不同土壤环境下落叶松人工林昆虫群落结构及动态的差异*
2017-06-23王利东张子龙
顾 伟 马 玲 孙 虎 王利东 张子龙
(1.东北林业大学林学院 哈尔滨 150040; 2.东北林业大学凉水自然保护区 伊春153106)
不同土壤环境下落叶松人工林昆虫群落结构及动态的差异*
顾 伟1,2马 玲1孙 虎2王利东2张子龙1
(1.东北林业大学林学院 哈尔滨 150040; 2.东北林业大学凉水自然保护区 伊春153106)
【目的】 研究不同土壤条件下落叶松人工林昆虫群落结构及动态的差异,为森林经营管理、生物多样性保护提供理论基础和科学依据。【方法】 以小兴安岭凉水自然保护区内封山育林状态下落叶松人工林为研究对象,根据地形土壤条件,选择埋藏潜育暗棕壤-落叶松人工林(A)、厚层埋藏暗棕壤-落叶松人工林(B)、暗棕壤性土-落叶松人工林(C)、典型暗棕壤-落叶松人工林(D)4种生境类型,分别设置3个20 m×20 m的样地,采用扫网法、巴氏罐诱法、震落法、高枝剪标准枝法对样地内昆虫群落进行系统的调查。【结果】 共捕获昆虫样本5 204头,经鉴定分属10目77科250种,各生境均以双翅目和膜翅目为优势类群,但所占比例具有较大差异。各生境昆虫群落科级优势类群季节动态也表现出较大的差异。聚类分析表明4种生境昆虫群落归为2类,A、B生境归为一类,C、D生境归为一类。不同生境间昆虫群落物种多样性指数和均匀度指数大小均表现为C生境>A生境>B生境>D生境,丰富度指数大小表现为样地B生境>D生境>A生境>C生境,优势集中性指数大小表现为D生境>B生境>A生境>C生境。时间序列上,B生境和D生境昆虫群落物种多样性指数与均匀度指数及丰富度指数均呈显著的正相关,但前者相关系数均显著大于后者。各生境昆虫群落中不同营养类群物种数均表现为植食性类群>中性类群>捕食性类群>寄生性类群,但各生境中不同营养类群物种数所占比例具有明显差异。各生境昆虫群落相对稳定性为B生境>C生境>D生境>A生境。主分量分析表明对昆虫群落变化起着主导作用的因子在各生境中具有较大差异。【结论】 不同土壤条件下落叶松人工林各昆虫群落结构动态具有较大差异,昆虫群落物种多样性指数与丰富度指数正相关系数越大,群落稳定性越强,同时中性昆虫对昆虫群落相对稳定性起到显著的调控作用。4种生境中厚层埋藏暗棕壤落叶松人工林综合环境条件最适宜昆虫群落发生发展。
昆虫群落; 稳定性分析; 主分量分析; 落叶松人工林; 凉水自然保护区
兴安落叶松(Larixgmelinii)是东北林区主要用材树种之一,近几十年来,随着天然林面积不断减少,大面积落叶松人工林成为东北林区重要的后备森林资源。昆虫种类繁多,习性各异,蕴藏着极大的生物资源量,对维持森林生态平衡起到重要作用。昆虫群落能占据大量尺度小、异质性强的各类生境,且对生境的变化极为敏感,具有广谱的生物地理学和生态学探针功能(Samways, 1993)。已有一些学者对落叶松人工林昆虫群落开展研究,李孟楼等(1995)认为秦岭火地塘林区华北落叶松(L.principis-rupprechtii)人工林昆虫群落结构随季节、海拔发生变化,并阐述了优势种的种群演替规律。赵东芳等(2011)研究了吉林敦化市不同年龄长白落叶松(L.olgensis)人工林昆虫多样性的变化规律,认为昆虫群落结构随林龄的增大经历着剧烈的演变。刘哲强等(2015)认为小兴安岭凉水自然保护区兴安落叶松人工林和和原始阔叶红松(Pinuskoraiensis)林昆虫群落结构相对于其他类型人工林更为稳定,捕食性昆虫和中性昆虫对昆虫群落稳定性发挥着关键性作用。土壤是森林生态系统的重要组成部分,土壤环境与昆虫的关系十分密切,很多昆虫生长发育的部分阶段或时期需要在土壤中完成,然而关于落叶松人工林昆虫群落随土壤环境不同而产生结构及动态差异的研究未见报道。
本研究以小兴安岭南坡凉水自然保护区内封山育林状态下恢复约60年的兴安落叶松人工林为研究对象,对4种不同土壤条件下林内昆虫群落进行了调查,研究分析昆虫群落结构、多样性及稳定性特征,探讨落叶松人工林昆虫群落随不同土壤环境的生态响应,为小兴安岭森林恢复期的森林经营管理、生物多样性保护提供理论基础和科学依据。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于黑龙江省伊春市带岭区凉水自然保护区(128°47′8″—128°57′19″E,47°6′49″—47°16′10″N),区内地带性植被是以红松为主的温带针阔混交林,具有明显的温带大陆性季风气候特征,冬季寒冷干燥、夏季温热多雨,年平均降水680 mm左右,年平均气温-0.3 ℃,年平均最高气温7.5 ℃,年平均最低气温-6.6 ℃(马建章等, 1993)。
研究地为阔叶红松林采伐后人工种植的兴安落叶松林,且在封山育林状态下已恢复约60年,面积19 hm2,植被覆盖率100%,郁闭度0.8。根据地形土壤条件,在研究地中选择4种不同生境样地,依次为生境(A): 埋藏潜育暗棕壤-落叶松人工林,阳坡相对下部,地势较平缓,坡度约10°,海拔387 m,土壤A1层约20 cm,团粒结构,腐殖质含量高,AB层约20 cm,深浅颜色交错,块状至片状结构,B层灰色有沙粒,有黏土矿物淀积; 生境(B): 厚层埋藏暗棕壤-落叶松人工林,阴坡相对中部,坡度20°,海拔400 m,土壤A1层约50 cm,其中A11层约25 cm,腐殖质含量高,团粒结构,A12层约20 cm,粒状结构,A13层10~20 cm,具有不明显的核粒状结构,B层约20~30 cm,较明显核状结构,有一定腐殖质,有沙粒; 生境(C): 暗棕壤性土-落叶松人工林,山顶,海拔418 m,土壤A1层约15 cm,腐殖质含量高,团粒结构,有朽木,大量黑炭,多根系,无B层,AC层约15 cm,粒状结构,较黏有沙粒; 生境(D): 典型暗棕壤-落叶松人工林,阳坡相对中上部,坡度30°,海拔406 m,土壤A1层约15 cm,团粒结构,有菌丝,AB层约30 cm,有不明显的混合堆积特征,质地较黏,多根系,有炭屑,B层约25 cm,有不明显的核状结构,有黏粒淀积,下部砂砾较多,有根系,BC层约10 cm,以风化沙为主,有铁淀积。
1.2 研究方法
1.2.1 调查方法 每种生境沿等高线方向取20 m×20 m样地3块。在2013年和2014年5月至9月中旬,每15天调查1次,遇雨天顺延。对草本层和灌木层昆虫采用扫网法进行调查,捕虫网直径38 cm,深度为70 cm,柄长1.3 m,扫网路径为在每个样地中以“Z”字形来回取100网,扫网时一个往返为1次,每次往返呈180°(顾伟等, 2011); 对栖息树干昆虫采用震落法和高枝剪标准枝法进行采集,对样地内全部乔木每株分别从4个方向套袋剪取2个枝条,地面铺设白布,震动树木收集掉落的昆虫(姬兰柱等, 2004)。对地表昆虫采用巴氏罐诱法(Baars, 1979)采集,用一次性塑料水杯(高9.0 cm,口径7.5 cm)作为巴氏罐诱法容器,每块样地内设诱杯50 个,引诱剂为醋、糖、医用酒精和水的混合物,质量比为2∶1∶1∶20,每个诱杯内放引诱剂40~60 mL,诱杯放置时间5天(顾伟等, 2014)。将采集到的鳞翅目等易损昆虫放入三角纸袋内,其他昆虫放入毒瓶中毒死后带回实验室分类鉴定并计数。
1.2.2 营养类群的划分 从昆虫的食性出发,以分类学上的科为单位,将昆虫群落大体分为4个功能群: 植食性昆虫、捕食性昆虫、寄生性昆虫、中性昆虫(包括腐生性昆虫和对植物几乎没有危害也不捕食其他动物的昆虫)(马玲等, 2012)。
1.3 数据处理
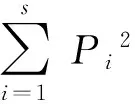
以每次调查数据为样本,以各类昆虫(按食性划分) 的种类数量和个体数量为变量,进行主分量分析,计算各因子的特征向量和负荷量值(余世孝, 1995)。以各生境中昆虫类群种类的发生情况为数据,进行欧氏距离的聚类分析(贾玉珍等, 2009; 杜秀娟等, 2009)。
群落相对稳定性测定采用群落物种数与个体数之比(St/Si) 及天敌类群种数与植食类群种数之比(Sn/Sp) 来表示。St/Si主要反映种类间数量上的制约作用,Sn/Sp则反映内部食物网络关系的复杂程度和相互制约程度(高宝嘉等, 1992)。
2 结果与分析
2.1 昆虫群落组成及季节动态
调查共捕获昆虫标本5 204号,分属10目77科250种,其中双翅目科数(25科)和种数(84种)均最多,其次是鞘翅目(18科56种)、半翅目(16科50种)、膜翅目(6科31种)、鳞翅目(6科20种)、脉翅目(2科3种)和革翅目(1科3种),长翅目、毛翅目和蛇蛉目最少,均为1科1种。个体数量上,双翅目和膜翅目为优势类群,个体数量分别占群落总个体数的51.77%和31.4%,半翅目和鞘翅目为群落常见类群,个体数量占群落比例共计15.37%,其他各目昆虫个体数量占群落比例合计不足2%,为群落稀有类群。
各生境中,昆虫群落组成具有一定差异,目级类群中,优势类群均为双翅目和膜翅目,但双翅目个体数所占比例在D生境中最高(62.91%),在A生境中最低(43.21%),膜翅目个体数量所占比例在B生境中最高(37.01%),在D生境中最低(23.21%)。科级类群中,A生境昆虫群落以膜翅目的蚁科(Formicidae)(24.51%)和姬蜂科(Ichneumonidae)(10.76%)为优势类群,菌蚊科(Mycetophilidae)、果蝇科(Drosophilidae)和厕蝇科(Fanniidae)为常见类群; B生境昆虫群落以膜翅目的蚁科(25.58%)和姬蜂科(11.1%)、双翅目的厕蝇科(14.49%)和菌蚊科(10.3%)为优势类群,果蝇科为常见类群; C生境和D生境昆虫群落均以菌蚊科(21.3%和27.14%)、姬蜂科(17.92%和14.73%)和厕蝇科(10.03%和10.41%)为优势类群,蚁科和毫蚊科(Trichoceridae)为常见类群。
各生境昆虫群落优势类群季节动态表现出较大的差异,A生境优势类群在5月末为蚁科、厕蝇科和叶甲科(Chrysomelidae),6月中旬为蚁科、姬蜂科和实蝇科(Tephritidae),6月末为蚁科和厕蝇科,7月中旬为蚁科、果蝇科和长足虻科(Dolichopodidae),7月末为菌蚊科、果蝇科、蚁科、眼蕈蚊科(Sciaridae)和厕蝇科,8月中旬为果蝇科、眼蕈蚊科、蚁科、蚊科(Culicidae)和姬蜂科,8月末为叶蝉科(Cicadellidae)、木虱科(Psyllidae)和姬蜂科,9月中旬为姬蜂科、木虱科和菌蚊科。
B生境优势类群在5月末为厕蝇科、蚁科和菌蚊科,6月中旬为蚁科、姬蜂科和毫蚊科,6月末为蚁科、厕蝇科和菌蚊科,7月中旬为蚁科、姬蜂科和果蝇科,7月末为蚁科、姬蜂科、果蝇科和菌蚊科, 8月中旬为蚁科、眼蕈蚊科和姬蜂科,8月末为菌蚊科、叶蝉科、蚁科和姬蜂科,9月中旬为姬蜂科、木虱科、蚁科和果蝇科。
C生境优势类群在5月末为姬蜂科、菌蚊科和厕蝇科,6月中旬为菌蚊科、蚁科、姬蜂科和食蚜蝇科(Syrphidae),6月末为菌蚊科、姬蜂科和厕蝇科,7月中旬为距甲科(Megalopodidae)、姬蜂科和蚁科,7月末为菌蚊科、姬蜂科、蚁科和厕蝇科,8月中旬为姬蜂科、蚁科和蚊科,8月末为木虱科、毫蚊科、菌蚊科、姬蜂科和叶甲科(Chrysomelidae),9月中旬为毫蚊科、姬蜂科、厕蝇科、菌蚊科和摇蚊科(Chironomidae)。
D生境优势类群在5月末为菌蚊科和厕蝇科,6月中旬为姬蜂科、菌蚊科和蚁科,6月末为菌蚊科、厕蝇科、蚁科和姬蜂科,7月中旬为距甲科(Megalopodidae)、菌蚊科和实蝇科(Tephritidae),7月末为菌蚊科、姬蜂科和蚁科,8月中旬为蚊科和姬蜂科,8月末为菌蚊科、毫蚊科、姬蜂科和叶蝉科,9月中旬为胡蜂科(Vespidae)、菌蚊科和厕蝇科。

表1 不同生境昆虫群落组成Tab.1 Insect communities in different types habitat
续表Continued
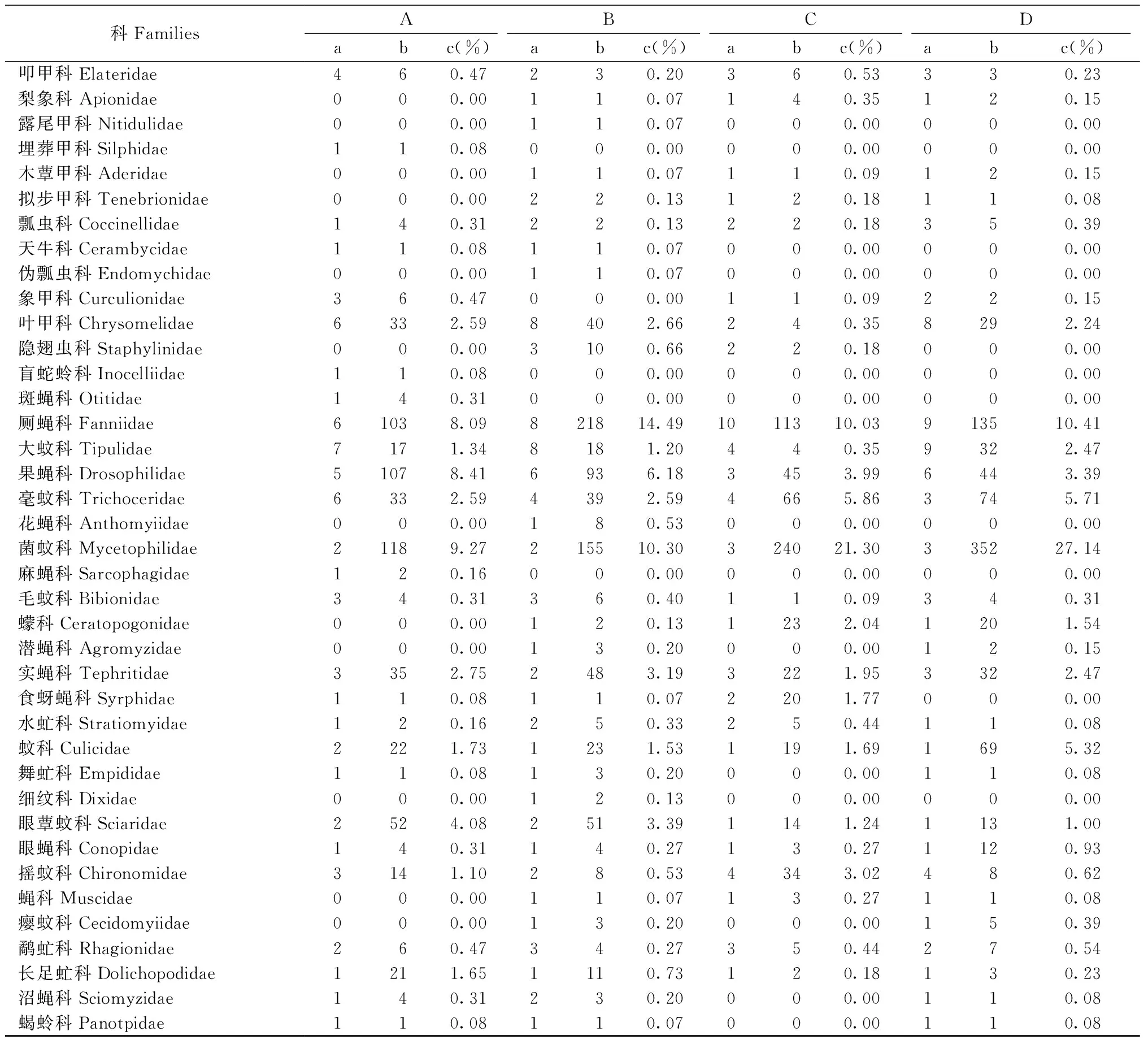
科FamiliesABCDabc(%)abc(%)abc(%)abc(%)叩甲科Elateridae460.47230.20360.53330.23梨象科Apionidae000.00110.07140.35120.15露尾甲科Nitidulidae000.00110.07000.00000.00埋葬甲科Silphidae110.08000.00000.00000.00木蕈甲科Aderidae000.00110.07110.09120.15拟步甲科Tenebrionidae000.00220.13120.18110.08瓢虫科Coccinellidae140.31220.13220.18350.39天牛科Cerambycidae110.08110.07000.00000.00伪瓢虫科Endomychidae000.00110.07000.00000.00象甲科Curculionidae360.47000.00110.09220.15叶甲科Chrysomelidae6332.598402.66240.358292.24隐翅虫科Staphylinidae000.003100.66220.18000.00盲蛇蛉科Inocelliidae110.08000.00000.00000.00斑蝇科Otitidae140.31000.00000.00000.00厕蝇科Fanniidae61038.09821814.491011310.03913510.41大蚊科Tipulidae7171.348181.20440.359322.47果蝇科Drosophilidae51078.416936.183453.996443.39毫蚊科Trichoceridae6332.594392.594665.863745.71花蝇科Anthomyiidae000.00180.53000.00000.00菌蚊科Mycetophilidae21189.27215510.30324021.30335227.14麻蝇科Sarcophagidae120.16000.00000.00000.00毛蚊科Bibionidae340.31360.40110.09340.31蠓科Ceratopogonidae000.00120.131232.041201.54潜蝇科Agromyzidae000.00130.20000.00120.15实蝇科Tephritidae3352.752483.193221.953322.47食蚜蝇科Syrphidae110.08110.072201.77000.00水虻科Stratiomyidae120.16250.33250.44110.08蚊科Culicidae2221.731231.531191.691695.32舞虻科Empididae110.08130.20000.00110.08细纹科Dixidae000.00120.13000.00000.00眼蕈蚊科Sciaridae2524.082513.391141.241131.00眼蝇科Conopidae140.31140.27130.271120.93摇蚊科Chironomidae3141.10280.534343.02480.62蝇科Muscidae000.00110.07130.27110.08瘿蚊科Cecidomyiidae000.00130.20000.00150.39鹬虻科Rhagionidae260.47340.27350.44270.54长足虻科Dolichopodidae1211.651110.73120.18130.23沼蝇科Sciomyzidae140.31230.20000.00110.08蝎蛉科Panotpidae110.08110.07000.00110.08
①A: 埋藏潜育暗棕壤样地 Buried-gley dark brown soil sample; B: 厚层埋藏暗棕壤样地 Thick-buried dark brown soil sample; C: 暗棕壤性土样地Dark brown loam soil sample; D: 典型暗棕壤样地 Typical dark brown soil sample; 下同The same below. a. 种数Species number; b. 个体数Individuals number; c. 相对多度Relative abundence (%).
2.2 不同生境昆虫群落相似性分析
以各生境昆虫群落种类数量为属性的聚类分析(图1)表明,基于不同土壤类型的4个人工落叶松林生境昆虫群落分为2类(图2),A生境和B生境归为一类,群落相似性较大, C生境和D生境归为一类,群落较为相似。同时聚类结果也表明,2个类群之间昆虫群落物种数量组成差异较大。

图1 不同生境昆虫的群落聚类Fig.1 Cluster dendrogram of insect community in different types habitat
2.3 不同生境昆虫群落多样性特征
由表2可知,不同生境间昆虫群落多样性特征指数均无显著差异,物种多样性指数和均匀度指数大小均表现为样地C>A>B>D,丰富度指数大小表现为样地B>D>A>C,优势集中性指数大小表现为D>B>A>C。C生境昆虫群落物种多样性指数和均匀度指数均最高,丰富度指数最低,A生境具有较高的物种多样性指数和均匀度指数,丰富度指数相对较低,B生境昆虫群落物种多样性指数和均匀度指数相对较低,丰富度指数最高,D生境昆虫群落物种多样性指数和均匀度指数均最低,丰富度指数相对较高,各样地群落优势集中性指数大小趋势与均匀度指数相反。各多样性特征指数间的相关性分析表明,昆虫群落在不同样地间分布的水平结构上,物种多样性指数与均匀度指数(r=0.908)表现为较大的正相关,与丰富度指数(r=-0.545)和群落优势集中性指数(r=-0.803)表现为较大的负相关,均匀度指数与群落优势集中性指数(r=-0.94)表现为较大的负相关。

表2 不同生境昆虫群落各多样性特征指数Tab.2 Diversity index of insect community in different habitats
2.4 不同生境昆虫群落多样性时间动态
昆虫群落物种多样性指数(F=8.397,P=0.000)、均匀度指数(F=2.773,P=0.029)、丰富度指数(F=6.410,P=0.000)、优势集中性指数(F=4.044,P=0.005)均随时间变化表现出极显著的差异。各生境昆虫群落多样性指数时间动态如图2所示,各生境均在7月中旬表现出最低或较低的指数值,5月末至7月中旬期间,A生境表现出一定的波动,B、C和D生境均逐渐升高至6月中旬后降低; 7月中旬至9月中旬期间,A和C生境昆虫群落多样性指数逐渐升高至8月末后逐渐降低,B和D生境具有较大的波动性,出现2个高峰值。各生境多样性指数最高值除B样地出现在7月末外,其他生境均出现在8月末。
各生境昆虫群落均匀度指数时间动态表现为: A和C生境在6月中旬至6月末期间变化趋势相反,其他时期变化趋势均一致,表现为较大的正相关性(r=0.658),在全年均具有较大的波动性; B和D生境在7月中旬至7月末期间变化趋势相反,其他时期均匀度变化趋势均保持一致,表现为显著的正相关性(r=0.722,P<0.05),全年均具有较大波动,出现3个高峰值。各样地均匀度指数最高值均出现在9月末。
各生境昆虫群落丰富度指数时间动态表现为: A与D生境除6月中旬至6月末、8月末至9月中旬变化趋势有一定差异,其他时期均保持相同的变化趋势,表现为显著的正相关(r=0.748,P<0.05),C和D生境除6月中旬至6月末变化趋势相反,其他时期均保持相似的变化趋势,表现为显著的正相关(r=0.736,P<0.05),A和C生境丰富度指数变化趋势在5月末至8月中旬一致,表现为较大的正相关(r=0.656),随后变化趋势产生较大的差异。B生境变化趋势在5月末至7月中旬与A、C生境相似,8月中旬至9月中旬与A生境相似,在7月中旬至8月中旬波动最显著。
各生境昆虫群落优势集中性指数时间动态表现为: B和C生境变化趋势在5月末至7月末均保持一致,随后C生境保持平稳趋势,B生境波动较大; D生境与B、C生境在5月末至7月中旬变化趋势相似,随后变化趋势与其他样地产生较大差异,同时D生境在全年大部分时期均与A生境呈现相反的变化趋势。
各生境中昆虫群落多样性各特征指数间相关分析表明(表3),各生境多样性指数与群落优势集中性指数均表现为极显著的负相关,与均匀度指数除C生境外均表现为显著正相关,与丰富度指数除A生境外均表现为极显著的正相关。均匀度指数与群落优势集中性指数除C生境外均表现为极显著的负相关,但各生境均出现6月末至7月中旬均匀度指数与优势集中性指数变化趋势相似的现象。

表3 不同多样性指数的相关系数(r)①Tab.3 Correlation coefficients (r) of different diversity indices
①*P<0.05,**P<0.01.
2.5 不同生境昆虫群落营养类群及稳定性分析
各生境昆虫群落中不同营养类群物种数均表现为植食性类群>中性类群>捕食性类群>寄生性类群(表4),其中植食性昆虫物种数在各自样地中所占比例为A最高,B最低; 中性昆虫物种数所占比例为B和D较高,A和C较低; 捕食性昆虫物种数所占比例为B>C>D>A; 寄生性昆虫物种数所占比例为C生境略高,其他样地比例相差不大。
采用高宝嘉等(1992)提出的昆虫群落相对稳定性指标(St/Si和Sn/Sp)对昆虫群落相对稳定性进行分析(表5),各样地昆虫群落St/Si值均较高,Sn/Sp值表现出一定的差异,B生境为最高值,A生境为最低值,B生境比A生境增加57.58%。综合St/Si和Sn/Sp值,各生境昆虫群落相对稳定性值为B>C>D>A。

图2 不同样地昆虫群落多样性指数(H′)、均匀度指数(J)、物种丰富度(R)、优势集中性指数(C)的时间动态Fig.2 Dynamics of the diversity index (H′),evenness index(J), specific richness(R), dominance index (C) of insect community in different habitats

昆虫类群InsecttaxaABCDabababab捕食性类群Predatory1314142311191218寄生性类群Parasitic312413313313植食性类群Defoliator2566275823542157中性类群Neutral1228173611271434
①a: 物种数 Species number; b: 所占比例 Proportion.
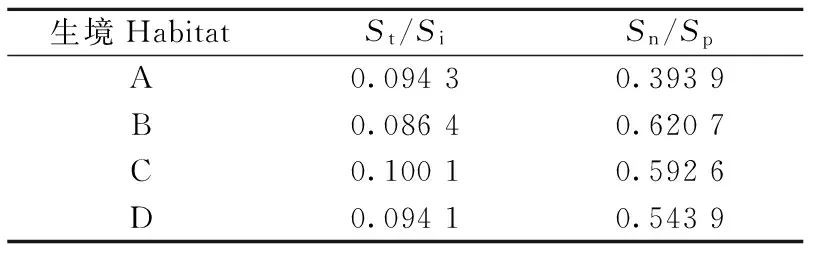
表5 不同样地昆虫群落相对稳定性Tab.5 Relative stability of insect communities in different types habitat
2.6 不同生境昆虫群落主分量分析
对各生境昆虫群落进行主分量分析(表6)。A生境昆虫群落的第1主分量代表寄生性昆虫个体数、捕食性昆虫物种数和个体数的综合因子,对昆虫群落的变化起着主导作用; 第2主分量代表植食性昆虫个体数和物种数的综合因子。前2个主分量累计贡献率80.89%,天敌昆虫和植食性昆虫共同影响昆虫群落的波动。
B生境昆虫群落的第1主分量代表中性昆虫物种数、捕食性昆虫物种数、寄生性昆虫物种数和个体数、植食性昆虫物种数和个体数的综合因子,是影响昆虫群落变化的主要因子; 第2主分量代表中性昆虫个体数和植食性昆虫物种数的综合因子; 第3主分量代表捕食性昆虫个体数和中性昆虫个体数的综合因子,前3个主分量累计贡献率85.06%。
C生境昆虫群落的第1主分量代表寄生性昆虫个体数、中性昆虫个体数和捕食性昆虫物种数的综合因子,是影响昆虫群落变化的主要因子; 第2主分量代表寄生性昆虫物种数、捕食性昆虫个体数、植食性昆虫物种数和个体数的综合因子; 第3主分量代表中性昆虫物种数、植食性昆虫物种数和捕食性昆虫物种数的综合因子,前3个主分量累计贡献率达92.45%。
D生境昆虫群落的第1主分量代表寄生性昆虫物种数和个体数、植食性昆虫个体数、中性昆虫物种数的综合因子,是影响昆虫群落变化的主要因子; 第2主分量代表捕食性昆虫个体数和物种数的综合因子,前2个主分量累计贡献率73.96%。

表6 不同样地昆虫群落的主分量分析①Tab.6 Principal component analysis of insect community in different types habitat
①X1: 捕食性昆虫种数Species number of predatory insects;X2: 捕食性昆虫个体数Individual number of predatory insects;X3: 寄生性昆虫种数Species number of parasitic insects;X4: 寄生性昆虫个体数Individual number of parasitic insects;X5: 植食性昆虫种数Species number of phytophagous insects;X6: 植食性昆虫个体数Individual number of phytophagous insects;X7: 中性昆虫种数Species number of neutral insects;X8: 中性昆虫个体数Individual number of neutral insects.
3 讨论
4种不同土壤生境的兴安落叶松人工林中昆虫群落优势目均为双翅目和膜翅目,但各样地中优势目个体数所占比例具有较大差异,典型暗棕壤落叶松林中具有最高比例的双翅目和最低比例的膜翅目,潜育暗棕壤落叶松林中双翅目比例最低,埋藏暗棕壤落叶松林中膜翅目比例最高。各生境优势科表现为从A生境到B生境,逐渐从以膜翅目蚁科和姬蜂科为优势科,渐变为膜翅目蚁科、姬蜂科和双翅目厕蝇科、菌蚊科为共同优势科,到C生境和D生境,双翅目菌蚊科的优势度逐渐增大为主优势科,膜翅目蚁科逐渐减小为常见类群。各生境不同季节优势科变化规律也表现为A生境和B生境在全年大部分时期以蚁科为主要优势科,C生境和D生境菌蚊科为优势科的时期占据了全年较长时间。对不同生境昆虫群落的聚类分析也表明,埋藏潜育暗棕壤和厚层埋藏暗棕壤落叶松人工林的昆虫群落相似性较大,暗棕壤性土和典型暗棕壤落叶松人工林昆虫群落相似性较大,土壤类型对落叶松人工林昆虫群落组成具有较大影响。
各生境多样性指数均较高,昆虫群落生存的环境较为优越。多样性指数与均匀度、丰富度指数的关系在不同的研究中结果各异,贺答汉等(1988)认为荒漠草原昆虫群落的多样性指数与均匀度是一致的, 表明群落结构是稳定的。万方浩等(1986)研究的稻田昆虫群落则得出在不同季节多样性指数与均匀度不一致。在本研究中,昆虫群落在不同生境间水平分布结构上,物种多样性指数与均匀度指数表现为较大的正相关,与丰富度指数表现为较大的负相关,说明两者对多样性指数均具有较大的影响。各生境昆虫群落时间结构上,厚层埋藏暗棕壤样地和典型暗棕壤样地昆虫群落物种多样性指数与均匀度指数、丰富度指数随时间变化趋势均表现为极显著或显著的正相关,但前者各相关系数明显高于后者,埋藏潜育暗棕壤样地和暗棕壤性土样地多样性指数均仅与均匀度指数或丰富度指数其中之一为显著正相关。说明各生境昆虫群落时间结构上,厚层埋藏暗棕壤落叶松林昆虫群落对环境的适应性更强,生存环境更为优越。
厚层埋藏暗棕壤落叶松林昆虫群落稳定性最强,埋藏潜育暗棕壤落叶松林昆虫群落稳定性最弱,前者多样性指数和均匀度指数均最低,但丰富度指数最高,且各指数时间动态变化趋势均为极显著正相关,后者仅有均匀度指数与多样性指数时间变化趋势较一致,且相关系数相对于其他群落较小。暗棕壤性土落叶松林昆虫群落相对稳定性大于典型暗棕壤落叶松林,两者多样性指数与丰富度指数时间动态变化趋势均极显著的一致,但前者相关系数显著大于后者,进一步说明在该区域,昆虫群落物种多样性指数与丰富度指数正相关性越显著,群落稳定性越强。
结合昆虫群落主分量分析,相对稳定性最强的厚层埋藏暗棕壤落叶松林中性昆虫物种数是主导昆虫群落变化最主要因子,同时捕食性昆虫物种数、寄生性昆虫和植食性昆虫均对昆虫群落起到较大影响,各类群较为均衡,随着群落相对稳定性降低,中性昆虫作为主导因子的影响作用逐渐降低,到相对稳定性最弱的埋藏潜育暗棕壤落叶松林中主导因子变为寄生性昆虫个体数和捕食性昆虫,中性昆虫对群落无显著影响,可见小兴安岭南坡落叶松人工林中性昆虫对群落稳定性的影响极为显著。
不同土壤条件对落叶松人工林昆虫群落产生直接或间接的重要影响,引起昆虫群落结构及动态变化的差异,厚层埋藏暗棕壤落叶松林昆虫群落最为稳定,反映该生境环境条件最优越,更利于昆虫群落的生存发展。对于埋藏潜育暗棕壤落叶松人工林,需要采取一定措施增加林下植物种类(Randlkoferetal., 2010),以此提升中性昆虫和天敌昆虫的比例,增强昆虫群落内部食物网络关系的复杂性,增强昆虫群落稳定性。
4 结论
不同土壤条件下落叶松人工林各昆虫群落结构差异较大,昆虫群落物种多样性指数与丰富度指数正相关系数越大,群落稳定性越强,同时中性昆虫对昆虫群落的相对稳定性起到显著的调控作用。4种生境中厚层埋藏暗棕壤落叶松人工林综合环境条件最适宜昆虫群落发生发展。
陈天乙. 1995.生态学基础教程. 天津: 南开大学出版社, 116-130.
(Chen T Y. 1995. Ecologybasic. Tianjin: Nankai University Press, 116-130. [in Chinese])
杜秀娟,任炳忠,吴艳光. 2009.长白山北坡不同海拔及干扰程度下访花食蚜蝇群落的差异.昆虫学报, 52(5): 551-560.
(Du X J,Ren B Z,Wu Y G.2009. Differences of flower-visiting hoverfly(Diptera: Syrphidae) communities in habitats with various degrees of disturbance and altitude in Changbai Mountain,N. E. China. Acta Entomologica Sinica,52(5): 551-560. [in Chinese])
高宝嘉,张执中,李镇宇. 1992.封山育林对昆虫群落结构及多样性稳定性影响的研究.生态学报, 12(1): 1-7.
(Gao B J,Zhang Z Z,Li Z Y.1992.Studies on the influence of the closed forest on the structure,diversity and stability of insect community.Acta Ecologica Sinica,12(1): 1-7. [in Chinese])
顾 伟,马 玲,刘哲强,等. 2014. 凉水自然保护区不同林型地表甲虫物种多样性. 东北林业大学学报,42(1): 131-135.
(Gu W, Ma L, Liu Z Q,etal. 2014. Diversity of ground-dwelling beetle with different forest type in liangshui Nature Reserve. Journal of Northeast Forestry University,42(1): 131-135. [in Chinese])
顾 伟,马 玲,丁新华,等. 2011.扎龙湿地不同生境的昆虫多样性.应用生态学报, 22(9): 2405-2412
(Gu W, Ma L, Ding X H,etal. 2011. Insect diversity of different habitat types in Zhalong Wetland,Northeast China.Chinese Journal of Applied Ecology, 22(9): 2405-2412. [in Chinese])
贺答汉,田 畴,任国栋,等.1988.荒漠草原昆虫的群落结构及其演替规律初探.中国草地, (6): 24-28.
(He D H, Tian C, Ren G D,etal.1988. A preliminary study on the structure of desert forage grassland.Chinese Journal of Grassland, (6): 24-28. [in Chinese])
姬兰柱, 董百丽,魏春艳,等. 2004.长白山阔叶红松林昆虫多样性研究.应用生态学报,15 (9): 1527-1530
(Ji L Z, Dong B L, Wei C Y,etal. 2004. Insect species diversity in Korean pine broad-leaved mixed forest in Changbai Mountains. Chinese Journal of Applied Ecology, 15 (9): 1527-1530. [in Chinese])
贾玉珍,赵秀海,孟庆繁. 2009.长白山针阔混交林不同演替阶段的昆虫多样性.昆虫学报, 52(11): 1236-1243
(Jia Y Z,Zhao X H,Meng Q F.2009.Insect diversity along a successional gradient in conifer and broad-leaved mixed forests in Changbai Mountain. Acta Entomologica Sinica, 52(11): 1236-1243. [in Chinese])
李孟楼,刘朝斌. 1995. 秦岭南坡火地塘林区落叶松人工林的昆虫群落及其结构. 西北林学院学报,10(1): 68-73.
(Li M L,Liu C B. Insect community and structure in larch forest on south slope of Qinling mountain. Journal of Northwest Forestry Cellege,10(1): 68-73. [in Chinese])
刘哲强,马 玲,焦 玥,等. 2015. 小兴安岭森林恢复期典型人工林与原始林昆虫群落结构动态. 应用生态学报,26(2): 555-562.
(Liu Z Q, Ma L, Jiao Y,etal. 2015. Structure dynamics of insect communities in typical artificial and primeval forests during restoring stages in Xiaoxing’an Mountain,Northeast China.Chinese Journal of Applied Ecology, 26(2): 555-562. [in Chinese])
马建章,刘传照. 1993. 凉水自然保护区研究. 哈尔滨: 东北林业大学出版社,18-26.
(Ma J Z, Liu C Z. 1993. The Researches of Liangshui Natural Reserve. Harbin: Northeast Forestry University Press, 18-26. [in Chinese])
马 玲,顾 伟,王利东,等. 2012. 扎龙湿地的昆虫群落生态位.林业科学, 48(5): 81-87.
(Ma L, Gu W, Wang L D,etal. 2012. Insect community niche in the Zhalong wetland. Scientia Silvae Sinicae, 48(5): 81-87. [in Chinese])
孙儒泳,李 博,诸葛阳,等. 1997.普通生态学.北京: 高等教育出版社, 416-426.
(Sun R Y, Li B, Zhuge Y,etal. 1997. General Ecology. Beijing: Higher Education Press, 416-226. [in Chinese])
万方浩,陈常铭.1986.综防区和化防区稻田害虫-天敌群落组成及多样性的研究.生态学报, 6(2): 159 -170.
(Wan F H, Chen C M. 1986. Studies on the structure of the rice pest-natural enemy community and diversity under IPM area and chemical control area. Acta Ecologica Sinica, 6(2): 159 -170. [in Chinese])
余世孝. 1995.数学生态学导论.北京: 科学技术文献出版社.
(Yu S X. 1995. Introduction to Mathematical Ecology.Beijing: Science and Technology Document Press. [in Chinese])
赵东芳,王志明,钱 晨. 2011. 长白落叶松人工林昆虫多样性的研究. 安徽农业科学, 39(12): 7056 -7059.
(Zhao D F, Wang Z M, Qian C. 2011. Study on the diversity of insects in planted forest ofLarixolgensisHenry. Journal of Anhui Agri Sci, 39(12): 7056 -7059. [in Chinese])
赵志模, 郭依泉. 1990.群落生态学原理与方法.重庆: 科学技术文献出版社重庆分社, 1-46.
(Zhao Z M, Guo Y Q. 1990.Principle and Method of Community Ecology. Chongqing: Chongqing Branch of Scientific and Technological Document Press, 1-46.[in Chinese])
Baars M A.1979.Catches in pitfall traps in relation to mean densities of carabid beetles. Oecologia,41: 25-46.
Randlkofer B,Obermaier E,Hilker M,etal.2010.Vegetation complexity: The influence of plant species diversity and plant structures on plant chemical complexity and arthropods.Basic and Applied Ecology,11: 383-395
Samways M J.1993. Insects in biodiversity conservation: Some perspectives and directives.Biodiversity and Conservation, 2(3): 258-282.
(责任编辑 朱乾坤)
Variation in Structure and Dynamics of Insect Community in Larch Plantations under Different Soil Conditions
Gu Wei1,2Ma Ling1Sun Hu2Wang Lidong2Zhang Zilong1
(1.School of Forestry, Northeast Forestry University Harbin 150040; 2.Liangshui Nature Reserve of Northeast Forestry University Yichun 153106)
【Objective】This study aimed to investigate the variation in insect community structure and dynamics in larch plantations under different soil conditions, so as to provide fundamental basis for the forest management and biodiversity conservation.【Method】This experiment was conducted at Liangshui Nature Reserve, in Xiaoxing’anling Mountains, China. The 60-year-old larch plantation, which was in the condition of close hillsides, was used for this. According to the terrain soil conditions 4 habitats were chosen in the forest, and the 4 habitats are buried-gley dark brown soil-larch plantation forest(A), thick buried dark brown soil-larch plantation forest(B), dark brown loam soil-larch plantation forest (C), typical dark brown soil-larch plantation forest(D). Three 20 m × 20 m plots were established in each habitat. Insect community was investigated by sweeping net method, the tin pot trap method, shaking-off method and high branch cutting standard branch method.【Result】A total of 5 204 insects were collected, belonging to 250 species, 77 families, and 10 orders. Diptera and Hymenoptera were the dominant taxa in each habitat, but the proportion had great difference. There was great difference in seasonal dynamics of the dominant family of insect communities over the four habitats. Cluster analysis showed that insect communities of the 4 habitats were classified into two categories, habitat A and habitat B were classified into one category, habitat C and habitat D were classified into the other category. Species diversity index and evenness index of insect communities in different habitats showed C>A>B>D, richness index showed B>D>A>C, dominant index showed D>B>A>C. In time series, the species diversity index of the B habitat and the D habitat was significantly positively correlated with the evenness index and richness index, but the former correlation coefficient was significantly higher than that of the latter. In all habitats, species number order of different trophic groups showed plant feeding taxa > neutral group > predators group > parasitic taxa, but the proportion of different trophic groups had significant difference. The relative stability of the insect communities in different habitats was B >D>C>A. Principal component analysis showed that the factors that played a dominant role in affecting the insect community change were different in different habitats.【Conclusion】The structure dynamics of the insect communities in larch plantations were significantly different under different soil conditions. The higher the positive correlation coefficient of species diversity index and richness index of insect community, the stronger the community stability. At the same time, neutral insects played a significant role in regulating the relative stability of insect communities. Environment conditions of thick buried dark brown soil-larch plantation were most suitable for the occurrence and development of insect community in the 4 habitats.
insect community; stability; principal component analysis; larch plantation; Liangshui Nature Reserve
10.11707/j.1001-7488.20170512
2015-11-08;
2016-05-25。
中央高校基本科研业务费专项资金(2572014BA06);黑龙江省自然科学基金项目(ZD201404)。
S718.7
A
1001-7488(2017)05-0097-10
*马玲为通讯作者。
