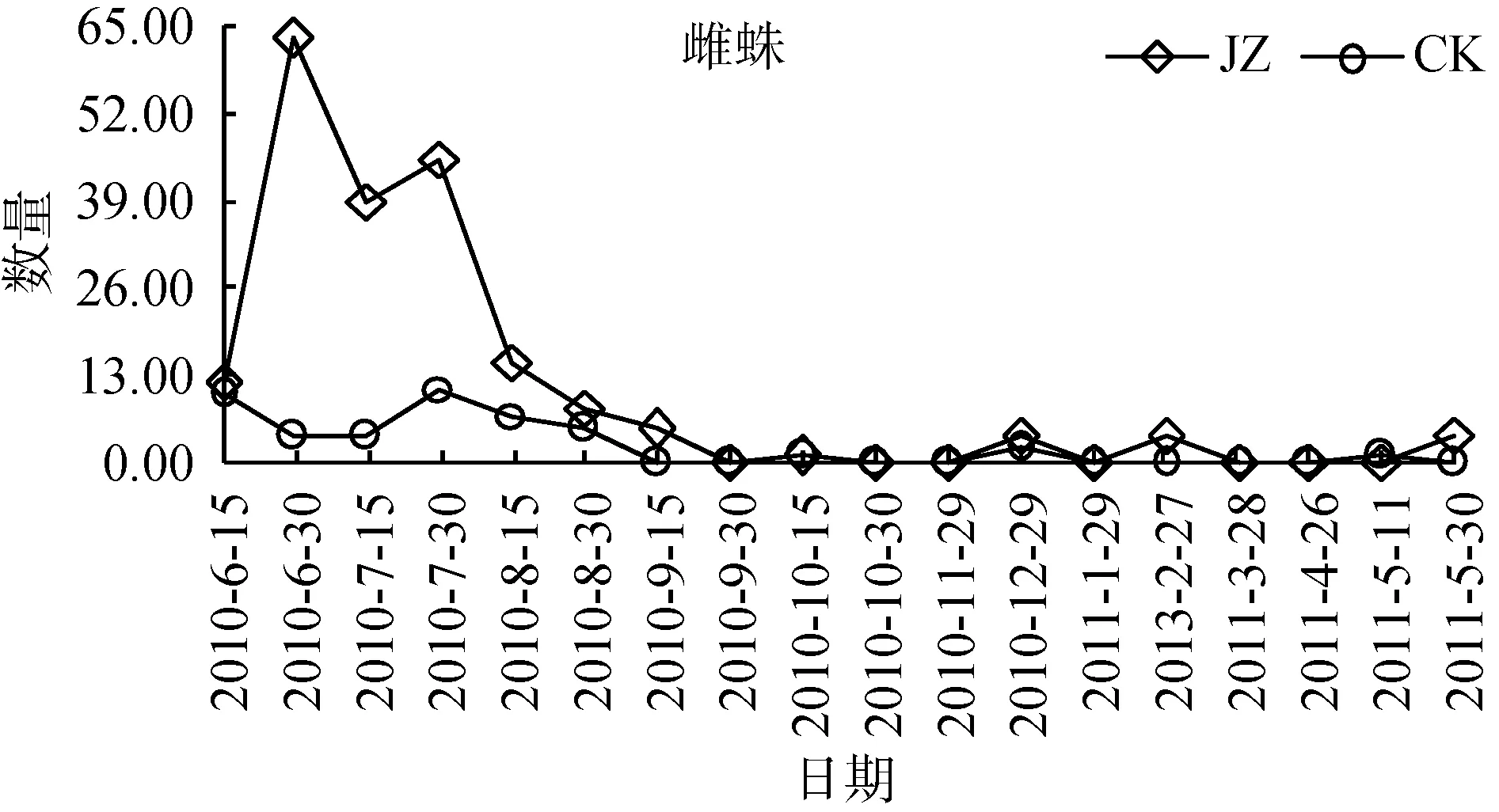
图4 两种茶园雌蛛数量发生动态变化Fig. 4 Dynamic changes of female spider populations in two tea plantations
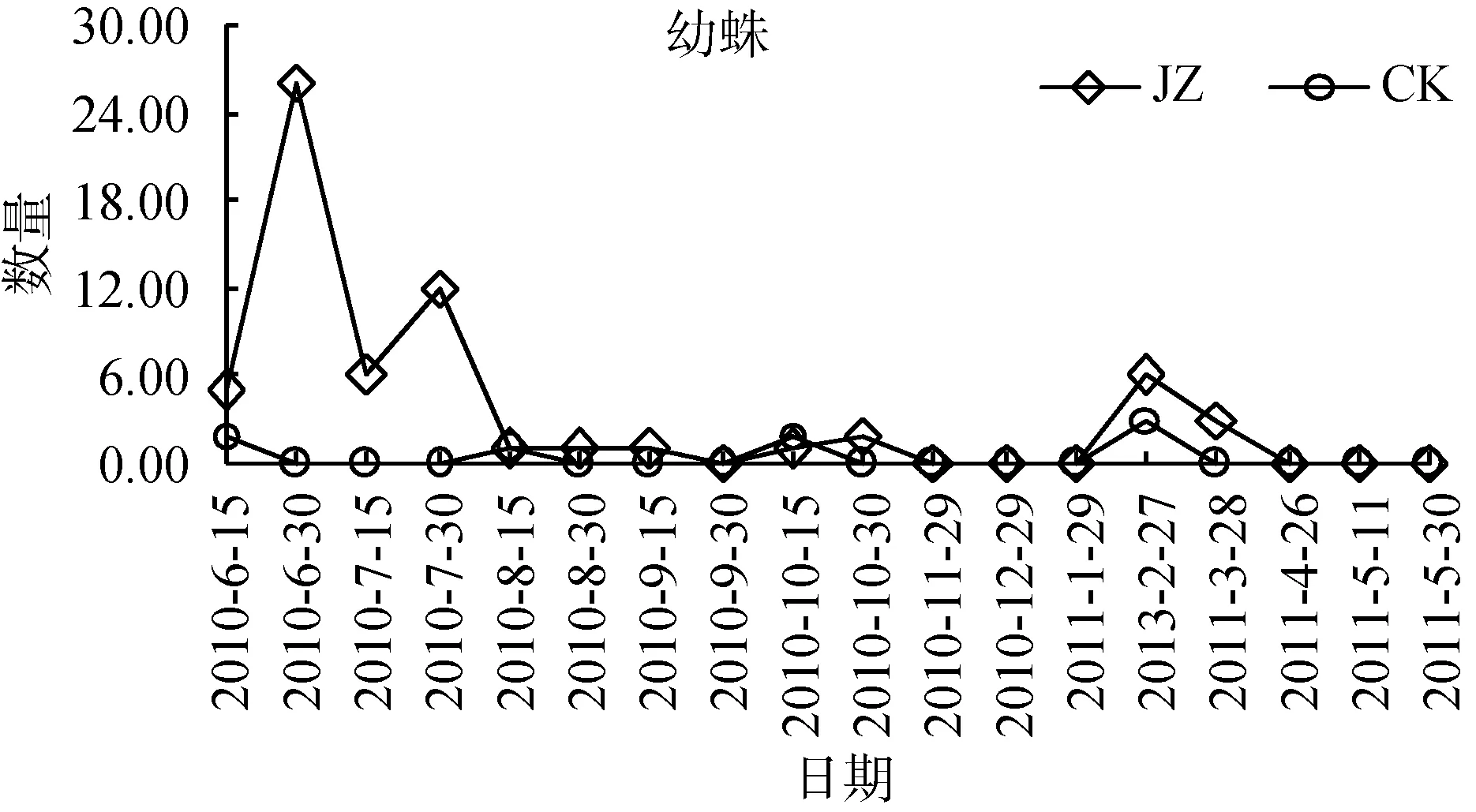
图5 两种茶园幼蛛数量发生动态变化Fig. 5 Dynamic changes of immature spider populations in two tea plantations
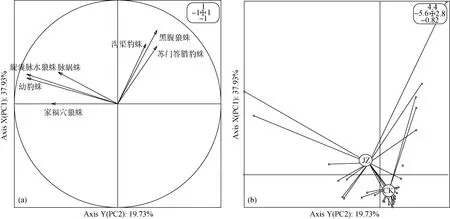
图6 两种茶园蜘蛛群落主成分分析Fig. 6 Principal component analysis of the spider guild of two tea plantations
4 讨论与结论
农田生境多样性种植对天敌栖息影响较大, 生境物种多样化与环境相对稳定, 可以提供天敌多样性与稳定性[18-19]。以往研究很少从栖境异质性去探讨, 只是侧重在蜘蛛对不同气候环境和化学农药适应性方面开展, 例如王定锋等[20]通过开展茶园蜘蛛群落结构的研究, 结果表明蜘蛛群落多样性受气象因子的影响, 湿度过高会对地面游猎型的蜘蛛产生影响。韩宝瑜[21]对茶园蜘蛛群落组成及动态差异进行调查分析, 研究发现有机茶园蜘蛛种类和个体数最多。而本次研究立足于目前农业关注化学农药减施的焦点, 从发展生态农业角度出发, 针对容易受到气候变化和农作物安全保障等问题所采取的措施; 利用复合间作模式改善栖境有效性, 特别是不施化学农药下实现害虫自然调控等均体现了重要的生态农业方式。由于蜘蛛对种植管理、人为干扰以及生态环境变化敏感, 能够很好地反映环境变化过程及其对生物多样性的影响[22]。所以, 探讨从狼蛛科蜘蛛群落变化过程来反映生态技术对改善茶园环境的友好变化过程, 亦是衡量实施生态农业技术措施对生物多样性影响的指示标志。本次试验研究共采集鉴定的狼蛛科有7个种, 复杂栖境茶园与简单栖境茶园对狼蛛科蜘蛛群落数量特征组成存在明显差异, 影响达到极显著(P<0.001 ); 复杂栖境茶园优势种合计3种, 其雌蛛、雄蛛和幼蛛个体数是明显增加。究其原因, 复杂栖境茶园, 实行多样性种植,从多方面实现环境友好, 而且茶园周围边界结构植物丰富, 为天敌提供栖息和食物来源, 有效促进狼蛛科蜘蛛群落结构特征数量的增加, 并使其年龄结构和性别比例得到合理调整。
茶园生态系统服务应用生物多样性, 是茶园害虫生态控制的发展方向[23]。以往研究表明农业生态系统中, 作物多样性种植有利于生物多样性保护[24-25]和提高蜘蛛群落的多样性[26]。固此, 复杂栖境茶园, 减少农药施用量, 是农业保护和利用生物多样性的重要体现。本研究系统分析了地面狼蛛科蜘蛛群落对复杂栖境茶园的反应; 研究表明, 茶园应用栖境异质性管理措施后, 狼蛛个体数量的活动密度显著高于简单栖境茶园。此外, 茶园栖境异质性对狼蛛科生物学特征影响较大, 主成分分析表明贡献率达57.66%。因此, 建议茶园以生态农业理念为导向, 广泛建立人与生态的和谐系统; 从农业耕作措施上尽量减免化学农药使用, 保护蜘蛛等物种的栖息地; 构建健康生态系统服务, 实现害虫可持续控制。
致谢:感谢湖北大学对蜘蛛标本鉴定。
[1] WHEATER C P, CULLEN W R, BELL J R. Spider communities as tools in monitoring reclaimed limestone quarry landforms[J]. Landscape Ecology, 2000, 15(5):401–406.
[2] 陈军, 宋大祥. 狼蛛科蜘蛛的繁殖行为[J]. 蛛形学报,1999, 8(1): 55–62.
[3] 颜忠诚, 陈永林. 放牧对蝗虫栖境结构的改变及其对蝗虫栖境选择的影响[J]. 生态学报, 1998, 18(3):278–282.
[4] 王思铭, 陈又清, 卢志兴, 等. 紫胶园异质性栖境下的蚂蚁共存机制[J]. 应用生态学报, 2010, 21(10): 2684–2690.
[5] 骆世明. 生态农业的模式与技术[M]. 北京: 化学工业出版社, 2009.
[6] 赵飞, 章家恩. 广东传统农业的生态学智慧述论[J]. 生态科学, 2015, 34(2): 173–178.
[7] COLE L J, MCCRACKEN D I, DOWNIE I S, et al.Comparing the effects of farming practices on ground beetle (Coleoptera: Carabidae) and spider (Araneae)assemblages of Scottish farmland[J]. Biodiversity and Conservation, 2005, 14: 441–460.
[8] 罗媛媛, 刘金亮, 黄杰灵, 等. 千岛湖陆桥岛屿上黑腹狼蛛(Lycosa coelestris)遗传多样性及其受生境片段化的影响[J].科学通报, 2014, 59(19): 1851–1860.
[9] 张征田, 庞振凌, 夏敏, 等. 镉污染对不同生境拟水狼蛛氧化酶和金属硫蛋白应激的影响[J]. 生态学报, 2011,31(16): 4579–4585.
[10] 张征田, 夏敏, 彭宇, 等. 河南南阳不同生境土壤重金属含量及其对拟水狼蛛生物学特性的影响[J]. 昆虫学报,2009, 52(9): 994–999.
[11] 王智, 宋大祥, 朱明生. 稻田蜘蛛和害虫的生态位研究[J].华南农业大学学报, 2005, 26(2): 47–51.
[12] 张永强, 侯美珍, 陆温, 等. 玉米蜘蛛群落结构及动态研究[J].广西农业大学学报, 1997, 16(4): 268–274.
[13] 汪波, 柯紫君, 汪昕蕾, 等.狼蛛对黑腹果蝇捕食数量的PCR检测[J].浙江农业学报, 2015, 27 (7): 1198–1201.
[14] 马艳滟, 李巧, 冯萍, 等.云南苍山火烧迹地不同恢复期地表蜘蛛群落多样性[J].生态学报, 2013, 33(3): 964–974.
[15] 林胜, 陈李林, 尤民生, 等.多作对稻田蜘蛛功能团结构及多样性的影响[J].福建农林大学学报(自然科学版),2011, 40(1): 1–7.
[16] BERGER WH, PARKER FL. Diversity of planktonic foraminifera in deep-sea sediments[J]. Science, 1970, 168:1345–1347.
[17] THIOULOUSE J, CHESSEL D, DOLEDEC S, et al.ADE-4: a multivariate analysis and graphical display software[J]. Statistics and Computing, 1997, 7: 75–83.
[18] FAHRIG L, BAUDRY J, BROTONS L, et al. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes[J]. Ecology Letters, 2011, 14: 101–112.
[19] 杨芳, 贺达汉. 生境破碎化对生物多样性的影响[J]. 生态科学, 2006, 25(6): 564–567.
[20] 王定锋, 刘丰静, 李慧玲, 等.宁德市低海拔茶园蜘蛛群落结构及发生动态[J]. 茶叶科学, 2013, 33(5): 457–464.
[21] 韩宝瑜. 有机、无公害和普通茶园蜘蛛群落组成及动态差异[J]. 蛛形学报, 2005, 14(2): 104–107.
[22] BUCHHOLZ S. Ground spider assemblage as indicator for habitat structure in inland sand ecosystems[J]. Biodiversity and Conservation, 2010, 19: 2565–2595.
[23] 侯柏华, 谭济才, 张润杰. 茶园害虫生态控制若干问题的探讨[J]. 生态科学, 2004, 23(3): 261–264.
[24] 黎健龙, 唐劲驰, 赵超艺, 等.不同景观斑块结构对茶园节肢动物多样性的影响[J].应用生态学报, 2013, 24(5):1305–1312.
[25] 高东, 何霞红, 朱书生.利用农业生物多样性持续控制有害生物[J]. 生态学报, 2011, 31(24): 7617–7624.
[26] 黎健龙, 涂攀峰, 陈娜, 等. 茶树与大豆间作效应分析[J].中国农业科学, 2008, 41(7): 2040–2047.
Effects of varying habitat heterogeneity on the wolf spider (Araneae:Lycosidae) communities in tea plantations
LI Jianlong, TANG Hao, ZHOU Bo, FANG Huachun*
Tea Research Institute, Guangdong Academy of Agricultural Sciences; Guangdong Provincial Key Laboratory of Tea Plant Resources Innovation and Utilization, Guangzhou 510640, China
Spiders can play an important ecological role in agricultural ecosystems, and they can also serve as indicators of improved ecosystem services. We examined the community-level responses of the wolf spider (Araneae: Lycosidae) in terms of its species composition, structure, and diversity by collecting samples from subtropical hilly tea plantations subjected to two types of production management: complex habitat management (JZ) and routine management (CK). The results showed that there were three dominant species n plantation JZ, and four dominant species in CK. The quantitative characteristics of community composition were significantly richer in plantation JZ than in CK (P<0.001). Specifically, the numbers of females, males, and immature wolf spiders in plantation JZ were 4.36-,3.63-, and 7.91-fold higher than those in CK. In plantation JZ, the characteristics of spider age, gender, and number of functional clusters were much higher than those in CK (P< 0.05), including the male and immature individuals total dynamic. Principal component analysis howed that distribution of community orientation offsets to the number of wolf spiders of the three species Houa jiafui, Pardosa pusiola,and Venonia spirocysta was higher in plantation JZ than in CK. Taken together, the results demonstrate that the complex habitat management of tea plantations is conducive to preserving the diversity of wolf spider species. In addition, to enhance natural pest control, such a management approaches may contribute further towards the sustainable improvement of agricultural ecosystems.
tea plantations; habitat heterogeneity; Lycosidae; community structure
10.14108/j.cnki.1008-8873.2017.03.023
S435.711; Q959.226+.2
A
1008-8873(2017)03-160-06
黎健龙, 唐颢, 周波, 等. 茶园异质性栖境对狼蛛科蜘蛛群落结构的影响[J]. 生态科学, 2017, 36(3): 160-165.
LI Jianlong, TANG Hao, ZHOU Bo, et al. Effects of varying habitat heterogeneity on the wolf spider (Araneae: Lycosidae) communities in tea plantations [J]. Ecological Science, 2017, 36(3): 160-165.
2016-08-18;
2016-10-12
国家自然科学基金项目(31600559); 广东省自然科学基金项目(2015A030313563); 广东省科技计划项目(2013B060500062, 2014A070713014,2014A020208054, 2015A070710012); 广东省农业科学院院长基金(201630)
黎健龙(1982—), 男, 广东广州人, 助理研究员, 主要从事茶树生态栽培与植保研究, E-mail: skylong.41@163.com
*通信作者:方华春, 男, 高级农艺师, E-mail: fanghc@126.com
