哈尔滨市12种常见树木的滞尘能力比较
2017-06-30黄靖懿黄泽刘慧婧车代弟
黄靖懿++黄泽++刘慧婧++车代弟
摘要:选取糖槭、色木槭等12种哈尔滨市常见树木,对其单位面积滞尘量、单叶滞尘量和叶片质膜透性、可溶性糖含量、丙二醛(MDA)含量、脯氨酸(Pro)含量进行测定。结果表明,不同树木间的滞尘量存在较大差异,选择不同滞尘量计量单位,树木滞尘量排序会相应发生变化;不同樹木在自然界中蒙尘后,叶片质膜透性、可溶性糖含量、丙二醛(MDA)含量、脯氨酸(Pro)含量均出现不同程度的上升,上升幅度因树木种类不同而存在较大差异,其中叶片质膜透性增长率与丙二醛(MDA)含量增长率之间存在显著的线性相关关系,通过隶属函数法综合4个生理指标,得到抗性排序为毛樱桃>榆叶梅>水曲柳>色木槭>暴马丁香>花楸>丁香>山荆子>糖槭>水蜡>榆树>核桃楸;毛樱桃、花楸、丁香单位叶面积滞尘量、单叶滞尘量相对较大,抗性较强,可作为哈尔滨市抗尘的优良树木。
关键词:树木;滞尘量;生理指标;滞尘能力;哈尔滨;毛樱桃;质膜透性
中图分类号: X173文献标志码: A文章编号:1002-1302(2017)08-0117-04
近年来,随着我国城市交通和城市化建设的高速发展,扬尘、土壤尘、机动车尾气等俨然成为21世纪城市的主要污染来源,产生一系列大气污染问题。颗粒物是当今城市大气污染的主要污染物[1],由于影响大气能见度、产生光化学烟雾、加剧温室效应,且含有重金属、细菌、病毒、PAHs等致癌物质,已引起政府、公众、学者前所未有的关注。
园林树木作为城市绿地的主体,对一定范围内的粉尘有明显过滤、吸附作用[2],而园林树木叶片滞留大气颗粒物的能力成为城市绿化树种选择的一个重要指标。近年来,国内外许多学者开展了关于城市树木滞尘能力的研究[3-7],但大部分研究仅从滞尘量大小来单一评价树木的滞尘能力,认为滞尘能力与滞尘量相关。也有很多学者建议,在研究树木滞尘量的同时,应关注树木本身的抗逆性,只有树木本身滞尘量大且受尘污染影响小,才是真正滞尘能力强的树木,才适合作为城市滞尘的树木[8-9],而目前蒙尘对树木抗性影响的研究鲜见报道。本试验选取黑龙江省哈尔滨市糖槭、色木槭等12种常用树木为研究对象,对其滞尘量和抗性进行分析,为哈尔滨市滞尘树木的选择提供参考。
1材料与方法
1.1试验树木的选择
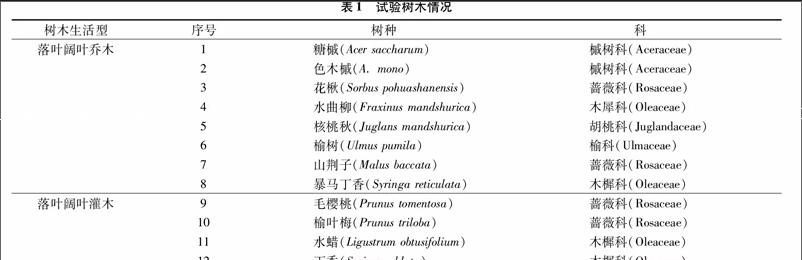
哈尔滨市自然植被属温带针阔混交林,依据城区常用树木的资料统计[10],本试验选取哈尔滨市12种常用树木进行滞尘能力和抗性指标测定,其中落叶阔叶乔木8种、落叶阔叶灌木4种(表1)。
1.2样品采集及处理
样品采集地点设在树木种类丰富、交通相对繁忙的哈尔滨市珠江路,分别于雨后若干天进行采样,具体采集日期为2016年6月11日(尘积累7 d)、6月23日(尘积累9 d)、7月9日(尘累计9 d)、7月22日(尘累计11 d)。选择树龄及生长状况基本相同的12种树木,定株、定时(09:00)采集树冠中下部向阳面光照强度一致的同龄叶片,将采集好的20~40张叶片装入已编好号码的密封袋中,置于4 ℃冰箱中带回实验室,运输途中应尽量避免振荡。为减少试验误差,对照组和试验组样品的采集应遵循同株、同向原则,试验期间每天对对照组样品进行冲洗,并挂牌标记。测量时,假设试验组初始叶片滞尘量为0。
1.3测定内容和方法
1.3.1滞尘量叶片的滞尘量采用质量差法进行测定[11]。将采集的叶片置于盛有去离子水的烧杯中浸泡2 h;用毛刷刷下叶片表面的颗粒物,并用小毛刷轻轻刷洗装叶片的保鲜袋;将叶片取出,清洗液用烘干已称质量(m1)的滤纸过滤;将滤纸放入烘干箱内80 ℃烘干24 h,用万分之一天平称质量(m2),m1与m2的差值即为采集叶片所附着的降尘颗粒物质量。清洗过的叶片用滤纸吸干,用LAI-2000叶面积仪测量面积S[12],计算阔叶树木单位叶面积滞尘量、单叶滞尘量,公式为:
单位叶面积滞尘量(g/m2)=(m2-m1)/N0S;单叶滞尘量(g/叶)=(m2-m1)S0/N0S。
式中:S为供试叶样的平均叶面积;S0为供试树木平均叶面积;N0为供试叶样数。
1.3.2生理指标将7月9日、7月22日采集的叶样用去离子水清洗干净,分别采用电导仪法、蒽酮比色法、茚三酮比色法、硫代巴比妥酸法测定叶片质膜透性、可溶性糖(SS)含量、游离脯氨酸(Pro)含量、丙二醛(MDA)含量这4个生理指标。由于2次采集的叶片尘累计时间相差较小,所以4个生理指标的数值为2次测得数据的平均值。每个供试叶片测定重复3次。
1.4数据处理
采用隶属函数法对树木在自然界蒙尘后尘抗性数据进行统计[13]。如果某一生理指标与尘抗性呈正相关关系,计算公式为:Xμ=(X-Xmin)/(Xmax-Xmin);如果某一指标与尘抗性呈负相关关系,则计算公式为:Xμ=1-(X-Xmin)/(Xmax-Xmin)。式中,X表示某一指标的测定值;Xmax表示某一指标测定值中的最大值;Xmin表示某一指标测定值中的最小值。本试验中质膜透性、MDA含量采用负相关计算公式,其他指标均采用正相关计算公式。试验数据采用Excel、SPSS19.0软件进行统计分析。
2结果与分析
2.1单位叶面积滞尘量
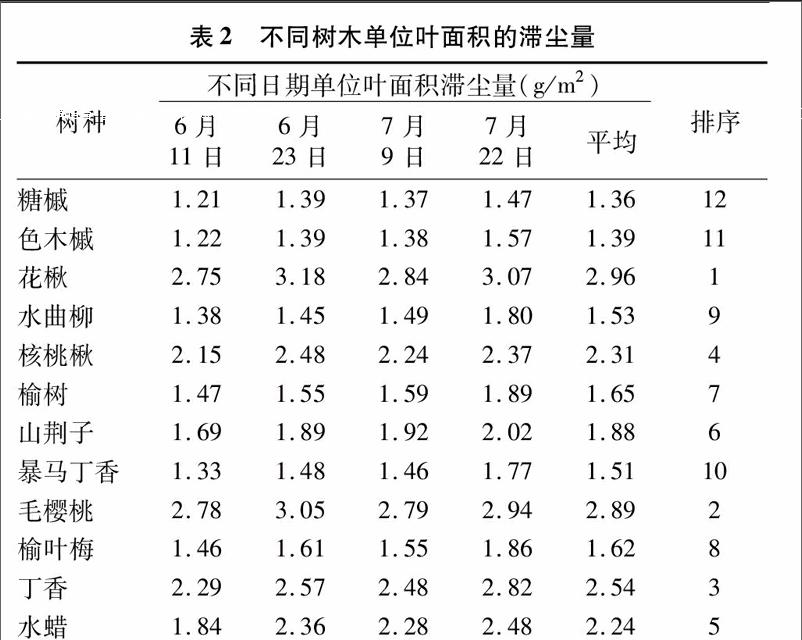
树木单位叶面积滞尘量是指在一定时间段里叶片单位面积粉尘的滞留量,其大小一定程度上反映树木的滞尘能力[14]。由表2可见,哈尔滨市12种树木的平均单位叶面积滞尘量为1.36~2.96 g/m2,经方差分析,不同树木间单位叶面积滞尘量存在差异;花楸单位叶面积滞尘量相对最大,为2.96 g/m2,毛樱桃次之,为2.89 g/m2,丁香第三,为 2.54 g/m2,糖槭单位叶面积滞尘量相对最小,仅为 1.36 g/m2,与最大的花楸相差约2倍,这与柴一新等研究结论[15]类似;落叶阔叶乔木与灌木的单位叶面积滞尘量整体排序分布较为均匀,并未因树木类型不同出现明显的排序分化现象,这与范舒欣等研究结论[16]类似。
2.2单叶滞尘量
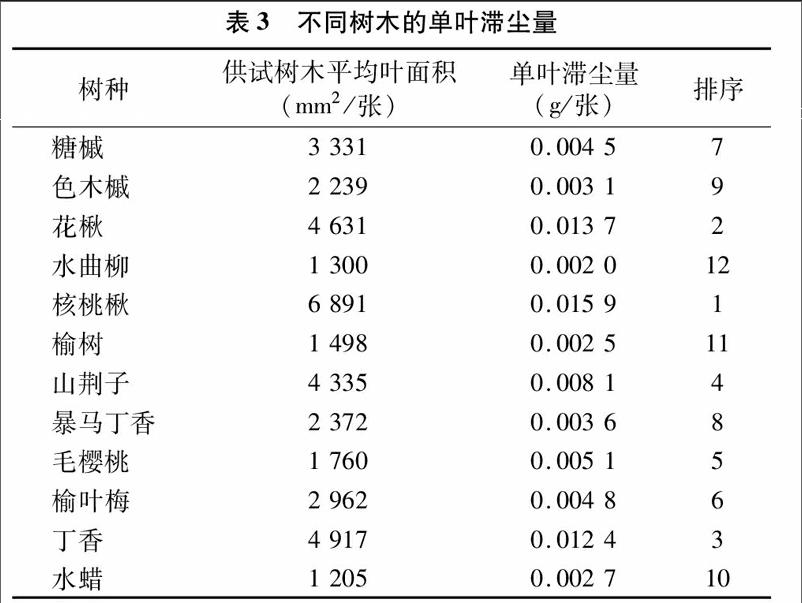
单叶滞尘量为一个独立叶片在一定时间内滞留的粉尘量[17]。由表3可见,哈尔滨市12种树木的单叶滞尘量为 0.002 0~0.015 9 g/张,不同树木间单叶滞尘量差异较大,其中核桃楸单叶滞尘量相对最大,为0.015 9 g/张,花楸次之,為0.013 7 g/张,丁香第三,为0.012 4 g/张,水曲柳的单叶滞尘量相对最小,仅为0.002 0 g/张,与核桃楸相差约7倍;落叶阔叶乔木与灌木的单叶滞尘量排序分布仍较为均匀,因植物类型不同可能带来的排序分化现象不明显。
2.3蒙尘对树木生理指标的影响
2.3.1蒙尘对叶片质膜透性的影响质膜是选择透过性膜,能调节和控制细胞内外物质的运输和交换,在逆境生理学中,细胞质膜透性是反映植物抗逆性程度的良好指标[18]。由图1可见,12种树木在自然界中蒙尘后,质膜透性均呈一定的增强趋势,不同树木的增长率有所差异,其增长率范围在 23%~108%之间;榆树的增长率相对最大,为108%,核桃楸次之,为87%,糖槭的增长率相对最小,为23%,方差分析(表4)显示,试验组与对照组差异极显著(P<0.01)。树木叶片蒙尘之后电导率增大,表明尘对叶片的细胞质膜通透性产生影响。
2.3.2蒙尘对叶片可溶性糖含量的影响可溶性糖含量在植物体内对环境胁迫的变化非常敏感,既是指示植物遭受环
境胁迫程度的重要指标,又是植物细胞内重要的渗透调节物质与适应环境的信号物质。由图2可知,树木蒙尘可使12种树木叶片的可溶性糖适量积累,与对照组相比,可溶性糖含量有小幅度增加,其增长率范围在10%~65%之间,这可能是叶片蒙尘后,其气孔受到不同程度的堵塞而使叶内O2浓度下降,CO2浓度相对升高,呼吸过程受到抑制[19],可溶性糖短暂积累;毛樱桃可溶性糖含量的增长率相对最大,为65%。
2.3.3蒙尘对叶片丙二醛(MDA)含量的影响丙二醛是植物对逆境反应的重要生理指标,也是植物细胞膜脂过氧化产物之一,其含量多少可反映植物适应环境或者受不良环境胁迫的情况,含量越高,受伤害的程度越大。由图3可知,蒙尘后树木叶片丙二醛含量均有不同程度的增加,其增长率范围在26%~98%之间;不同树木叶片的丙二醛含量存在明显差异,其中榆树的增长率相对最大,为98%,糖槭的增长率相对最小,为26%,说明糖槭的抗尘性强于榆树。由图4可知,树木叶片蒙尘后丙二醛含量的增长率与质膜透性增长率呈正相关关系(决定系数r2≈0.88),这与钮福祥等的结论[20-21]较为一致,说明胁迫环境下过氧化作用加强,可积累较多的丙二醛,造成膜透性增强、离子外渗。
2.3.4蒙尘对叶片脯氨酸(Pro)含量的影响脯氨酸是植物体内重要的渗透调节物质,胁迫条件下植物体内游离脯氨酸
含量与植物抗性之间存在相关性,它的积累可保护植物抵抗逆境的胁迫。有研究表明,当植物遇到干旱、冰冻、高温、低温等环境时,都会在体内出现脯氨酸的累积[22]。由图5可知,蒙尘后树木叶片的脯氨酸含量均有所增长,其增长率范围在12%~68%之间;不同树木叶片的脯氨酸含量增长存在差异,丁香的脯氨酸含量增长率相对最大,为68%,核桃楸次之,为53%,水曲柳第三,为29%,毛樱桃的增长率相对最小,仅为12%。
2.4蒙尘后树木抗性的综合评定
植物受到逆境时,其生理变化是错综复杂的,并受到多种因素的影响,孤立地用一个指标评价植物抗性能力很难真实反映植物的抗性本质。因此,为综合评价各种树木的抗尘能力,采用隶属函数值法,以各项指标隶属度的平均值作为树木抗性能力综合鉴定标准进行比较。由于质膜透性、丙二醛含量表征植物受损程度,与植物抗性呈负相关,分析时采用反隶属函数值;脯氨酸含量、可溶性糖含量表征植物对逆境的适应能力,分析时采用隶属函数值[23]。由表5可见,树木抗性大小依次为毛樱桃>榆叶梅>水曲柳>色木槭>暴马丁香>花楸>丁香>山荆子>糖槭>水蜡>榆树>核桃楸;落叶阔叶乔木与灌木的抗性整体排序分布较为均匀,以丁香为界,抗性排名前6位中乔木占66%,灌木占33%。
3结论与讨论
树木滞尘是一种极为有效的生物滞尘方式,不同树木的单位叶面积滞尘量主要取决于树木叶片结构特征、叶表面粗糙程度、被毛情况及滞尘方式等[24];而单叶滞尘量是基于单位叶面积滞尘量与树木平均单叶面积共同计算得到的,侧重于反映叶片形状与大小对树木滞尘的影响,所以树木单位叶面积滞尘量与单叶滞尘量的排序情况会有所不同,不应以某一个单一的计量结果来评价树木的滞尘量。本研究发现,不同树木单位叶面积滞尘量与单叶滞尘量之间差异显著;丁香上表皮虽无柔毛,但叶表气孔周围具脊状突起,其滞尘方式以附着为主,加之叶片面积较大且尖端易蜷曲向上,所以其单位叶面积滞尘量和单叶滞尘量均较大;而水蜡单位叶面积滞尘量为 2.24 g/m2,排序在中等偏上,滞尘方式以滞留为主,但由于其叶片面积较小,致使单叶滞尘量偏小。不过,有研究表明,水蜡的单位叶面积滞尘量较小[25],与本研究结论相反,这说明以滞留为主的滞尘方式更易受外界环境的影响,其滞尘量不仅与自身形态有关,也与其生长的外界环境污染程度有关。
质膜透性、丙二醛含量、脯氨酸含量、可溶性糖含量是植物对逆境反应的4个重要生理指标。丙二醛是植物细胞膜脂过氧化产物之一,能够与细胞内各种成分发生反应,从而引起酶和膜的严重损伤[26];可溶性糖和脯氨酸含量在细胞质中的增加,可以提高植物细胞的渗透调节能力,降低质膜受伤害的程度,对植物抵抗逆境具有重要作用[27]。试验结果表明,蒙尘后12种树木叶片的膜脂过氧化产物MDA含量和细胞膜透性都逐渐增加;抗性强的毛樱桃蒙尘后,细胞膜透性、MDA含量增长率相对较小,而抗性弱的榆树细胞膜透性、MDA含量增长率相对较大,这2项指标与12种树木的抗性排序基本一致,说明质膜透性、MDA含量可作为树木抗尘的重要鉴定指标。脯氨酸、可溶性糖都是植物细胞重要的渗透调节物质,在逆境环境下适当积累,可使自身适应不良环境而产生一定的忍耐力[28]。不同的树木对蒙尘的反应不同,在细胞内积累的渗透调节物质也不同,毛樱桃可溶性糖含量的增长率相对最大,脯氨酸含量的增长率相对较小,而丁香可溶性糖含量的增长率较小,脯氨酸含量的增长率相对较大,这说明蒙尘后毛樱桃细胞内积累的主要渗透调节物质是可溶性糖,而丁香是脯氨酸。
在对树木滞尘量研究的同时,也应关注树木本身的抗逆性,只有本身的滞尘量大且受尘污染影响小的树木,才是真正滞尘能力强的树木,才适合作城市滞尘的树木。毛樱桃、花楸、丁香等单位叶面积滞尘量和单叶滞尘量相对较大,抗性也较强,可作为哈尔滨市抗尘的优良绿化树木;榆树单位面积滞尘量和单叶滞尘量相对较小,抗性较差,在尘污染较严重的地区不适宜种植该树种;核桃楸单位面积滞尘量和单叶滞尘量相对较大,但其抗性较弱,可以作为大气尘污染的监测植物。需指出的是,本试验大多数树木都能够感知蒙尘,并调节自身生理指标变化来响应蒙尘,但不同树木存在着各个生理指标变化的不协同性和复杂性,有待进一步探讨。另外,在树木叶片单位面积滞尘量、单叶滞尘量抗性排序中,不同树木类型间未出现明显的排序分化现象,乔、灌木的排序分布较为均匀,不同生活型的树木可在自然界中截留不同高度、不同来源、不同类型的粉尘颗粒,这就须要合理配置不同类型的乔、灌、草植物,形成不同群落配置模式,在增加景观效果的同时,又可有效提升绿地滞尘效应,缓解空气污染,改善城市的环境质量。
参考文献:
[1]乔玉霜,王静,王建英. 城市大气可吸入颗粒物的研究进展[J]. 中国环境监测,2011,27(2):22-26.
[2]闫晓云,张秋良,韩鹏,等. 呼和浩特市绿化树种综合评价及树种选择[J]. 干旱区資源与环境,2011,25(3):135-140.
[3]于志会,赵红艳,杨波. 吉林市常见园林植物滞尘能力研究[J]. 江苏农业科学,2012,40(6):173-175.
[4]杨瑞卿,肖扬. 徐州市主要园林植物滞尘能力的初步研究[J]. 安徽农业科学,2008,36(20):8576-8578.
[5]王蓉丽,方英姿,马玲. 金华市主要城市园林植物综合滞尘能力的研究[J]. 浙江农业科学,2009(3):574-577.
[6]康博文,刘建军,王得祥,等. 陕西20种主要绿化树种滞尘能力的研究[J]. 陕西林业科技,2003(4):54-56.
[7]Beckett K P,Freer S P,Taylor G. The capture of particulate pollution by trees at five contrasting urban sites[J]. Arboricultural Journal,2000,24(2):209-230.
[8]戚继忠,魏进华,张倩,等. 城市树木滞尘能力研究及存在的问题与对策[J]. 世界林业研究,2013,26(3):52-57.
[9]余海龙,黄菊莹. 城市绿地滞尘机理及其效应研究进展[J]. 西北林学院学报,2012,27(6):238-241,247.
[10]陈玮,何兴元,张粤,等. 东北地区城市针叶树冬季滞尘效应研究[J]. 应用生态学报,2003,14(12):2113-2116.
[11]刘霞,李海梅. 园林植物滞尘效应的研究[J]. 北方园艺,2007(8):73-76.
[12]高金晖,王冬梅,赵亮. 植物叶片滞尘规律研究——以北京市为例[J]. 北京林业大学学报,2007,29(2):94-99.
[13]刘丹,陈保军,杨永贵,等. 4种幼树对二氧化硫胁迫的抗性生理响应[J]. 西北植物学报,2013,33(12):2441-2450.
[14]Wang F. Study on dust-retention ability of major afforestation tree species in Tianjin airport economic area[J]. Journal of Arid Land Resources and Environment,2015,29(1):100-104.
[15]柴一新,祝宁,韩焕金. 城市绿化树种的滞尘效应——以哈尔滨市为例[J]. 应用生态学报,2002,13(9):1121-1126.
[16]范舒欣,晏海,齐石茗月,等. 北京市26种落叶阔叶绿化树种的滞尘能力[J]. 植物生态学报,2015,39(7):736-745.
[17]周志翔,邵天一,王鹏程,等. 武钢厂区绿地景观类型空间结构及滞尘效应[J]. 生态学报,2002,22(12):2036-2040.
[18]夏宗良,刘全军,武轲,等. 二氧化硫胁迫对玉米幼苗叶片膜脂过氧化和抗氧化酶的影响[J]. 玉米科学,2009,17(4):51-54.
[19]江胜利,金荷仙,许小连. 杭州市常见道路绿化植物滞尘能力研究[J]. 浙江林业科技,2011,31(6):45-49.
[20]钮福祥,郭小丁. 甘薯品种抗旱性生理指标及其综合评价初探[J]. 作物学报,1996,22(4):392-398.
[21]张木清,陈如凯,余松烈. 水分胁迫下蔗叶活性氧代谢的数学分析[J]. 作物学报,1996,22(6):729-735.
[22]夏晗,黄金生. 低温、干旱和盐胁迫下沙冬青幼苗脯氨酸质量分数的变化[J]. 吉林林业科技,2007,36(4):12-20.
[23]李彦慧,李向应,白瑞琴,等. 4种李属彩叶树木对SO2的抗性[J]. 林业科学,2008,44(2):28-33.
[24]王会霞,石辉,李秧秧. 城市绿化植物叶片表面特征对滞尘能力的影响[J]. 应用生态学报,2010,21(12):3077-3082.
[25]赵勇,李树人,阎志平,等. 城市绿地的滞尘效应及评价方法[J]. 华中农业大学学报,2002,21(6):582-586.
[26]张倩,何婧,王桢,等. 中华芦荟在不同胁迫下的丙二醛和可溶性糖含量的变化[J]. 西南民族大学学报(自然科学版),2009,35(2):290-292.
[27]许祥明,叶和春,李国凤. 脯氨酸代谢与植物抗渗透胁迫的研究进展[J]. 植物学通报,2000,17(6):536-542.
[28]李彦奇,姚正培. 低温胁迫对三种早春短命植物生理生化的影响[J]. 新疆农业科学,2012,40(9):1608-1615.
