传代次数对人胶质瘤U87细胞系生物学特性的影响
2017-07-03王济舟漆松涛
王济舟,曾 宇,宋 烨,漆松涛
南方医科大学南方医院,广东 广州 510515
基础研究
传代次数对人胶质瘤U87细胞系生物学特性的影响
王济舟,曾 宇,宋 烨,漆松涛
南方医科大学南方医院,广东 广州 510515
目的探索不同传代次数对人胶质瘤U87细胞系生物学特性的影响及其分子机制。方法以两种不同传代次数的U87(I)、U87(II)为研究对象,使用MTT细胞增殖实验、Transwell小室迁移实验及Boyden小室侵袭实验分别检测U87细胞的增殖、迁移和侵袭能力;使用Western Blot技术检测U87(Ⅰ)及U87(Ⅱ)的CTHRC1, FOXM1, PLOD2, MMP9, TGF-β, E-cadherin,Slug, Snail, Vimentin, PI3K, p-PI3K, Akt, p-Akt的表达量差异。结果U87(Ⅰ)较U87(Ⅱ)更容易形成网状结构,有更强的侵袭能力,但在增殖和迁移能力上二者无明显差异。在EMT相关蛋白表达水平上,U87(Ⅰ)中的Snail、Vimentin的表达量较U87(Ⅱ)的高,而E-Cadherin、Slug则较低;在PI3K/Akt通路蛋白表达水平上,U87(Ⅰ)的p-Akt,Akt的表达量均低于U87(II),而p-PI3K的表达量却高于U87(II),两者在PI3K的表达量上无明显差异;除此之外,U87(I)中PLOD2、CTHRC1及MMP9的表达量也明显高于U87(II),而TGF-β的表达量则低于后者。结论随着传代次数的增加,U87细胞在侵袭能力以及多个促癌基因表达上发生了变化,这可能造成分子机制研究的前后结果不一致,降低结果的可信程度,因此研究人员在进行细胞系实验时,应尽可能在短时间内,使用同一批次细胞完成生物学特性、分子机制研究,以减少传代次数对肿瘤细胞系生物学特性以及基因表达的影响。
胶质瘤;U87;传代次数;侵袭能力
稳定的肿瘤细胞系是如今科学研究人员进行肿瘤研究的重要工具[1-5],细胞系的永生特性使得研究者可以不断的深入探索该细胞系所代表的肿瘤的发生、发展机制,从而为肿瘤的诊断、治疗及预后找到新的突破口。然而实验者在操作细胞时极为容易将细胞弄混,许多研究者便因混淆细胞系而遭遇撤稿[6-7]。此外,Lin等[8]研究发现,不同代数的前列腺癌细胞之间存在异质性,说明在不断的传代过程中,细胞的生物学特性及基因表达都有可能出现变化。人胶质瘤U87细胞系建立于1966年,是由瑞典乌普萨拉大学的研究人员利用一位44岁的胶质母细胞瘤女患者的组织进行培养得到的,现已是全球各地研究人员研究胶质瘤的最常用的细胞系之一[9]。这个细胞系的广泛使用为广大研究人员提供了便利,但现有研究中却鲜有研究者研究不同传代次数的U87细胞的异同,以及明确述及他们的实验是否是在相同传代次数的U87细胞中进行,故而这些实验结果可靠性都是值得推敲的。本实验旨在探索不同传代次数U87细胞的分子表型及生物学功能差异,以明确在相同传代次数的U87细胞中进行分子生物学实验的必要性。
1 方法
1.1 细胞培养
人胶质瘤U87细胞系购自中国科学院上海生命科学研究院细胞库。U87细胞采用10%胎牛血清(Gemini)的DMEM培养基,在37 ℃、5% CO2、饱和湿度的条件下传代培养,待细胞生长状态良好时用于实验。
1.2 Western blot
Western blot的操作流程及具体方法[10], 本实验中使用的抗体包括anti-CTHRC1, FOXM1, PLOD2,MMP9, TGF-β, E-cadherin, Slug, Snail, Vimentin,PI3K, p-PI3K, Akt, p-Akt, β-actin, GAPDH。具体的抗体见补充表格。所有图片均为Bio-Rad公司的化学发光仪拍摄获得,并使用Image-Pro Plus 6.0测量所获条带灰度值。
1.3 MTT细胞增殖实验
本实验使用MTT实验检测细胞增殖能力[11]。将U87细胞铺于96孔板的孔中(1000个/孔,200 μL/孔),每种细胞均设5个复孔。细胞贴壁后加入20 μL MTT(5 mg/mL稀释于PBS中, Sigma, St Louis, MO),4 h后吸出培养基,使用150 μLDMSO(Sigma, St Louis,MO)溶解甲瓒结晶,并用酶标仪测量490 nm的吸光度。此后每天同一时间加入MTT及测量吸光度,共观察7 d。
1.4 细胞迁移和侵袭实验
细胞迁移及侵袭实验方法同我们此前发表文章中所述[12]。对于细胞迁移实验,我们将1×105细胞用100 μL无血清培养基重悬,铺入小室中,2 h 30 min后使用甲醇固定,并用Giemsa染色液染色。对于细胞侵袭实验,我们首先在小室中铺入100 μL的基质胶(康宁)(250 μg/mL),在37 ℃、5% CO2孵箱中静置4 h后再铺入细胞,具体步骤同细胞迁移实验。统计时选取200 x视野,在四个视野中计数穿过膜的细胞数并取平均值。
1.5 数据分析
使用SPSS 21.0统计学软件进行数据分析,数据采用均数±标准差表示。细胞迁移、侵袭实验以及Western灰度值结果均采用非配对样本t检验,P<0.05为差异具有统计学意义。
2 结果
2.1 传代次数不同的细胞的形态及生长特性不同
U87(Ⅰ)及U87(Ⅱ)为不同传代次数的两批细胞,其中U87(Ⅱ)的传代次数较多,二者的传代次数差异>100代。将相同数量的U87(Ⅰ)及U87(Ⅱ)铺入10 cm的培养皿中,7 h后于镜下观察二者的形态及密度,两种细胞的密度无明显差异,但U87(Ⅰ)的外观较U87(Ⅱ)更加饱满,体积稍大。4 d后再次观察两种细胞,外观出现了明显的差异。U87(Ⅰ)形成了明显的较为规则的网状结构,网状空隙较大,空隙中无贴壁细胞,U87(Ⅱ)虽然也形成了一定的网状结构,但细胞的排列较为散乱,所形成的网状空隙较小(图1)。
2.2 传代次数不同的细胞的侵袭能力不同
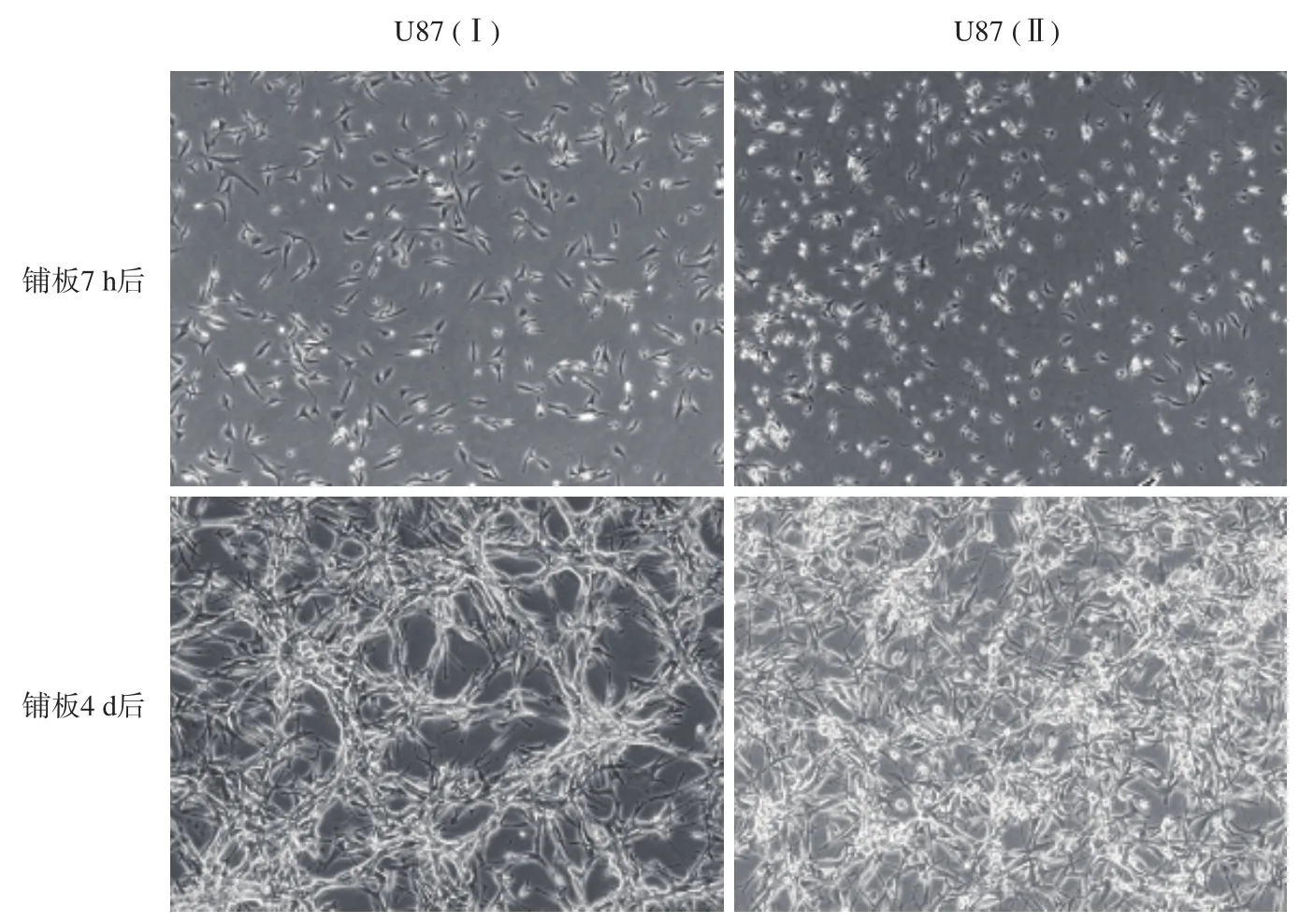
图1 U87(Ⅰ)与U87(Ⅱ)在镜下的形态差异
为探究U87(Ⅰ)及U87(Ⅱ)的生物学特性的差异,我们进行了MTT实验、Transwell小室迁移实验及Boyden小室侵袭实验分别检测两者的增殖、迁移、侵袭能力,3种实验均独立重复了3次,Transwell小室迁移实验及Boyden小室侵袭实验的结果为4个不同视野的平均值。MTT实验显示,两种细胞在增殖能力上无明显差异(图2)。Transwell实验显示,U87(Ⅰ)的迁移能力较U87(Ⅱ)稍强,但无统计学差异(P=0.1169)。而Boyden实验显示U87(Ⅰ)较U87(Ⅱ)的侵袭能力更强,有统计学差异(P=0.0403, 图3)。

图2 U87(Ⅰ)和U87(Ⅱ)之间增殖能力的差异
2.3 传代次数不同的细胞的蛋白表达量存在差异
为进一步研究U87(Ⅰ)及U87(Ⅱ)出现侵袭能力差异的分子机制,本研究使用Western blot分析两种细胞中EMT相关蛋白、PI3K/Akt信号通路相关蛋白及部分促癌基因的蛋白的表达水平。研究发现,在EMT相关的蛋白表达量上,E-ca及Slug在U87(Ⅰ)中表达量较低,而Snail、Vimentin在U87(Ⅰ)中表达量较高;在PI3K/Akt通路上,Akt及p-Akt在87(Ⅰ)的表达量均低于U87(Ⅱ),但p-PI3K的表达量却高于后者,而PI3K的表达量在两个细胞系之间无明显差异。此外,U87(Ⅰ)的MMP9、CTHRC1、PLOD2表达量明显高于U87(Ⅱ),而TGF-β的表达量则较U87(Ⅱ)的低,FOXM1的表达量无明显差异(图4)。
3 讨论
U87是目前胶质瘤研究常用的稳定细胞系,然而本研究团队在培养U87细胞的过程中发现细胞的形态及生长特性随传代次数的增加发生了变化,传代次数较少的U87比较饱满,细胞较大,易于结网,在传代3~4 d后出现明显的网状结构,细胞会在网状的骨架中增殖,而不会向骨架中的空隙中生长,而连续传代半年后的U87细胞体积相对较小,不易结网。既往的研究表明,恶性程度高的胶质瘤细胞系如U251、U87容易形成网状结构,Francescone等[13]认为这种网状结构是胶质瘤细胞血管生成拟态的微观表现,是其高度恶性的表现之一。在此基础上,本研究深入探究了形态相异的同种细胞系、不同传代次数的细胞之间的生物学特性及蛋白表达差异。结果显示,二者的生长和迁移能力无明显差别,但传代次数较少的U87的侵袭能力要强于传代次数多的U87。而WB的结果则反映出二者在数个与肿瘤生物学行为相关的基因在蛋白水平的表达量存在明显的不同。本研究在前期研究中发现PLOD2及CTHRC1高表达可以促进U87细胞的迁移和侵袭能力,并且PLOD2在缺氧状态下可促进U87的生存。此外,MMP9为基质金属蛋白酶家族的成员之一,许多研究证实MMP9可以促进胶质瘤的迁移、侵袭能力[14-15]。本实验发现,传代次数少的U87的PLOD2、CTHRC1及MMP9表达明显高于传代次数多的U87,这可能可以解释为何前者较后者拥有更强的侵袭能力。

图3 U87(Ⅰ)和U87(Ⅱ)之间迁移、侵袭能力的差异
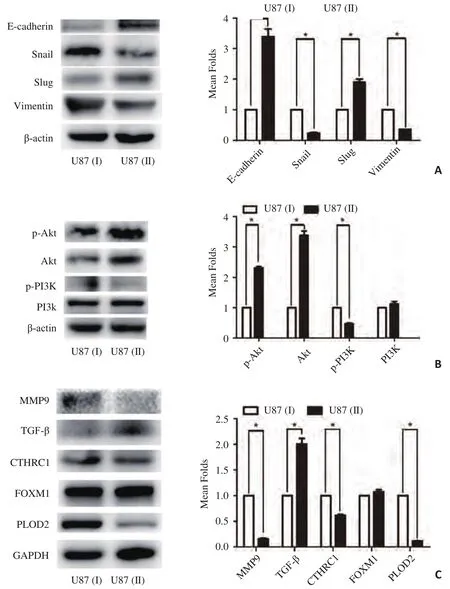
图4 U87(Ⅰ)和U87(Ⅱ)之间多种促癌基因表达量在蛋白水平上的差异
细胞系在传代过程中可能出现不同细胞系交叉污染或者标记错误的情况,许多研究者因发现前期实验使用了错误的细胞系而遭遇撤稿[6-7]。2016年Allen等[16]报道称他们通过DNA指纹技术发现ATCC细胞库所使用的U87与50年前建立的原始肿瘤样本并不匹配。本实验仅使用了U87及U251两种细胞系,从细胞形态可看出,U87(Ⅰ)与U87(Ⅱ)形态相近,且与U251形态明显不同。为排除U87被U251污染的情况,研究对比了U251与U87(Ⅰ)之间部分蛋白表达量的差异。在PLOD2及CTHRC1的表达含量上,二者无显著差异,故而可以推测,U87(Ⅰ)与U87(Ⅱ)所出现的蛋白表达含量差异并不是U251污染所致,而更可能是传代过程中出现的基因突变、表观遗传学修饰等因素所致。
不同传代次数影响细胞的迁移、侵袭能力,同时改变了癌基因或抑癌基因的表达,这可能有两方面原因,一方面细胞本身存在异质性,通过优胜劣汰逐渐改变细胞的整体特性,另一方面,在培养过程中,不断有外界的刺激或者干扰,如支原体污染,更换血清品牌,消化传代等物理、化学因素,而对细胞的基因表达产生影响,继而出现突变。在实验操作中,采用液氮冻存的方法保种,在培养细胞出现污染、凋亡或者用尽后,往往采取复苏细胞,重复进行实验,而这样的操作可能因不同批次的细胞之间的差异,造成前后实验结果不相符,增加数据分析的复杂性。因此,研究人员在使用肿瘤细胞系进行实验的过程中,应该尽量保证使用同一批细胞,并且在短时间内完成分子机制、功能的相关研究,减少因传代次数造成的细胞异质性所带来的影响。
此外,随着传代次数的增加及体外培养条件的改变,肿瘤细胞系的分子特性与其最初在体内时有所差别,目前主流研究多用原位成瘤实验或者皮下成瘤实验模拟肿瘤在体内的生物学过程[17-18],但如果实验时肿瘤细胞系的分子特性已经出现改变,成瘤实验并不能确切地反映出肿瘤在病人体内发生发展的真实情况。故而肿瘤研究应不仅仅使用稳定的细胞系,也应重视原代细胞培养与使用。当原代细胞能够相对稳定的传代后,通过培养单克隆的原代细胞,建立新的传代次数较少的细胞系,然后在该细胞系中验证分子机制,这样可以尽可能地模拟肿瘤在病人体内时的生物学特性,增加实验结果的可信程度。
[1]Barretina J, Caponigro G, Stransky N, et al. The cancer cell line encyclopedia enables predictive modelling of anticancer drug sensitivity[J]. Nature, 2012, 483(7391): 603-7.
[2]Haverty PM, Lin E, Tan J, et al. Reproducible pharmacogenomic profiling of cancer cell line panels[J]. Nature, 2016, 533(763): 333-7.
[3]Cancer cell line encyclopedia consortium, genomics of drug sensitivity in cancer consortium. pharmacogenomic agreement between two cancer cell line data sets[J]. Nature, 2015, 528(7580):84-7.
[4]Whitington T, Gao P, Song W, et al. Gene regulatory mechanisms underpinning prostate cancer susceptibility[J]. Nat Genet, 2016,48(4): 387-97.
[5]Que T, Song Y, Liu Z, et al. Decreased miRNA-637 is an unfavorable prognosis marker and promotes glioma cell growth,migration and invasion via direct targeting Akt1[J]. Oncogene,2015, 34(38): 4952-63.
[6]He YC, Chen FC, Cai Y. Retracted: knockdown of tumor protein D52-like 2 induces cell growth inhibition and apoptosis in oral squamous cell carcinoma[J]. Cell Biol Int, 2016, 40(3): 361-4.
[7]Zhang P, Yang Y, Zweidler-Mckay P, et al. Retraction: critical role of notch signaling in osteosarcoma invasion and metastasis[J]. Clin Cancer Res, 2013, 19(18): 5256-7.
[8]Lin HK, Hu YC, Yang L, et al. Suppression versus induction of androgen receptor functions by the phosphatidylinositol 3-kinase/Akt pathway in prostate cancer LNCaP cells with different passage numbers[J]. J Biol Chem, 2003, 278(51): 50902-7.
[9]Dolgin E. Venerable brain-cancer cell line faces identity crisis[J].Nature, 2016, 537(7619): 149-50.
[10]Song Y, Luo Q, Long H, et al. Alpha-enolase as a potential cancer prognostic marker promotes cell growth, migration, and invasion in glioma[J]. Mol Cancer, 2014, 13(5): 65-7.
[11]Song Y, Hu Z, Long H, et al. A complex mechanism for HDGF-mediated cell growth, migration, invasion, and TMZ chemosensitivity in glioma[J]. J Neurooncol, 2014, 119(2):285-95.
[12]Qi S, Song Y, Peng Y, et al. ZEB2 mediates multiple pathways regulating cell proliferation, migration, invasion, and apoptosis in glioma[J]. PLoS One, 2012, 7(6): e38842-4.
[13]Francescone R, Scully S, Bentley B, et al. Glioblastoma-derived tumor cells induce vasculogenic mimicry through Flk-1 protein activation[J]. J Biol Chem, 2012, 287(29): 24821-31.
[14]Joseph JV, van Roosmalen IA, Busschers E, et al. Serum-Induced differentiation of glioblastoma neurospheres leads to enhanced migration/invasion capacity that is associated with increased MMP9[J]. PLoS One, 2015, 10(12): e0145393-5.
[15]Wang F, Xiao W, Sun J, et al. MiRNA-181c inhibits EGFR-signaling-dependent MMP9 activation via suppressing Akt phosphorylation in glioblastoma[J]. Tumour Biol, 2014, 35(9):8653-8.
[16]Allen M, Bjerke M, Edlund H, et al. Origin of the U87MG glioma cell line: Good news and bad news[J]. Sci Transl Med, 2016,8(354): 354re3-5.
[17]Depner C, Zum BH, Böğürcü N, et al. EphrinB2 repression through ZEB2 mediates tumour invasion and anti-angiogenic resistance[J].Nat Commun, 2016, 7(4): 12329-33.
[18]Kim D, Fiske BP, Birsoy K, et al. SHMT2 drives glioma cell survival in ischaemia but imposes a dependence on glycine clearance[J]. Nature, 2015, 520(7547): 363-7.
Effect of different passage number on the biological characteristics of U87 glioblastoma cell line
WANG Jizhou, ZENG Yu, SONG Ye, QI Songtao
Department of Neurosurgery, Nanfang Hospital, Southern Medical University, Guangzhou 510515, China
ObjectiveTo unravel the differences of biological characteristics and the uderlying molecular mechanisms between different passages of U87 glioblastoma cell line.MethodsU87 (I) and U87 (II), with less or more passage number respectively,were established examined separately by MTT assay, transwell chamber assay, boyden chamber assay. Western blot was used to analyze the expression of CTHRC1, FOXM1, PLOD2, MMP9, TGF-β, E-cadherin, Slug, Snail, Vimentin, PI3K, p-PI3K, Akt and p-Akt separately in these two types of cells.ResultsCompared to U87 (II), U87 (I) was more prone to form tubules and showed more invasiveness, while there were no differences between U87 (I) and U87 (II) in proliferation and migration. The expression of markers of EMT varies in these two types of cells, with more expression of E-cadherin and Slug and less expression of Vimentin and Snail in U87 (II). Moreover, U87 (II) was found to have greater amount of Akt and p-Akt, but have smaller amount of p-PI3k. No significant difference was found on the expression of PI3K between these cells. Additionally, the expression of PLO2, CTHRC1 and MMP9 were higher in U87 (I) than that in U87 (II), while the expression of TGF-β in U87 (I)was lower than that in the latter one.ConclusionAs the passage number increased, U87 cell lines exhibited changes in invasiveness as well as some oncogenic genes’ expressions. It may lead to the different results during different periods, making the results inconvincible. Together, our results showed that, to lessen the influence of passage numbers on cells’ biological characteristics, scientists should use cells with same passage numbers and finish the experiments as quickly as they can.
glioma; U87; passage number; invasiveness
2017-02-06
国家自然科学基金(81502178)
王济舟,博士研究生,E-mail: spiderpy@163.com;
漆松涛,主任医师,教授,博士生导师,E-mail: qisongtaonfyy@126.com
