黑麦草幼苗对冻融及酸沉降的生理响应特征
2017-06-30李晨包国章成文革李子勇李
李晨++包国章++成文革++李子勇++李艳芳
摘要:以黑麥草品种冬牧70为材料,研究冻融及pH值4.5模拟酸沉降条件对黑麦草幼苗体内抗氧化酶活性和渗透调节物质含量的影响。结果表明,在冻融胁迫下,黑麦草幼苗体内可溶性蛋白、可溶性糖、丙二醛(MDA)含量呈上升趋势,超氧化物歧化酶(SOD)、过氧化物酶(POD)活性明显提升,最高值分别为 107.35、6.23 U/mg;在pH值4.5酸沉降胁迫下,黑麦草幼苗体内可溶性蛋白含量逐渐降低,在-2 ℃时降到最低点,下降了20.37%,可溶性糖、丙二醛含量随温度下降而降低,最低值分别为10.42 mg/g、17.61 μmol/g;冻融与酸沉降复合胁迫处理对黑麦草幼苗的作用高于单一胁迫的作用效果,黑麦草幼苗体内可溶性蛋白、丙二醛含量、POD活性呈上升趋势,可溶性糖含量、SOD活性分别下降。试验结果表明,黑麦草幼苗对冻融和酸沉降逆境胁迫存在着明显的相互适应现象,对冻融及酸沉降型伤害有很强的适应性,且有一定的耐酸性和耐寒性。
关键词:黑麦草幼苗;冻融胁迫;酸沉降胁迫;生理特性
中图分类号: S181文献标志码: A文章编号:1002-1302(2017)08-0159-04
二年生黑麦草(ryegrass)是我国重要的牧草和草坪草之一,广泛分布于温带地区,具有抗寒性强、覆盖能力强、返青早、抗病虫害能力强等特性[1-2]。黑麦草富含蛋白质、矿物质及维生素,其经济利用价值较高。冬牧70黑麦草是从美国引进的越年生禾本科植物,是我国北方重要的牧草和草坪草之一,于20世纪90年代初引入我国,种植冬牧70黑麦牧草是改善我国北方农区春季饲草严重不足现状的可行途径之一,具有巨大的经济效益和社会效益。
冻融是指气候的日、年和多年变化导致特定气候区域地球表层一定范围的土冻结和融化作用,地球上受冻融作用的面积约占全球陆地总面积的70%,我国受冻融作用的面积约占国土陆地总面积的98%。冻融过程引起的环境改变对植物生长发育产生一系列直接或间接影响,使植被形态特征和生理功能等方面形成了特殊的适应性。揭示黑麦草对逆境环境的适应机制是培养抗寒性强的黑麦草品种的关键,这有助于采取有效的措施减轻低温对植物造成的危害。
酸沉降胁迫对植物的生长发育会产生严重不利影响,这已成为全球性环境污染问题之一。研究黑麦草幼苗对模拟酸沉降生理指标的影响,可以了解黑麦草抗酸沉降的能力,为研究植物对酸沉降胁迫适应能力提供理论依据。
过氧化物酶(POD)和超氧化物歧化酶(SOD)是清除活性氧的关键酶,其活性的高低反映了植物抗逆性的强弱,植物能通过应激性反应,激活抗氧化剂和诱导抗氧化酶的活性,减弱膜脂过氧化作用,保持膜的稳定性[3]。丙二醛(MDA)从膜上产生的位置释放出后,可与蛋白质、核酸反应,引起膜蛋白与膜内脂的变化,从而改变细胞膜系统。可溶性糖作为渗透调节物质和信号物质,可减缓由胁迫造成的生理代谢不平衡。
为此,本研究将通过冻融及酸沉降胁迫的方法,测定黑麦草幼苗体内可溶性蛋白含量、MDA含量、POD活性、SOD活性和可溶性糖含量这些生理指标在逆境胁迫中的变化和恢复能力,为揭示牧草和草坪草抗冻能力及防治酸沉降对其危害提供理论依据。
1材料与方法
1.1试材培养
试验于2015年9月在吉林大学环境与资源学院室内实验室进行。供试材料为冬牧70黑麦草(ryegrass),由吉林省农业科学院种子部门提供。
挑选籽粒饱满、大小均匀的种子,用0.1% KMnO4溶液进行消毒,溶液消毒2 h后取出并以矿泉水冲洗干净备用。将种子整齐排放在铺有2层滤纸的长方形托盘(30 cm×40 cm)内,放于SPX-300光照培养箱中进行培养,其温度设定为25 ℃,每天光照12 h,加水4次,保持滤纸湿润。培养 7 d 待幼苗长至16 cm时,选取长势一致的幼苗用于试验。
1.2模拟酸沉降母液的配制
根据吉林省长春市天然降水pH值及离子水平,依据文献[4]方法配制模拟酸沉降母液,考虑天然降水离子成分的复杂性,选用矿泉水配制模拟酸沉降的原溶液。用浓硫酸和浓硝酸(SO42- ∶NO3-=5 ∶1)配制pH值为4.5的酸沉降母液,用PHS-3C型雷磁酸度计测定与校核。
1.3试验方法
1.3.1人工冻融试验把试验中需要冻融胁迫的黑麦草幼苗样品标记好后进行融冻—冻融处理。融冻试验处理为每2 h降温6 ℃,冻融试验为每2 h升温6 ℃。试验的温度梯度设计为10、4、-2、4、10 ℃,融冻温度阶段为10 ℃—-2 ℃,冻融温度阶段为-2 ℃—10 ℃。每2 h变温随即取样,重复取6份样本进行指标测定。
1.3.2模拟酸沉降试验取1盘铺放均匀的黑麦草种子,标记为酸沉降组进行试验。培育7 d,待种子长出幼苗后,对酸沉降组均匀滴加50 mL酸沉降溶液。滴加时间为09:00,放置24 h后,进行指标测定。每2 h取样1次,重复抽取6份样本进行试验测定。
1.4测定生理指标
硫代巴比妥酸(TBA)比色法测定丙二醛、可溶性糖含量[5];考马斯亮蓝法测定可溶性蛋白质含量[5];SOD、POD活性均用南京建成生物工程研究所生产的试剂盒测定,用分光光度计进行测定。每个指标均设6组平行样。
1.5数据处理
采用Origin Pro 8.5绘制图表,用统计软件SPSS 16.0进行相关数据分析,图中数据均为6次重复测定值的平均值。
2结果与分析
2.1冻融及酸沉降胁迫对黑麦草幼苗体内蛋白质含量的影响
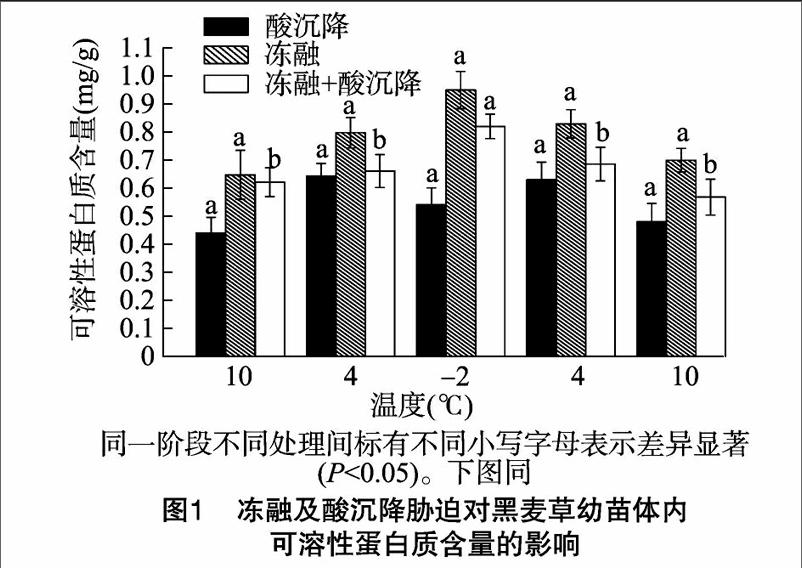
如图1所示,与冻融+酸沉降复合胁迫相比,经冻融胁迫的黑麦草幼苗体内蛋白质含量高于复合胁迫;在融冻阶段,当温度下降到-2 ℃时,黑麦草幼苗体内可溶性蛋白含量达到最高值,为0.95 mg/g,比复合胁迫下的蛋白质含量增加了15.83%;在冻融阶段,温度回升到10 ℃时,可溶性蛋白含量逐渐下降,复合胁迫组、冻融组蛋白质含量分别为 0.57、0.70 mg/g。这说明低温胁迫可使黑麦草体内积累渗透调节物质,增加了其可溶性蛋白含量,黑麦草自身的抗寒性可提高细胞的保水能力,保护生物膜及防止活性氧对膜脂和膜蛋白的氧化作用[6]。在融冻—冻融胁迫下,黑麦草幼苗体内抗逆蛋白质合成量快速增加,使束缚水相对增多,降低了幼苗体内细胞结冰而致死的概率,起到一定的抗寒作用[7]。经pH值4.5酸沉降胁迫处理后的黑麦草幼苗体内蛋白质含量先升高随后下降,在温度为-2 ℃时,降到最低值0.54 mg/g,在4 ℃时,达到最高值,为0.64 mg/g。本试验中,酸沉降胁迫下的黑麦草幼苗体内可溶性蛋白含量低于复合胁迫(P<0.05),降幅为3.03%~29.03%。这是由于在酸沉降胁迫下黑麦草体内的蛋白质遭到破坏,产生逆境胁迫生理效应,从而诱发幼苗体内自由基反应,破坏细胞膜结构的完整性,导致黑麦草体内细胞代谢紊乱,这与黄开志关于模拟酸雨对蔬菜细胞透性的影响研究结果[8]一致。黑麦草幼苗经冻融及酸沉降复合胁迫处理后,可溶性蛋白质含量随温度降低而升高,在温度为-2 ℃时上升到最高值,为0.82 mg/g。与经酸沉降胁迫的黑麦草幼苗相比,经冻融处理的黑麦草幼苗可溶性蛋白质含量均明显升高。
2.2凍融及酸沉降胁迫对黑麦草幼苗体内可溶性糖含量的影响
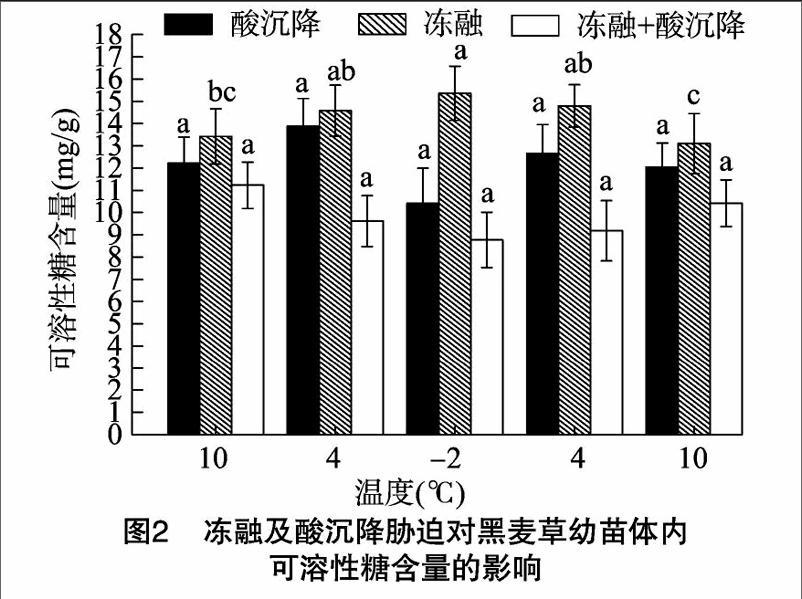
在pH值4.5酸沉降胁迫下,与冻融+酸沉降复合胁迫下相比,黑麦草幼苗可溶性糖含量明显升高,上升了 27.54%~44.49%;在温度为4 ℃时,达到最高值,为 13.9 mg/g,随着温度的逐渐下降,在-2 ℃时,可溶性糖含量降到最低值,为10.42 mg/g(图2)。这表明酸沉降不仅严重影响黑麦草叶片对氮素的吸收和利用,也阻碍了茎叶中的可溶性糖向根系运输,造成幼苗生长速率下降、叶片变黄,表现出明显的伤害症状,这与童贯和等关于小麦幼苗体内可溶性糖的影响研究结果[9]一致。在冻融胁迫下,与复合胁迫相比,黑麦草幼苗体内可溶性糖含量随温度降低而显著升高,在-2 ℃时达到最高值,为15.76 mg/g,在温度为10 ℃时降到最低值,为13.10 mg/g,降低了16.88%。本试验结果显示,冻融胁迫下黑麦草幼苗体内的可溶性糖类含量呈上升趋势,这表明冻融胁迫能对植物溶质的积累和渗透起到调节作用[10]。在冻融+酸沉降复合胁迫条件下,黑麦草幼苗体内可溶性糖含量随温度下降而降低,在温度降到最低点-2 ℃时,得到最小值8.77 mg/g。分析可得,低温胁迫下黑麦草幼苗体内会大幅度地积累可溶性糖类,其抗寒性也随之增强。
2.3冻融及酸沉降胁迫对黑麦草幼苗体内MDA含量的影响
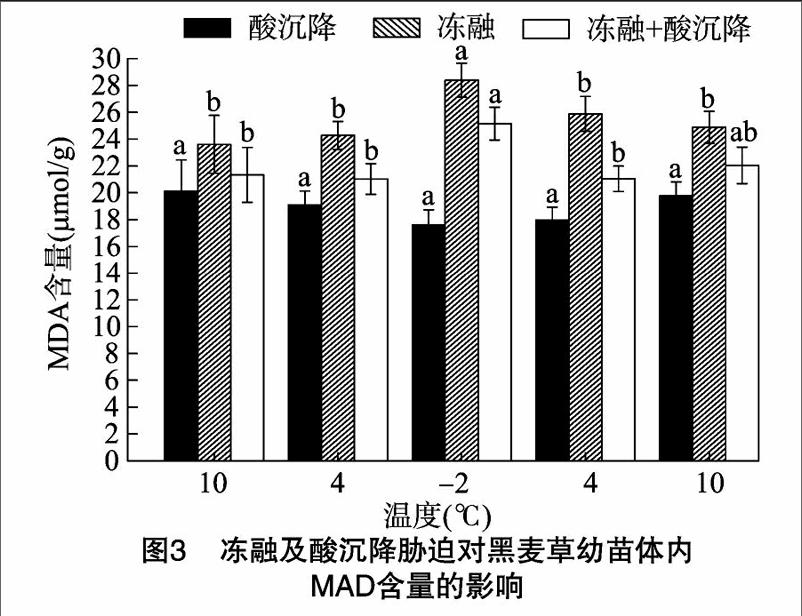
由图3可知,在冻融胁迫下,黑麦草幼苗体内MDA含量高于冻融+酸沉降复合胁迫下的含量,增加10.74%~12.88%;在融冻阶段,温度为-2 ℃时,幼苗体内MDA含量达到最高值,为28.39 μmol/g;在冻融阶段,温度在10 ℃时,MDA含量最低,为22.62 μmol/g。这表明在冻融胁迫下,黑麦草幼苗细胞膜系统受到损害,产生高活性的过氧化产物,使黑麦草幼苗体内MDA含量逐渐上升。这与宋采博等关于不同温度对白三叶幼苗生长的影响研究结果[11]相一致。经pH值4.5酸沉降胁迫处理后,与复合胁迫处理相比,黑麦草幼苗体内MDA含量随温度降低而下降,下降了10.16%~4282%(P<0.05),在-2 ℃时达到最低值 17.61 μmol/g(图3),这表明酸沉降胁迫影响黑麦草幼苗体内MDA的分泌,且随着酸胁迫的加深MDA含量增加,直接导致叶片膜结构的损害[12]。经冻融+酸沉降复合胁迫处理后,黑麦草幼苗体内MDA含量随温度下降而升高,在温度为-2 ℃时,达到最高值25.15 μmol/g(图3)。上述分析显示,在单一冻融胁迫下,黑麦草幼苗体内MDA含量随温度降低而升高,但在pH值4.5的酸沉降胁迫下其体内MDA含量明显降低。这说明酸沉降可降低黑麦草幼苗体内MDA含量,减少冻融对黑麦草幼苗的损伤,增强了黑麦草幼苗自身的抗寒性。
2.4冻融及酸沉降胁迫对黑麦草幼苗体内SOD活性的影响
在冻融胁迫条件下,与冻融+酸沉降复合胁迫相比,黑麦草幼苗体内SOD活性提高,在温度为-2 ℃时,SOD活性达到最高值107.35 U/mg,提升了 37.12%(图4)。SOD活性升高说明黑麦草自身抗寒性变强,膜伤害程度变低。SOD可有效地清除过多的自由基而抑制过氧化作用,能及时清除幼苗体内的有害物质,保护黑麦草自身不受损伤[12]。与经复合胁迫处理的幼苗相比,经pH值4.5酸沉降胁迫的幼苗体内SOD活性较高,其活性趋势先降低,随后升高,在10 ℃时,SOD活性达到最高值,为122.51U/mg,在温度为4℃时,
SOD活性降到最低点,为93.88 U/mg(图4)。这与吕家根等关于酸雨胁迫下小麦微弱延迟发光及其生理变化的研究结果[13]一致。在冻融+酸沉降复合胁迫条件下,黑麦草幼苗体内SOD活性随温度下降而降低,在温度为-2 ℃时降到最低点,为78.29 U/mg(图4)。这表明酸沉降胁迫能增强黑麦草幼苗体内SOD活性,逆境胁迫可以促进活性氧产生,加强膜脂氧化作用,平衡黑麦草体内活性氧。
2.5冻融及酸沉降胁迫对黑麦草幼苗体内POD活性的影响
如图5所示,与冻融+酸沉降复合胁迫相比,在冻融胁迫条件下的黑麦草幼苗体内POD活性降低,下降了11.71%~20.67%;在融冻阶段,幼苗体内POD活性随温度下降而升高,在-2 ℃时,达到最高值6.23 U/mg;在冻融阶段,幼苗体内POD活性随温度升高而降低,在温度为10 ℃时,降到最低值5.1 U/mg。本研究表明,在冻融单一胁迫下,黑麦草幼苗体内POD活性随温度降低逐渐升高,冻融促使黑麦草幼苗的细胞膜透性降低,促进细胞自由基积累,从而激活了黑麦草细胞内抗氧化系统,使POD活性逐渐增强[14]。与复合胁迫下的黑麦草幼苗体内POD活性相比,pH值4.5酸沉降胁迫下的幼苗体内POD活性随温度下降逐渐升高,在温度降到 -2 ℃ 时,升到最高值 6.28 U/mg,在温度上升到10 ℃时,POD活性下降到最低值4.68 U/mg(图5)。这与王涛等关于模拟酸雨胁迫对菲白竹生理特性的影响研究结果[15]一致。黑麦草幼苗体内POD活性逐渐提高,说明黑麦草自身有较强的抗酸能力,对逆境的适应能力强,这与郭慧媛等关于模拟酸雨对毛竹叶片抗氧化酶活性的影响研究结果[16]一致。在冻融+酸沉降复合胁迫条件下,幼苗体内POD活性随温度降低而升高,在-2 ℃时,升到最高值,为6.96 U/mg;在温度升到10 ℃时,POD活性下降,为5.49 U/mg(图5)。以上分析表明,冻融+酸沉降复合胁迫能使黑麦草幼苗体内的POD活性含量增加。
3结论
变温胁迫可使黑麦草积累渗透调节物质,增加了其可溶性蛋白、可溶性糖、MDA含量,SOD和POD活性也明显提升。在酸沉降胁迫下,黑麦草幼苗体内可溶性蛋白、可溶性糖、MDA含量呈下降趋势,SOD和POD活性提高。结果表明,黑麦草自身具有较强的抗寒性,在低温胁迫下可以起到自我保护的作用,同时黑麦草也具有一定的抗酸性,对逆境的适应能力强。
参考文献:
[1]李国辉,李志坚,胡跃高. 青刈黑麦产草量与营养动态分析[J]. 草地学报,2000,8(1):49-54.
[2]多立安,赵树兰. 刈割冬牧70黑麦地上生物量动态与草群再生效应的研究[J]. 植物研究,2001,21(2):304-307.
[3]Dax M,Yin W X,Zhao Y X,et al. The production and scavenging of reactive oxygen specices in plants[J]. Chin J Biology,2001,17(2):121-125.
[4]杜子銀,蔡延江,王小丹,等. 土壤冻融作用对植物生理生态影响研究进展[J]. 中国生态农业学报,2014,22(1):1-9.
[5]黄婷,邵辉,吴坤胜,等. 模拟酸雨对黑麦草种子萌发及生理特性的影响[J]. 种子,2011,30(4):8-11.
[6]黄辉,黄朝法. 酸雨对植物生理生态特性的影响研究进展[J]. 林业勘察设计,2008(1):46-49.
[7]李轶冰,杨顺强,任广鑫,等. 低温处理下不同禾本科牧草的生理变化及其抗寒性比较[J]. 生态学报,2009,29(3):1341-1347.
[8]黄开志. 模拟酸雨对蔬菜细胞透性和营养及卫生品质的影响[J]. 生物学通报,2000,35(2):34.
[9]童贯和,梁惠玲. 模拟酸雨及其酸化土壤对小麦幼苗体内可溶性糖和含氮量的影响[J]. 应用生态学报,2005,16(8):1487-1492.
[10]孙亚琴,尚鹤,邓仕槐,等. 模拟酸雨胁迫对夹竹桃MDA和MP的影响[C]//四川省环境科学学会.四川省水污染控制工程学术交流会论文集. 成都,2009:176-181.
[11]宋采博,王波. 不同温度对白三叶种子发芽及幼苗生长的影响[J]. 湖北农业科学,2009,48(11):2772-2774.
[12]王强,金则新,彭礼琼. 模拟酸雨对乌药幼苗生理生态特性的影响[J]. 浙江大学学报(理学版),2013,40(4):447-455.
[13]吕家根,占达东,王周平,等. 酸雨胁迫下小麦微弱延迟发光及其生理生态变化相关性研究[J]. 化学学报,2003,61(5):760-764.
[14]王晨光,王希,苍晶,等. 低温胁迫对水稻幼苗抗冷性的影响[J]. 东北农业大学学报,2004,35(2):205-207.
[15]王涛,张珊珊,谢寅峰,等. 模拟酸雨胁迫对菲白竹生理特性的影响[J]. 东北林业大学学报,2014,42(11):17-21.
[16]郭慧媛,马元丹,王丹,等. 模拟酸雨对毛竹叶片抗氧化酶活性及释放绿叶挥发物的影响[J]. 植物生态学报,2014,38(8):896-903.
