青藏高原植物物候的变化及其影响*
2017-07-06孟凡栋斯确多吉崔树娟王奇李博文汪诗平
孟凡栋,斯确多吉,崔树娟,王奇,李博文,汪诗平③†
①中国科学院青藏高原研究所,北京 100101;②中国科学院青藏高原地球科学卓越创新中心,北京 100101;③中国科学院大学,北京 100049
青藏高原植物物候的变化及其影响*
孟凡栋①③,斯确多吉①②,崔树娟①③,王奇①③,李博文①③,汪诗平①②③†
①中国科学院青藏高原研究所,北京 100101;②中国科学院青藏高原地球科学卓越创新中心,北京 100101;③中国科学院大学,北京 100049
植物物候是植物生活史的周期性循环事件。在气候变化和人类活动的影响下,青藏高原植物物候发生了显著的变化。近年来,青藏高原的物候初始期变化总体表现为返青期和初花期提前、枯黄期推迟的趋势,而果实期则保持相对稳定;增温延长了生长季,主要是通过延长开花等繁殖物候而实现的。青藏高原的物候变化主要受温度、水分和放牧的驱动,暖湿化和适度放牧有利于植物提前或延长物候期,而暖干化则延迟或缩短物候期。物候变化对植物种群、群落、生态系统、农牧区生产以及旅游业(如赏花节)等都会产生显著的影响。然而,目前青藏高原植物物候研究仍然十分缺乏,尤其是物候变化对生态系统结构和功能的影响研究更少。针对当前青藏高原物候研究存在的问题给出了一些建议:在未来的研究中应该尽可能从生理尺度到生态系统尺度的不同视角来研究气候变化和人类活动对物候的影响,以及物候变化对生态系统结构和功能的反馈。
青藏高原;气候变化;放牧;植物物候;影响
植物物候是植物生活史的周期性循环事件,如返青或者开花[1],在中国是一门古老的学科,已有数千年的历史[2]。因为物候与气候变化存在高度的相关性,使其成为研究气候变化最简单有效的证据。青藏高原被称为“世界屋脊”,因其独特的自然环境而在全球变化中扮演重要的角色。青藏高原生态系统对全球变化极为敏感,在气候变化和人类活动的影响下,青藏高原植物物候发生了显著的变化,而物候的变化将会对草地生态系统的生产力以及碳循环等产生重要的影响。因此,深入理解青藏高原植物物候的变化机制,采取合适的方法和对策应对青藏高原生态系统的变化,对青藏高原的草地规划和管理具有重要的意义。
1 青藏高原植物物候变化
青藏高原的草本植物生命周期通常只有一年,随着气候的季节性变化,植物包括返青、现蕾、开花、结实、果后营养、初黄和枯黄七个物候序列初始期和相应的持续期。七个物候序列间是存在内在联系以及等级关系的。返青、果后营养、初黄和枯黄组成了被子植物的营养期;现蕾、开花和结实组成了被子植物的繁殖期;而营养期和繁殖期又组成了植物的整个生长季(图1)。不同等级的物候序列间对气候变化和人类活动的响应存在权衡。也就是说,一个物候期的改变可能会对后序物候期的变化产生影响。所以,对植物物候序列的研究有助于更深入地理解气候变化和人类活动的影响以及揭示单个对其他物候的影响大小。

图1 不同等级的物候事件[3]
1.1 营养期的物候变化
对营养物候初始期而言,近30年来,青藏高原的物候变化总体表现为返青期提前、枯黄期推迟的趋势[4-6]。果后营养期和初黄期的研究相对较少,有限的研究发现它们是延迟的[7]。相对于初始期,物候的持续期研究更少,其中,在海北站有相对完整的物候记载,研究发现各营养物候初始期相应的物候持续期也是显著延长的。同时,植物的营养期显著延长,主要是由延长的枯黄期引起的[3]。
返青期是当前物候研究关注最多的物候事件,但研究方法和区域的不同使得物候的变化还存在较多争议。当前青藏高原的物候研究以遥感为主,然而它们的结果却存在较大的差异。尽管这期间青藏高原年均温度呈上升趋势,但自20世纪90年代中后期到21世纪初,物候变化呈现与上述变化不大甚至是相反的趋势[6,8-10](图2),而自2005年以后,返青期又开始提前[4]。这说明遥感方法还存在很多不确定性。地面的定位观测结果显示,近30年不同站点的物候返青期既没有一致的提前趋势,也没有出现遥感方法中的拐点变化[11](图3)。这说明不同尺度的物候观测之间不能相互验证。
青藏高原物候变化存在显著的区域差异。总体上,从高原东南部到西北返青期是逐渐延迟的,枯黄期是逐渐提前的,而生长季长度则相应地缩短[12]。尤其是青藏高原东北部(青海省东部农业区、环青海湖区、三江源区)的植物物候提前幅度最大[11,13];而中东部以及南部区域的植物物候变化幅度较小,甚至有些是倾向于延迟的[11,13]。同时,海拔变化也对区域物候差异有显著影响,海拔3 500 m以下的物候变化没有规律性,而超过3 500 m后物候会随海拔变化呈现规律性的变化[12]。有人却认为只有高寒草甸的返青期是显著提前的,而高寒草原并没有发生显著的变化[14]。

图2 1982—2011年的植被返青期的年际变异[6]
1.2 繁殖期的物候变化
因遥感无法捕捉到个体植物的物候信息,而青藏高原又缺乏物候的地面观测,所以青藏高原繁殖期物候的研究相对较少,并且这些观测站点也主要集中在高原东部区域(图4)。少数研究的结果表明,近年来初花期是显著提前的,而初果期或末果期保持相对稳定[7,15]。高寒草甸群落中,开花越早的物种,花期持续期越长[3];并且,开花持续期延长会显著延长植物的繁殖期[3]。
1.3 生长季的变化
无论遥感还是地面观测,大多数的研究表明青藏高原的生长季是延长的[4-6]。有人认为生长季的延长主要受返青期的显著提前影响[4]。这是因为他们的研究只关注了返青期和枯黄期的变化,而根据这两者推算出它们的贡献量大小。但实际上,不同物候期之间是存在权衡的,海北站所观测的物种水平生长季长度变化表明所有物种的生长季延长,并且生长季延长主要是由繁殖期延长引起的[3]。
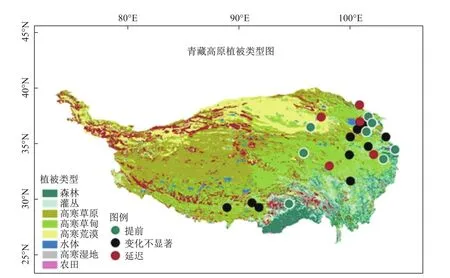
图3 青藏高原草本植物返青期地面观测站点(绿色、黑色和红色分别表示返青期提前、变化不显著、延迟)[11]
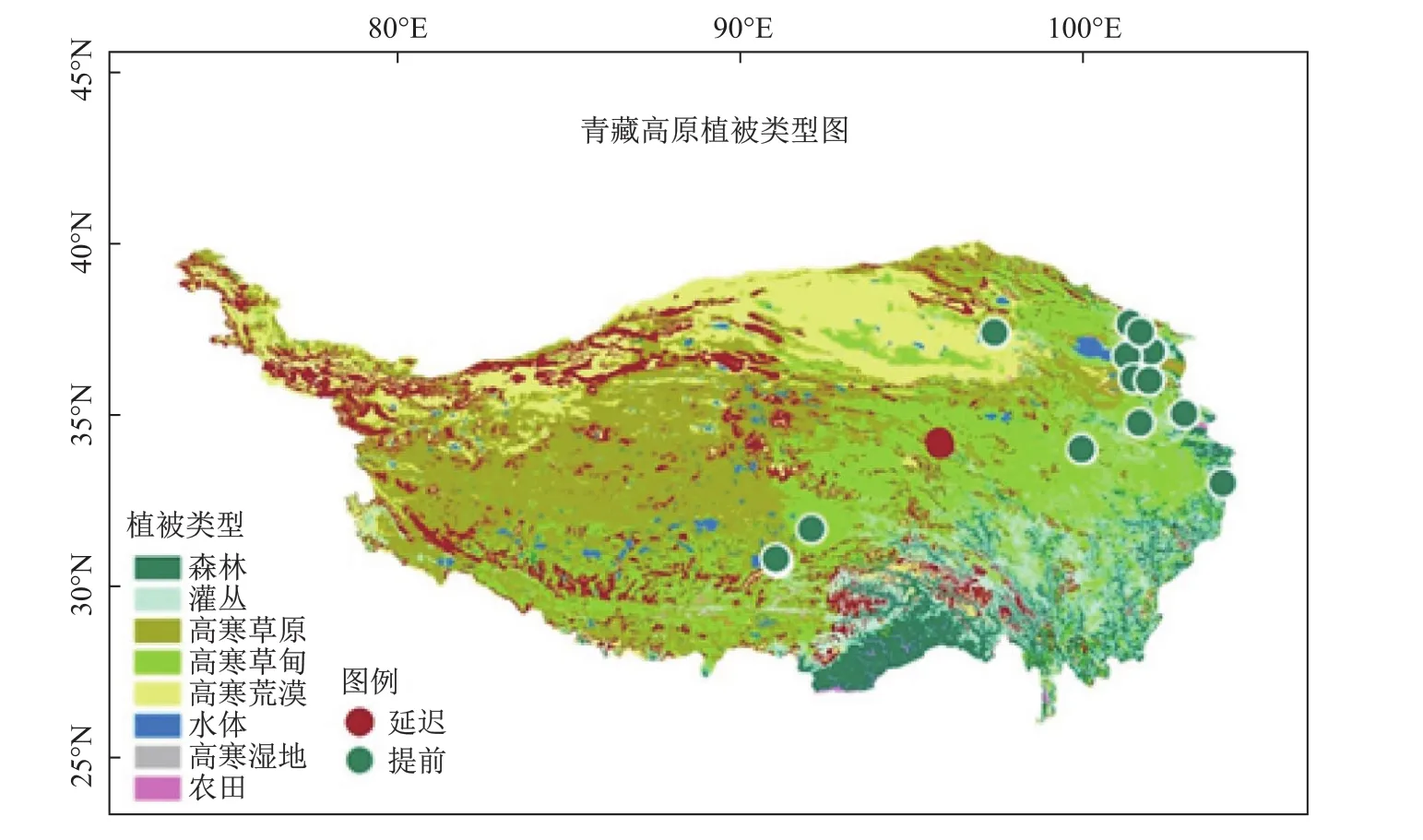
图4 青藏高原草本植物初花期地面观测站点(绿色和红色分别表示初花期提前和延迟)
2 青藏高原植物物候变化的原因
2.1 自然因素
植物物候受多种环境因素的影响,但当前物候研究普遍认为温度和水分变化决定物候期的改变[3,7,16-20]。青藏高原近几十年经历了持续的增温过程,而水分的变化却是多样的。
对早期物候(返青期、现蕾期和初花期)而言,增温的效应是存在季节差异的。春季增温会使得早期物候提前[4,19,21]。例如:春季每增温1℃,返青期整体上提前4.1天[21];冬季增温对早期物候的影响存在争议性。有人认为冬季增温影响了青藏高原植物的春化过程从而延迟返青;也有人认为冬季增温对早期物候影响不大[22],因为冬季增温幅度还未达到影响植物对低温需求的阈值[23]。有关水分变化对早期物候影响的研究较少。在高寒区域植物解除休眠前的积雪融化时间对其早期物候影响较大[24];但对植物活动前期无积雪覆盖的青藏高原而言,土壤解冻时间及其后续土壤水分含量是影响植物早期物候的主要因素[25]。同时,冬春季降水量的增加也能显著提前早期物候[26],在干旱区域这种效应更明显[22]。尤其是,有研究发现有些物种能够感知季风雨的来临而返青[27]。总而言之,以上这些结果说明早期物候变化主要受温度和水分有效性变化的影响,暖湿化能够提前早期物候,而暖干化则延迟早期物候。
对中期物候(果期)而言,结实的初始期或结束期对温度变化的响应是保持相对稳定的,这有利于植物在多变的环境中繁衍生息[7]。但也有人认为降水的时间变化对繁殖期的影响较大。例如,吕新苗等[28]发现生长季初期降雨的提前显著提前了果期。
对晚期物候(果后营养期、初黄期和枯黄期)而言,增温引起青藏高原的高寒草地植被果后营养期、初黄期和枯黄期显著延迟[4,15,21]。水分含量增加也会显著地延迟晚期物候[18]。
相对于初始期,较少有研究关注物候持续期变化对气候变化的响应[3]。有研究表明增温缩短了植物的生长季,这可能是因为增温加速了植物的生长导致的[29]。然而,多数研究表明增温延长了营养期、繁殖期和生长季[3,18-19]。水分状况对物候持续期也有显著影响:高水分含量会显著延长持续期[18];而干旱一般会缩短植物的物候持续期,尤其是开花期,因为可以减少因开花而增加的水分丧失量[18]。但低水分状况可以减缓花芽和结实的过程[30],然而也有相反的研究结果报道[19]。
综上所述,青藏高原暖湿化有利于植物的发育,从而提前或者延长植物物候期;而暖干化则抑制植物的发育,从而延迟或者缩短植物物候期。
2.2 人为因素
青藏高原是放牧顶级群落[31],放牧作为青藏高原重要的人类活动,对微环境和植物生活史对策有着显著的干扰[32-33]。然而,关于放牧对物候的影响报道较少。放牧能够显著地提高地表温度,这可能是因为地表立枯物和凋落物减少,提高了透光率,而土壤表层可以吸收更多的红外辐射[33]。同时,放牧也会导致土壤湿度下降,但差异并不显著[34]。在这种情境下,有研究发现放牧引起物种返青始期提前和枯黄末期延迟,并且延长了生长季[33-34]。有人对比了围封与放牧处理后物候的变化发现,围封显著地改变了物候期的变化。以生长季长度变化为例,围封3年以后显著延长了高寒草甸建群种高山嵩草(Kobresia pygmaea)的生长季长度[35](图5)。尤其增温和放牧之间存在交互作用,这种效应对物候的影响是叠加的[33-34]。在这两种效应影响下,植被较易发生退化[36],而退化又会反过来影响物候的变化[36-37]。例如:上述遥感监测返青期出现差异的原因可能是因为植被退化导致群落盖度下降,从而提高地表反射率,进而使气温降低。这种作用在春季尤为明显[37]。

图5 围封与放牧间的高山嵩草生长季长度差异[35](*表示围封与放牧间在0.05水平下的显著性差异)
3 青藏高原植物物候变化的影响
理论上讲,植物物候变化直接导致的后果是改变了植物各生育期开始和结束的时间,即改变了各生育期长短,从而直接或间接影响植物种群、群落以及生态系统的结构和功能[38-40]。由于青藏高原相关的物候变化影响/后果研究十分稀少,故此部分内容主要从植物种群、群落、生态系统以及植物生产等层面简要介绍物候变化的常规以及可能后果,同时穿插、引用青藏高原相关的有限研究内容。
3.1 植物种群
物候变化对植物个体和种群的影响涉及植物生理(如抗逆性)、生长(如生物量的增加幅度)与繁殖(如开花或结果的数量、比率等)的变化。例如:Inouye[41]发现,返青提前使植物易于遭受早春冻害和霜害影响,致使植物生长与繁殖受阻。开花时间变化时,若传粉昆虫出现的时间不相应改变,则会导致授粉率和繁殖率降低[42]。另外,Xia 等[43]研究表明,物候变化是控制植物生产力的重要因素之一,因此当枯黄期起始时间不变或延迟前提下,返青提前必然导致生长期延长,植物生产量(单位面积每年度的生产量)提高。生长期延长还可能导致植物对营养和繁殖生长的分配时间发生变化[44],致使植物繁殖率及其影响的种群结构(年龄结构)、密度发生变化[45-46],但相关研究鲜有报道。
3.2 植物群落
群落尺度上,当某一物种因生长期延长、生物量增加、种群密度提高时,与此伴生的其他物种则可能因遭受竞争排斥而降低种群密度,甚至消失[47]。有研究表明,增温导致灌木类植物物候提前[48],致使其种群扩散,降低了其他适宜家畜采食的优良牧草在群落中的比例[49],从而降低了草地提供牧业生产的服务功能。研究表明,植物初级生产力的大小与物候变化十分相关[44,50-51],但这些研究仅停留在基于观测的假设和推理基础之上,直接连接物候变化和群落结构之间关系的研究还相对匮乏。另外,有关物候变化引发的群落结构(物种组成、丰富度等)变化研究也十分稀少,但就理论推理看,物候变化引起种群结构变化继而导致物种组成和丰富度发生变化。
3.3 生态系统
物候变化还可能引起生态系统结构和功能发生变化。例如:气候变化引起的物候变迁(如开花和结果起始时间以及持续时间变化),可能导致依赖于植物的某一种资源(花蜜或者果实等)的专一性采蜜昆虫以及专一性采食果实或种子的鸟类生存遭受干扰[52],继而导致生态系统结构和功能发生变化。另外,物候变化改变植物和群落生长期长短以及生产力,间接导致生态系统呼吸以及碳储汇等功能发生变化[53]。
3.4 农牧业生产
农牧业生产是青藏高原的支柱产业。西藏当地居民历来注重利用植物物候变化特征从事农牧业生产[54]。近年来,随着全球气候不断变暖,我国西北地区各地,包括青藏高原地区的农作物种植时间和返青物候有所提前[55-57],农作物产量也因此获得了提高[56],这显然与物候变化改变植物初级生产力之间的普遍关系一致[44,51]。另外,如前所述,气温变暖还可能导致灌木类植物物候提前[48],使其产量提高,从而降低优质牧草的产量[49],影响草地对牧业生产的服务功能。当前在青藏高原各类草地生态系统中,除了气候变暖,还包括降水量、二氧化碳浓度、氮沉降水平以及人类活动(放牧强度和模式)等多种环境因素的变化。这些环境因素的独立及其互作效应对草地植物/植被物候的各类事件及其持续时间的影响以及物候变化对草地植物种群、群落以及生态系统的影响是复杂的,但相关研究十分稀少。
3.5 其他
物候作为一些特殊行业(如观花旅游行业)的重要指示,其变化特征会直接或间接影响相关行业的发展。例如:近年来,随着国力增强,人们消费需求提高,全国各地相继开发了一系列观花、赏花旅游节日(有些是传统的赏花节日),而这些产业在某种程度上受开花起始时间与持续时间长短影响。近年来,青藏高原各区域也开发了各类观花旅游项目,其中较为著名的有青海的油菜花旅游季、西藏林芝的桃花节等,但相关研究十分稀少。
4 有关思考与建议
尽管气候变化与物候相关的研究十分热门,而且在国内外已积累了一定数量的研究基础,但大多研究仅限于现象描述,很少涉及生理、生态过程和机理研究。另外,现有研究大多仅限于利用单一某种研究手段(如遥感观测或野外试验),很少考虑到结合各类研究手段和方法综合探讨气候变化对物候的影响。再者,尽管大多研究假设或预测物候变化对植物种群、群落以及生态系统功能的影响,直接连接和证明各类物候事件及其持续时间的变化与生态系统不同尺度之间的关系研究却十分匮乏,因此我们建议:
(1)加强从生理、生态的不同视角探讨气候变化(如变暖)对植物物候各事件及其持续时间的影响过程和机理;
(2)加强遥感观测、模型研究和控制试验(实验)之间互通、合作,从不同尺度、不同视角探讨气候变化与物候变迁之间的关系;
(3)应加强利用试验(实验)手段剖析和验证物候变化及其功能之间的关系。
(2017年5月8日收稿)
[1] WOLKOVICH E M, COOK B I, DAVIES T J. Progress towards an interdisciplinary science of plant phenology: building predictions across space, time and species diversity [J]. New Phytologist, 2014, 201(4): 1156-1162.
[2] 竺可桢.中国近五千年来气候变迁的初步研究[J]. 中国科学, 1973, 9(2): 168-189.
[3] LI X, JIANG L, MENG F, et al. Responses of sequential and hierarchical phenological events to warming and cooling in alpine meadows [J]. Nature Communications, 2016, 7:12489.
[4] SONG CQ, YOU S C, KE L H, et al. Spatio-temporal variation of vegetation phenology in the Northern Tibetan Plateau as detected by MODIS remote sensing [J]. Acta Phytoecologica Sinica, 2011, 35(8): 853-863.
[5] 张宪洲, 杨永平, 朴世龙, 等. 青藏高原生态变化[J]. 科学通报, 2015, 60(32): 3048-3056.
[6] ZHANG G, ZHANG Y, DONG J, et al. Green-up dates in the Tibetan Plateau have continuously advanced from 1982 to 2011 [J]. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(11): 4309-4314.
[7] JIANG L L, WANG S P, MENG F D, et al. Relatively stable response of fruiting stage to warming and cooling relative to other phenological events [J]. Ecology, 2016, 97(8): 1961-1969.
[8] PIAO S L, FANG J Y, HE J S. Variations in vegetation net primary production in the Qinghai-Xizang Plateau, China, from 1982 to 1999 [J]. Climatic Change, 2006, 74(1/2/3): 253-267.
[9] YU H, LUEDELING E, XU J. Winter and spring warming result in delayed spring phenology on the Tibetan Plateau [J]. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(51): 22151-22156.
[10] SHEN M, SUN Z, WANG S, et al. No evidence of continuously advanced green-up dates in the Tibetan Plateau over the last decade [J]. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(26): E2329.
[11] CHEN X, AN S, INOUYE D W, et al. Temperature and snowfall trigger alpine vegetation green-up on the world's roof [J]. Global Change Biology, 2015, 21(10): 3635-3646.
[12] DING M, ZHANG Y, SUN X, et al. Spatiotemporal variation in alpine grassland phenology in the Qinghai-Tibetan Plateau from 1999 to 2009 [J]. Chinese Science Bulletin, 2013, 58(3): 396-405.
[13] 丁明军, 张镱锂, 孙晓敏, 等. 近10年青藏高原高寒草地物候时空变化特征分析[J]. 科学通报, 2012, 57(33): 3185-3194.
[14] ZHOU H K, YAO B Q, XU W X, et al. Field evidence for earlier leafout dates in alpine grassland on the eastern Tibetan Plateau from 1990 to 2006 [J]. Biology Letters, 2014, 10 (8): 1565-1579.
[15] 徐振锋, 胡庭兴, 李小艳, 等. 川西亚高山采伐迹地草坡群落对模拟增温的短期响应[J]. 生态学报, 2009, 29(6): 2899-2905.
[16] MENG F D, JIANG L L, ZHANG Z H, et al. Changes in flowering functional group affect responses of community phenological sequences to temperature change [J]. Ecology, 2017, 98(3): 734-740.
[17] MENG F, ZHOU Y, WANG S, et al. Temperature sensitivity thresholds to warming and cooling in phenophases of alpine plants [J]. Climatic Change, 2016, 139: 579-590.
[18] MENG F, CUI S, WANG S, et al. Changes in phenological sequences of alpine communities across a natural elevation gradient [J]. Agricultural and Forest Meteorology, 2016, 224: 11-16.
[19] WANG S P, WANG C S, DUAN J C, et al. Timing and duration of phenological sequences of alpine plants along an elevation gradient on the Tibetan plateau[J]. Agricultural and Forest Meteorology, 2014, 189/190: 220-228.
[20] WANG S P, MENG F D, DUAN J C, et al. Asymmetric sensitivity of first flowering date to warming and cooling in alpine plants [J]. Ecology, 2014, 95(12): 3387-3398.
[21] PIAO S, CUI M, CHEN A, et al. Altitude and temperature dependence of change in the spring vegetation green-up date from 1982 to 2006 in the Qinghai-Xizang Plateau [J]. Agricultural and Forest Meteorology, 2011, 151(12): 1599-1608.
[22] SHEN M. Spring phenology was not consistently related to winter warming on the Tibetan Plateau [J]. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(19): E91-E92.
[23] CONG N, SHEN M, PIAO S, et al. Little change in heat requirement for vegetation green-up on the Tibetan Plateau over the warming period of 1998–2012 [J]. Agricultural and Forest Meteorology, 2017, 232: 650-658.
[24] FORREST J, INOUYE D W, THOMSON J D. Flowering phenology in subalpine meadows: does climate variation influence community coflowering patterns? [J]. Ecology, 2010, 91(2): 431-440.
[25] DORJI T, TOTLAND O, MOE S R, et al. Plant functional traits mediate reproductive phenology and success in response to experimental warming and snow addition in Tibet [J]. Glob Chang Biol, 2013, 19(2): 459-472.
[26] 曾彪. 青藏高原植被对气候变化的响应研究(1982-2003)[D]. 兰州:兰州大学, 2008.
[27] LI R, LUO T, MOLG T, et al. Leaf unfolding of Tibetan alpine meadows captures the arrival of monsoon rainfall [J]. Sci Rep, 2016, 6: 20985.
[28] 吕新苗, 康世昌, 朱立平, 等. 西藏纳木错植物物候及其对气候的响应[J]. 山地学报, 2009, 27(6): 648-654.
[29] HALEVY A. Handbook of flowering, vol 1–6 [M]. Boca Raton: CRC Press, 1995.
[30] GALEN C. Why do flowers vary? [J]. BioScience, 1999, 49(8): 631.
[31] 李雄, 胡向阳, 杨永平. 青藏高原重要牧草高山嵩草的研究现状[J].草业科学, 2013, 206(1): 30-39.
[32] 赵彬彬, 牛克昌, 杜国祯. 放牧对青藏高原东缘高寒草甸群落27种植物地上生物量分配的影响[J]. 生态学报, 2009, 29(3): 1596-1606.
[33] 徐广平. 高寒草甸植物和植物群落对增温和放牧的响应与适应研究[D]. 兰州: 兰州大学, 2010.
[34] 段吉闯. 气候变化和放牧对高寒草甸主要植物和植物群落的影响[D]. 北京: 中国科学院研究生院, 2012.
[35] ZHU J, ZHANG Y, LIU Y. Effects of short-term grazing exclusion on plant phenology and reproductive succession in a Tibetan alpine meadow [J]. Scientific Reports, 2016, 6: 27781.
[36] BISIGATO A J, CAMPANELLA M V, PAZOS G E. Plant phenology as affected by land degradation in the arid Patagonian Monte, Argentina: A multivariate approach [J]. Journal of Arid Environments, 2013, 91: 79-87.
[37] CHEN H, ZHU Q, WU N, et al. Delayed spring phenology on theTibetan Plateau may also be attributable to other factors than winter and spring warming [J]. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(19): E93.
[38] BUIDE M L, D AZ-PEROMINGO J A , GUITI N J. Flowering phenology and female reproductive success in Silene acutifolia Link ex Rohrb [J]. Plant Ecology, 2002, 163(1): 93-103.
[39] GALEN C, STANTON M L. Consequences of emergence phenology for reproductive success in Ranunculus adoneus (Ranunculaceae) [J]. American Journal of Botany, 1991, 78(7): 978-988.
[40] MINER B G, SULTAN S E, MORGAN S G, et al. Ecological consequences of phenotypic plasticity [J]. Trends in Ecology & Evolution, 2005, 20(12): 685-692.
[41] INOUYE D W. Effects of climate change on phenology, frost damage, and floral abundance of montane wildflowers [J]. Ecology, 2008, 89(2): 353-362.
[42] HEGLAND S J, NIELSEN A, LAZARO A, et al. How does climate warming affect plant-pollinator interactions? [J]. Ecology Letters, 2009, 12(2): 184-195.
[43] XIA J, NIU S, CIAIS P, et al. Joint control of terrestrial gross primary productivity by plant phenology and physiology [J]. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(9): 2788-2793.
[44] HAGGERTY B P, GALLOWAY L F. As time goes by: Response of reproductive phenology to growing season length in American Bellflower, with implications for climate change [C]//ESA Conference Pape, 2009.
[45] TVERAA T, STIEN A, B RDSEN B, et al. Population densities, vegetation green-up, and plant productivity: impacts on reproductive success and juvenile body mass in reindeer [J]. Plos One, 2013, 8(2): e56450.
[46] ANDERSON M G, EMERY R B, ARNOLD T W. Reproductive success and female survival affect local population density of canvasbacks [J]. Journal of Wildlife Management, 1997, 61(4): 1174.
[47] JAEGER R. Competitive exclusion as a factor influencing the distributions of two species of terrestrial salamanders [J]. Ecology, 1971, 52(4): 632-637.
[48] XU Z F, HU T X, WANG K Y, et al. Short-term responses of phenology, shoot growth and leaf traits of four alpine shrubs in a timberline ecotone to simulated global warming, Eastern Tibetan Plateau, China [J]. Plant Species Biology, 2009, 24(1): 27-34.
[49] KLEIN J A, HARTE J and ZHAO X Q. Experimental warming, not grazing, decreases rangeland quality on the Tibetan Plateau [J]. Ecological Applications, 2007, 17(2): 541-557.
[50] SPIKE B P, TOLLEFSON J J. Relationship of plant phenology to corn yield loss resulting from western corn rootworm (Coleoptera: Chrysomelidae) larval injury, nitrogen deficiency, and high plant density [J]. Entomological Society of America, 1989, 82(1): 226-231.
[51] ZHOU S, ZHANG Y, CAYLOR K K, et al. Explaining inter-annual variability of gross primary productivity from plant phenology and physiology [J]. Agricultural & Forest Meteorology, 2016, 226/227: 246-256.
[52] MCKINNEY A M, CARADONNA P J, INOUYE D W, et al. Asynchronous changes in phenology of migrating broad-tailed hummingbirds and their early-season nectar resources [J]. Ecology, 2012, 93(9): 1987-1993.
[53] YLANNE H, STARK S, TOLVANEN A. Vegetation shift from deciduous to evergreen dwarf shrubs in response to selective herbivory offsets carbon losses: evidence from 19years of warming and simulated herbivory in the subarctic tundra [J]. Global Change Biology, 2015, 21(10): 3696-3711.
[54] 张亚生, 孙全平, 张华国. 试论西藏高原农事活动的“时宜”标志——物候[C]//第十六届中国科协年会——分5生态环境保护与绿色发展研讨会, 2014.
[55] 付刚, 钟志明. 西藏高原玉米物候和生态特征对增温响应的模拟试验研究[J]. 生态环境学报, 2016, 25(7): 1093-1097.
[56] 刘国一, 唐亚伟, 孙全平, 等. 提早播种对高原青稞增产的成因研究[J]. 西藏科技, 2015(8): 9-13.
[57] 肖薇薇. 气候变化对中国北方主要农作物物候影响及潜在适应性研究[D]. 北京: 中国科学院大学, 2014.
(编辑:段艳芳)
Changes of plant phenophases and their effects on the Qinghai-Tibetan Plateau
MENG Fandong①③, TSECHOE Dorji①②, CUI Shu juan①③, WANG Qi①③, LI Bowen①③, WANG Shiping①②③
①Institute of Tibetan Plateau Research, Chinese Academy of Sciences, Beijing 100101, China; ②CAS Center for Excellence in Tibetan Plateau Earth Sciences, Beijing 100101, China; ③University of Chinese Academy of Sciences, Beijing 100049, China
Plant phenology is the recurrence of life history events. Impacts of climate change and human activity on phenology were significant in the Qinghai-Tibetan Plateau, and warming and optimal grazing advanced the timings of green-up and flowering but delayed the end date of senescence. However, fruiting time kept relative stable compared with other phenophases. Warming prolonged the duration of plant activity period which was mainly derived from prolonged flowering duration as well as other reproductive phenophases. These changes mainly attributed to changes of temperature, moisture and grazing. Warming and wet, moderate grazing contributed to advance and lengthen phenophases, whereas warming and drought led to delay and shorten phenophases. Changes of phenophases had significant influences on structure and function of populations, community, ecosystem and productions and lives of pastoral and tourism. However, there were few studies on phenology, especially the effects of changes of phenology on structure and function of ecosystem on the Tibetan plateau. Therefore, we suggest that we should pay more attentions to effects of climate change and human activity on alpine plant phenology from physiology-level to ecosystem-level, and to feedbacks of phenology on ecosystems in the future.
Qinghai-Tibetan Plateau, climate change, grazing, plant phenology, effect
10.3969/j.issn.0253-9608.2017.03.005
*国家科技基础性工作专项重点项目(2012FY111400)、科技部气候变化重大专项(2013CB956000)、中国科学院战略性先导科技专项(B类)(XDB03030403)、国家自然科学基金重点和面上项目(41230750、31272488和31470524)资助
†通信作者,研究方向:高寒草地生态与气候变化和人类活动。E-mail: wangsp@itpcas.ac.cn
