气候变暖会使青藏高原树线一直上升吗?*
2017-07-06王亚锋梁尔源芦晓明朱海峰朴世龙朱立平
王亚锋,梁尔源②④†,芦晓明④,朱海峰②,朴世龙②③,朱立平②④
①中国科学院青藏高原研究所,北京 100101;②中国科学院青藏高原地球科学卓越创新中心,北京 100101;③北京大学城市与环境学院,北京 100871;④中国科学院大学,北京 100049
气候变暖会使青藏高原树线一直上升吗?*
王亚锋①,梁尔源①②④†,芦晓明①④,朱海峰①②,朴世龙①②③,朱立平①②④
①中国科学院青藏高原研究所,北京 100101;②中国科学院青藏高原地球科学卓越创新中心,北京 100101;③北京大学城市与环境学院,北京 100871;④中国科学院大学,北京 100049
树线上树木生长主要受生长季低温限制,因此气候变暖将导致高山树线向更高海拔迁移。然而,全球树线调查数据显示,近百年来约50%调查样点树线上升明显,其余则保持静止状态。以上事实揭示,除了气候因素,非气候因素(如竞争等种间关系)也在一定程度上调控着树线的位置变化。青藏高原拥有全球海拔最高的天然树线,是探讨气候和非气候因素对树线变化影响的理想区域。研究发现:树线之上低矮稀疏的灌丛或草丛成为幼苗萌发或生长的“避难所”,从而有利于树线大幅度爬升;而高大茂密的灌丛则会形成一道宽厚的“封锁墙”,从而抑制树线的上升。因此,变暖背景下树线位置倾向于向更高海拔爬升,但爬升速率受到树线之上种间关系的调控。变暖并不一定会造成树线位置的显著上升。
树线;青藏高原;气候变暖;种间关系;树轮
树线是指直立树木分布的海拔上限或纬度北界[1],通常被视为森林生态系统响应气候变化的潜在敏感区和预警区[2],在生态学和气候研究中备受关注。作为“地球的第三极”[3],青藏高原分布着北半球最高海拔的高山树线,是探讨树线对气候响应的理想场所[4-5]。目前全球变暖已经导致了青藏高原冰川显著退缩、雪线上升、湖泊大幅度扩张[3,6],然而,我们依然不清楚变暖背景下青藏高原的树线结构与格局是否发生了显著的变化,气候变暖是否已经导致了青藏高原树线位置的显著上升。基于样地调查和分析,本文旨在揭示青藏高原树线的时空变化格局,并利用统计模型探讨树线变化与生物或非生物因子之间的关系,为科学评估变暖对青藏高原森林生态系统的影响奠定基础。
1 树线类型以及形成机制
树线有多种分类方法。依据外观特征,树线可划分为四种类型:渐变型树线、急变型树线、树岛型树线和矮曲林树线[7]。若按成因划分,树线有三种基本类型:山地条件制约型树线、外界干扰(如人类活动、动物或昆虫的啃噬等)影响型树线和气候影响型树线[8-9]。由于对气候变化具有潜在的敏感性,气候影响型树线也是最受关注的树线类型。
历史上围绕气候树线形成了五类假说[2,10-11]。一是胁迫假说:频繁的冰冻和霜冻导致的干旱或强光辐射限制了树木的生长;二是干扰假说:风、雪压、雪崩、草本和真菌可能造成生物量或分生组织的损害,而低温下的树木生长发育不能弥补这些损失;三是碳平衡假说:碳的吸收和释放之间的平衡保持在较低的水平上,不足以维持最低生长的需要;四是繁殖假说:传粉、花粉管的生长、种子发育、种子传播、发芽和幼苗定居在严酷的树线环境下受限,从而阻碍了树木更新;五是生长限制假说:树线并非光合作用受限制导致的碳源不足,而是低温限制了碳源的利用而形成的。不过,这些假说都是基于温带地区的部分山地得出的结论,可能具有一定的片面性[2]。
早期的学者试图用某一特定值的等温线来解释气候树线成因。早期的一些研究认为,树线的位置与10 ℃等温线相重合[12]。后来的研究指出,10 ℃等温线仅仅是一种粗略估计,其有效覆盖范围在纬度40°~70°之间[10]。与中、高纬度树线温度相比,热带、亚热带树线温度较低,可以用3~6 ℃等温线来解释[10]。总体来说,全球树线位置可能落在5.5~7.5 ℃范围内[10]。更深入的监测研究揭示,全球树线(42°S~68°N)与6.7 ℃等温线相吻合[14]。在此基础上,学者提出了生长限制假说来解释全球树线形成。青藏高原气象监测数据支持以上结论[14-15],因此变暖背景下青藏高原树线可能会显著上升。
2 青藏高原树线变化的空间格局
基于树轮生态学和样地全样本调查法,在青藏高原东部调查了树线大样地 30m× (150~200)m 14个[16]。树线样地沿着纬度梯度设置,共涵盖了祁连山区、玉树三江源区、念青唐古拉山区、西藏昌都地区、云南横断山区,空间分布如图1所示。祁连山区树线海拔仅3 800 m左右,玉树三江源一带树线分布在4 200 m以上,而藏东南的树线可达4 900 m,成为北半球最高海拔的树线。由于位置偏僻、山高坡陡或深水阻隔等因素的影响,树线附近没有发现人为活动干扰的证据(砍伐、放牧、旅游等)。此外,树线之上存在不同高度和盖度的杜鹃灌丛或草地群落,暗示生物交互作用的存在[16-17]。多样的气候类型,复杂的地形地貌,不同的树种、生物交互作用的存在,以及缺乏人为干扰等,使青藏高原树线波动研究具有鲜明的区域特色。
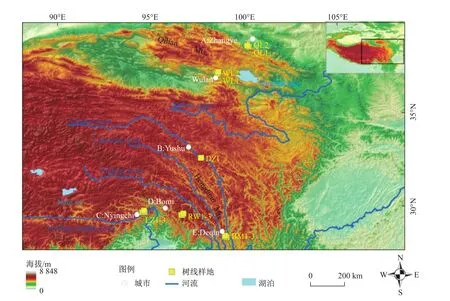
图1 青藏高原不同山区树线样地分布图[16]
一些学者在青藏高原个别山区开展过树线研究,但缺乏大空间尺度的系统研究。例如:通过比较不同历史时期的树线景观照片,美国学者发现云南横断山区树线在1923—2003年期间上升了67 m[18],而贡嘎山树线过去100年来仅发生了微小的变化[19];位于藏东的两条树线上升显著[20],而祁连东部三条树线在1891年以来却保持静止状况[21]。据此推测,青藏高原树线变化可能存在地区差异。
基于大空间尺度上的树线研究进一步证实,过去100年来,青藏高原不同地区的树线上升幅度呈现明显的空间差异性[16]。具体来说,祁连山北部边缘两块样地的树线分别上升了52 m和80 m,而祁连山中南部的两块样地的树线分别上升了13 m和54 m。类似的变化格局发生在西藏昌都然乌湖一带:一块样地的树线上升69 m,但其余两块样地的树线仅上升了4~ 6 m。玉树三江源一带一块样地的树线上升幅度为65 m。横断山区一块样地的树线上升幅度为19~28 m。此外,林芝地区色季拉山山区的三块样地树线上升幅度为0~0.8 m。总之,近百年来青藏高原不同山区的树线上升幅度为0~80 m。巧合的是,近百年来全球尺度上的树线变化也具有这种空间异质性特征[22],因此,探讨青藏高原树线变化的成因可为理解全球树线波动机制奠定基础。

图2 藏东南色季拉山急尖长苞冷杉树线之上茂密的杜鹃灌丛
3 青藏高原树线变化的驱动机制分析
过去100年来的气候变暖是千年尺度上的最暖期[23-25],但青藏高原树线并没有出现一致的、大幅度的爬升[16],暗示除了变暖以外,树线变化可能还受到非温度因素的影响。作为天然树线,可以排除人为干扰因素对青藏高原树线迁移的影响[5,16,26]。另外,研究所选取的树线多数位于和缓的山坡上,且树线以上地形平整、土壤发育良好,因此地形因子对树线迁移的影响并不显著[16]。那么,究竟是哪个非气候因子调控着树线上升的幅度呢?研究者猜测,林芝地区个别样点上树线的静止可能与树线之上茂密的高灌丛有关[16]。这为理解青藏高原大空间尺度上的树线变化机制提供了有益的线索(图2)。
藏东南色季拉山急尖长苞冷杉树线之上分布着茂密的高灌木群落,高度达3 m,冠层盖度在90%以上,形成了一道又宽又厚的“封锁墙”,种子落地于此便很快遭到“封杀”,从而导致树线长期处于相对静止(图2)[16]。与此相反,树线之上密度较小的矮灌丛群落能够为种子萌发和幼苗生长提供良好的“避难所”,这解释了祁连山、三江源、然乌湖、横断山区部分样点上树线的大幅度上升[16]。当树线之上的植被状况介于二者之间时,树线上升幅度则相对有限。以上场景展示了种间关系对树线上升幅度的影响。为进一步探讨二者之间的关系,定义一植被厚度指数TI(树线之上优势植被的高度×盖度),发现厚度指数的变化与树线变化幅度相当吻合(图3)。此外,当同时考虑种间关系、温度等多个因素时,预测模型能解释97%的树线变化。因此,变暖有利于树线位置的上升,但是树线上升幅度受到种间关系的调控。如果树线之上具有坚实的“封锁墙”,变暖并不会造成树线位置的显著上升。
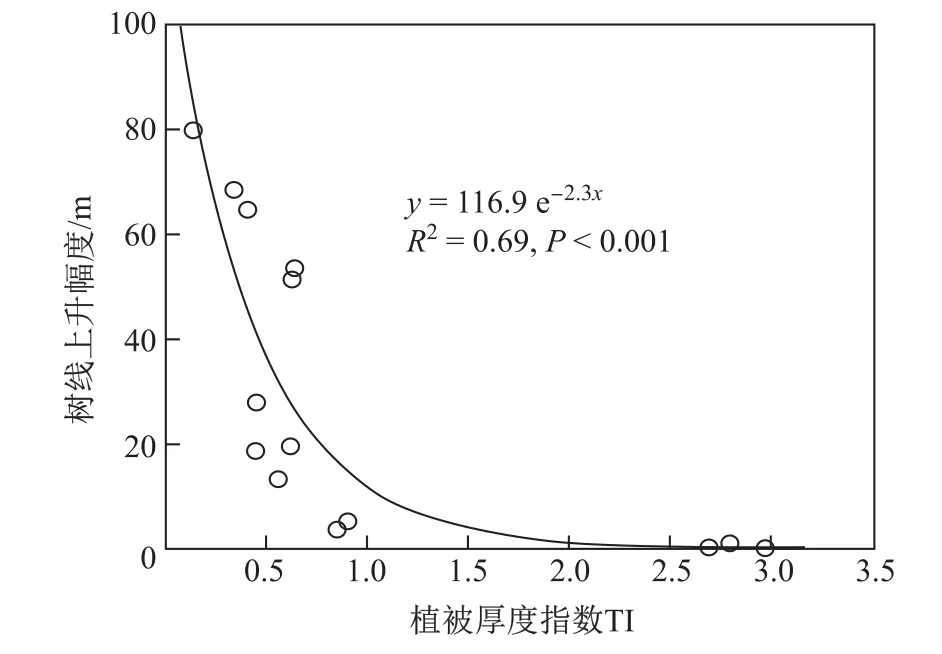
图3 树线之上植被厚度指数(TI)与树线上升幅度之间的关系[16]
4 结论
青藏高原树线上树木的生长和更新主要也受低温控制,由此预期气候变暖将导致青藏高原树线向更高海拔爬升。诸多研究结果证实了这一设想。尽管如此,青藏高原树线变化幅度仍存在显著的空间异质性,表明温度和非温度因素均能影响树线位置的变化。通过分析不同的生物、非生物因子对树线变化的相对贡献,发现气候变暖导致的树线上升会受到种间关系的调控。鉴于此,未来变暖情景下,树线上升速率在很大程度上由种间关系(例如互利、竞争)及其强度决定,变暖并不一定会造成树线位置的显著上升。
(2017年5月8日收稿)
[1] KULLMAN L. 20th century climate warming and tree-limit rise in the Southern Scandes of Sweden [J]. Ambio, 2001, 30(2): 72-80.
[2] 崔海亭, 刘鸿雁, 戴君虎, 等. 山地生态学与高山林线研究[M]. 北京:科学出版社, 2005.
[3] YAO T, THOMPSON L, YANG W, et al. Different glacier status with atmospheric circulations in Tibetan Plateau and surroundings [J]. Nature Climate Change, 2012, 2(9): 663-667.
[4] 李文华. 西藏森林[M]. 北京: 科学出版社, 1985.
[5] LIANG E, WANG Y, ECKSTEIN D, et al. Little change in the fi r treeline position on the southeastern Tibetan Plateau after 200 years of warming [J]. New Phytologist, 2011, 190(3): 760-769.
[6] ZHU L, XIE M, WU Y. Quantitative analysis of lake area variations and the influence factors from 1971-2004 in the Nam Co basin of the Tibetan Plateau [J]. Chinese Science Bulletin, 2010, 55(13): 1294-1303.
[7] HARSCH M A, BADER M Y. Treeline form—a potential key to understanding treeline dynamics [J]. Global Ecology and Biogeography, 2011, 20(4): 582-596.
[8] HOLTMEIER F K, BROLL G. Sensitivity and response of northern hemisphere altitudinal and polar treelines to environmental change at landscape and local scales [J]. Global Ecology and Biogeography, 2005, 14(5): 395-410.
[9] HERREO A, ZAMORA R, CASTRO J, et al. Limits of pine forest distribution at the treeline: herbivory matters [J]. Plant Ecology, 2012, 213(3): 459-469.
[10] KÖRNER C. A re-assessment of high elevation treeline positions and their explanation [J]. Oecologia, 1998, 115(4): 445-459.
[11] KÖRNER C. Alpine plant life: functional plant ecology of high mountain ecosystems [M]. Berlin: Springer, 2003.
[12] GRACE J. Plant response to wind [M]. London: Academic Press, 1977.
[13] KÖRNER C, PAULSEN J. A world-wide study of high altitude treeline temperatures [J]. Journal of Biogeography, 2004, 31(31): 713-732.
[14] SHI P, KÖRNER C, Hoch G. A test of the growth-limitation theory for alpine treeline formation in evergreen and deciduous taxa of the Eastern Himalayas [J]. Functional Ecology, 2008, 22(2): 213-220.
[15] LIU B, LIANG E, ZHU L. Microclimatic conditions for Juniperus saltuaria treeline in the Sygera Mountains, southeastern Tibetan Plateau [J]. Mountain Research and Development, 2011, 31(1): 45-53.
[16] LIANG E, WANG Y, PIAO S, et al. Species interactions slow warming-induced upward shifts of treelines on the Tibetan Plateau [J]. Proceedings of the National Academy of Sciences of the United States of America, 2016, 113(16): 4380-4385.
[17] WANG Y, CAMARERO J J, LUO T, et al. Spatial patterns of Smith fir alpine treelines on the south-eastern Tibetan Plateau support that contingent local conditions drive recent treeline patterns [J]. Plant Ecology & Diversity, 2012, 5(3): 311-321.
[18] BAKER B B, MOSELEY R K. Advancing treeline and retreating glaciers: implications for conservation in Yunnan, P R China [J]. Arctic, Antarctic and Alpine Research, 2007, 39(2): 200-209.
[19] 冉飞, 梁一鸣, 杨燕, 等. 贡嘎山雅家埂峨眉冷杉林线种群的时空动态[J]. 生态学报, 2014, 34(23): 6872-6878.
[20] LYU L, ZHANG Q, DENG X, et al. Fine-scale distribution of treeline trees and the nurse plant facilitation on the eastern Tibetan Plateau [J]. Ecological Indicators, 2016, 66: 251-258.
[21] GOU X, ZHANG F, DENG Y, et al. Patterns and dynamics of treeline response to climate change in the eastern Qilian Mountains, northwestern China [J]. Dendrochronologia, 2012, 30(2): 121-126.
[22] HARSCH M A, HULME P E, MCGLONE M S, et al. Are treelines advancing? A global meta-analysis of treeline response to climate warming [J]. Ecology Letters, 2009, 12(10): 1040-1049.
[23] LIU X, QIN D, SHAO X, et al. Temperature variations recovered fromtree-rings in the middle Qilian Mountain over the last millennium [J]. Science in China Series D: Earth Sciences, 2005, 48(4): 521-529.
[24] ZHU H, ZHENG Y, SHAO X, et al. Millennial temperature reconstruction based on tree-ring widths of Qilian juniper from Wulan, Qinghai Province, China [J]. Chinese Science Bulletin, 2008, 53(24): 3914-3920.
[25] THOMPSON L G, MOSLEY-THOMPSON E, BRECHER H, et al. Abrupt tropical climate change: past and present [J]. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(28): 10536-10543.
[26] 王亚锋, 梁尔源. 树线波动与气候变化研究进展[J]. 地球环境学报, 2012, 3(3): 855-861.
(编辑:沈美芳)
研究发现长非编码RNA调控RNA聚合酶I的转录功能
长度大于200个核苷酸的长非编码RNA,曾被认为是基因组转录的“暗物质”。然而,近年来的大量研究表明,长非编码RNA参与一系列细胞重要功能调控,如细胞核亚结构的形成、基因表达调控、表观遗传调控等。2017年5月5日的《细胞》杂志报道了长非编码RNA一种全新的功能:调控RNA聚合酶I的转录活性。
该研究由中国科学院生物化学与细胞生物学研究所陈玲玲研究组完成,首次在人类细胞中发现一种新的名为SLERT的长非编码RNA,可以调控细胞核仁功能和RNA聚合酶I转录功能,拓展了长非编码RNA的作用机制。
SLERT长为694个核苷酸,来源于人蛋白编码基因TBRG4内含子区域,定位于细胞核仁中。人类基因组中含有约400个拷贝的核糖体DNA(rDNA)。围绕rDNA区域可形成核仁——RNA聚合酶I转录核糖体RNA (rRNA) 以及rRNA加工的重要场所。作为细胞内含量最多的一类RNA,rRNA的转录失调与疾病发生密切相关,转录不足易导致骨髓衰竭性贫血,转录过多则易引发多种癌症。
在此项研究中,研究人员利用基因编缉技术精确敲除位于细胞核仁中的SLERT后发现,SLERT的缺失导致了RNA聚合酶I转录活性的降低。进一步研究发现,SLERT可以与RNA解旋酶DDX21结合。通过超高分辨率的结构照明显微技术,对DDX21在细胞核仁中的定位进行观察,首次发现DDX21在细胞核仁中围绕RNA聚合酶 I复合体形成直径约为400 nm的环状结构。而且,形成的这一环状结构与RNA聚合酶I转录偶联并抑制RNA聚合酶I转录。这个环状结构将RNA聚合酶I“围困”在其中,其“包围圈”大小直接影响RNA聚合酶I转录的活性。深入的研究表明,SLERT与DDX21的结合可以改变DDX21的蛋白构象,从而调整了DDX21环在细胞核仁中的规则排布,最终通过解除DDX21环对RNA聚合酶 I的抑制作用起到正调控RNA聚合酶I转录活性的作用。
这项研究阐释了细胞核仁中长非编码RNA SLERT、RNA解旋酶DDX21和RNA聚合酶 I复合体之间相互作用的分子机制,解析了DDX21环的大小对于RNA聚合酶I转录的调控机制以及SLERT对DDX21环的控制作用,以崭新的视角揭示了长非编码RNA的新机制,也为深入研究细胞核仁结构及功能提供了新方向。
[段艳芳 编译]
Are treelines advancing in response to climate warming on the Tibetan Plateau?
WANG Yafeng①, LIANG Eryuan①②④, LU Xiaoming①④, ZHU Haifeng①②, PIAO Shilong①②③, ZHU Liping①②④
① Institute of Tibetan Plateau Research, Chinese Academy of Sciences, Beijing 100101, China; ② CAS Center for Excellence in Tibetan Plateau Earth Sciences, Beijing 100101, China; ③ College of Urban and Environmental Sciences, Peking University, Beijing 100871, China; ④ University of Chinese Academy of Sciences, Beijing 100049, China
Tree growth at treelines is generally limited by low temperature so that warming tends to shift treeline upward. However, a global meta-analysis showed that treelines shifted upward at around 50% during the last century among the investigated treelines, and others kept static. Such evidence suggested that non-climate factors (such as interspecific competition) apart from climatic factors controlled treeline shift. The Tibetan Plateau hosts the highest treeline in the Northern Hemisphere, being a good locale for investigating climatic and biotic factors influencing treeline changes. As showed by our research, short and sparse shrub and grass acted as“tree nurseries”for recruitment, and benefitted treeline upward shift. Dense shrubs just above treeline inhibited tree establishment, and slowed upward movement of treelines. Climatic warming tended to promote the upward shift of alpine treelines at local and regional scales. However, upslope migration rates were controlled largely by interspecific interactions. Warming may not cause the upward shift of treelines if dense shrub dominate the above treelines.
treeline, Tibetan Plateau, climate warming, species interaction, tree ring
10.3969/j.issn.0253-9608.2017.03.004
*国家科技基础性工作专项重点项目(2012FY111400)和国家自然科学基金项目(41525001、41301207、41130529)资助
†通信作者,国家杰出青年科学基金获得者,研究方向:树轮生态学与气候学。E-mail: liangey@itpcas.ac.cn
