盐胁迫下木炭对小麦幼苗根系蛋白表达的影响
2017-06-22陈计芳段江燕
陈计芳,段江燕
(山西师范大学生命科学学院,山西临汾 041000)
盐胁迫下木炭对小麦幼苗根系蛋白表达的影响
陈计芳,段江燕
(山西师范大学生命科学学院,山西临汾 041000)
为明确木炭对小麦盐胁迫的缓解效应,以冬小麦品种临汾6339为材料,对小麦幼苗分别进行盐和木炭单因素处理及二者复合处理,应用蛋白质双向电泳与质谱联用技术对小麦根系差异表达蛋白进行了分析。结果表明,相对盐处理,在盐和木炭复合处理的小麦根系中共检测到丰度变化在2倍以上的差异表达蛋白点12个。经过MALDI-TOF-TOF分析及数据库检索,被鉴定的12个蛋白点按其功能大致可归为5类。其中,第Ⅰ类为与能量代谢相关的蛋白质,包括ATP合酶和ATP酶;第Ⅱ类为与糖代谢相关的蛋白质,包括磷酸甘油醛异构酶、磷酸丙糖异构酶、6-磷酸葡糖酸脱氢酶、苹果酸脱氢酶以及乌头酸水合酶;第Ⅲ类为与氨基酸代谢相关的蛋白质,如半胱氨酸合成酶;第Ⅳ类为与遗传物质有关的蛋白质,如染色体的结构蛋白;第Ⅴ类为未知功能蛋白质。
小麦;根系;盐胁迫;木炭;蛋白表达
土壤盐渍化影响植物正常生长发育[1],因而一直是世界农业发展研究关注的问题。研究表明,NaCl等环境胁迫条件会使植物部分蛋白发生变性[2-7]。施用NO供体SNP[8]、稀土元素铈[9]、赤霉素[10]等物质可缓解盐胁迫对植株的伤害,但这些材料都普遍存在着价格贵、稀缺、难制备、不易用于生产等缺点。木炭是一种碳含量极其丰富的生物炭。它富含微孔,不但可以提高土壤有机质含量,还可以有效地保存水分和养料,提高土壤肥力,已广泛应用于固碳减排、水源净化、重金属吸附、土壤改良等方面。有研究发现,生物炭对土壤盐分表聚现象有显著的影响,但影响效果与生物炭添加量密切相关。当添加量较低(不超过5%)时,抑制土壤蒸发,降低土壤表层返盐量;当添加量较大时(不小于10%),增强土壤蒸发能力,加剧表层土壤盐碱化的程度[11]。生物炭浸提液可以提高盐胁迫下水稻幼苗的抗氧化能力和对盐胁迫的耐受性[12]。施用棉秆碳可有效改良新疆盐渍化土壤的理化性质,改善土壤特性,在提高土壤质量的同时增加作物产量[13]。在回收利用的废水污泥土壤中,加入木炭能减少温室气体排放,并能促进小麦生长和增产[14]。Díaz-Rojas等[5]的研究也得出同样的结论。施用0.05%的木炭对甜椒的小孢子花粉粒胚的形成有明显促进作用[16]。在木薯培养过程中,加入木炭可促进木薯的萌发和细胞的分化。然而,有关木炭对植物抗氧化酶活性、蛋白质含量及表达等方面的影响[17],目前尚未见报道。本研究拟借助蛋白质组学技术,分析在NaCl胁迫下加入木炭后小麦幼苗根部蛋白的差异表达,以期从蛋白质水平揭示木炭对小麦受到的盐胁迫伤害的缓解效应。
1 材料与方法
1.1 供试材料
供试冬小麦为品系6339,种子由山西省农科院小麦研究所提供。木炭为市场上比较普遍的阔叶木炭。
1.2 试验设计
选取饱满、大小均一的冬小麦种子,将其用70%~75%的酒精表面消毒0.5~1.0 min后,用无菌水漂洗数次,用MilliQ水浸种24~48 h,待其露白后,腹沟朝下,均匀排列于铺有两层滤纸的大培养皿里,置于培养箱中培养,每天光照10 h[18]。
根据预试验结果,将NaCl、木炭分别按照1.0%和4.0%的比例与营养土混匀后装入花盆,以不加盐和木炭为对照,共形成4个处理,即CK、NaCl、木炭及NaCl+木炭处理。待小麦长到两叶一心时移栽到花盆中,9 d后取出小麦幼苗,剪根称量待用。
1.3 方法
1.3.1 小麦幼苗根部总蛋白的提取与制备
尿素/硫脲法为本实验室提取蛋白的常用方法,具体提取过程参照刘伟霞[19]的报道,但略有改善,即把提取液组分中的DTT省略掉。
1.3.2 小麦幼苗根部蛋白质的双向电泳
第一向固相pH 梯度等电聚焦电泳(Isoelectricfocusing,IEF):取出IPG预制胶条(7 cm,pH 3~10),将上样蛋白量和水化上样液混匀。等电聚焦程序:50 V水化16 h,100 V线性除盐1.5 h,300 V除盐升压1.5 h,1 000 V线性升压30 min,3 000 V快速升压30 min,6 000 V聚焦20 000 Vh,500 V保持。
第二向SDS-PAGE 电泳:聚焦完毕后,取出胶条,在10 mL平衡缓冲液Ⅰ(6 mol·L-1Urea、2% SDS、1.5 mol·L-1Tris-HCl(pH 8.8)、20%Glycerinum、2% DTT)和10 mL平衡缓冲液Ⅱ(6 mol·L-1Urea、2%SDS、1.5 mol·L-1Tris-HCl、20%Glycerinum、2.5%Iodoacetamide)的溶胀盘里摇晃15 min。将胶条放到第二向SDS-PAGE凝胶的上表面,将凝胶转移至电泳槽中,开始选用80 V,待观测到蛋白样品条带成一条较平缓的直线后改换电压120 V继续电泳,等到溴酚蓝指示剂的蓝色出现在胶底部时,停止电泳。
1.3.3 考马斯亮蓝染色
电泳结束后,取出胶条转入500 mL固定液(95%Alcoho,10.0%Glacial acetic acid.)固定30 mim后,选用考马斯亮蓝染色法(0.1%Coomassie brilliant blue R-250、95% Alcohol、10.0%Glacial acetic acid 定容至1 000 mL)染色2 h,后转至1 000 mL脱色液(95%Alcohol、5.0%Glacial acetic acid),期间不断更换脱色液,直至条带清晰可见。
1.3.4 双向电泳图谱分析
将已染色的凝胶用扫描仪进行扫描,并使用PDQuest 8.0.1软件对图像进行分析,将经过裁剪、标记的各实验组图像进行自动检测和匹配,以2倍表达差异为标准 (如2倍上升表达和2倍下降表达) 选择差异蛋白点进行质谱鉴定。
1.3.5 质谱鉴定与数据库检索
切下胶点,利用质谱仪MicrOTOF-QII(BrukerDaltonics)鉴定蛋白质。用Data Analysis Software和Mascot search engine version 2.3.01对质谱数据进行分析和搜索(通常使用软件MASCOT在TAIR数据库中检索,对所搜索结果进行分析鉴定,得分低于60者,定位为假阳性蛋白,应舍弃)。
1.3.6 生物信息学分析
依据UniProtKB数据库(http://www.uniprot.org/)提供的蛋白质注释信息对蛋白质进行功能分类。
2 结果与分析
2.1 不同处理下小麦根部差异蛋白点的比较
通过2-DE技术,共检测出100多个蛋白点,这些蛋白点pH值和分子量分别主要集中于3.5~7.5和30~85 kD范围(图1、图2)。其中,与CK相比,木炭处理(C)蛋白点表达上调8个,表达下调10个,未表达10个;NaCl处理(Y)蛋白点表达上调1个,表达下调18个,未表达11个;NaCl+木炭复合处理(F)蛋白点表达上调5个,表达下调12个,未表达13个。而与Y处理相比,F处理蛋白点表达上调15个,表达下调0个,未表达4个。

C:木炭;Y:NaCl;F:NaCl+木炭。图2同。
C: Charcoal; Y: NaCl; F: NaCl+Charcoal.The same in Fig.2.
图1 不同处理的蛋白表达图谱
Fig.1 Images of protein spots under different treatments
2.2 小麦根部差异表达蛋白点的质谱鉴定
选取复合处理中丰度变化相对于盐处理在2倍以上的12个差异表达蛋白点进行MALDI-TOF-TOF/MS质谱鉴定(表1)。依据UniProtKB数据库中对应蛋白点的信息资料,将所鉴定的12个差异表达的小麦幼苗根系蛋白按其功能归为五类。第Ⅰ类为与能量代谢相关的蛋白,即ATP酶(Vacuolar proton-ATPase subunit A,SSP1901和SSP1701);第Ⅱ类为与糖代谢相关的蛋白,包括磷酸甘油醛异构酶(Phosphoglycerate kinase,SSP1902和SSP2903)、磷酸丙糖异构酶(Triosephosphate isomerase, SSP1401)、6-磷酸葡糖酸脱氢酶(6-phosphogluconate dehydrogenase,decarboxylating,SSP1502)、苹果酸脱氢酶(Malic enzyme,SSP4801)以及乌头酸水合酶(Aconitate hydratase,SSP4901);第Ⅲ类为与氨基酸代谢相关的蛋白,如半胱氨酸合成酶(Cysteine synthase,SSP1503);第Ⅳ类为与遗传物质相关的蛋白,如染色体的结构蛋白(Chromosome 3B,genomic scaffold,cultivar Chinese Spring,SSP0501、SSP1903和SSP3601);第Ⅴ类为未知功能的蛋白质。
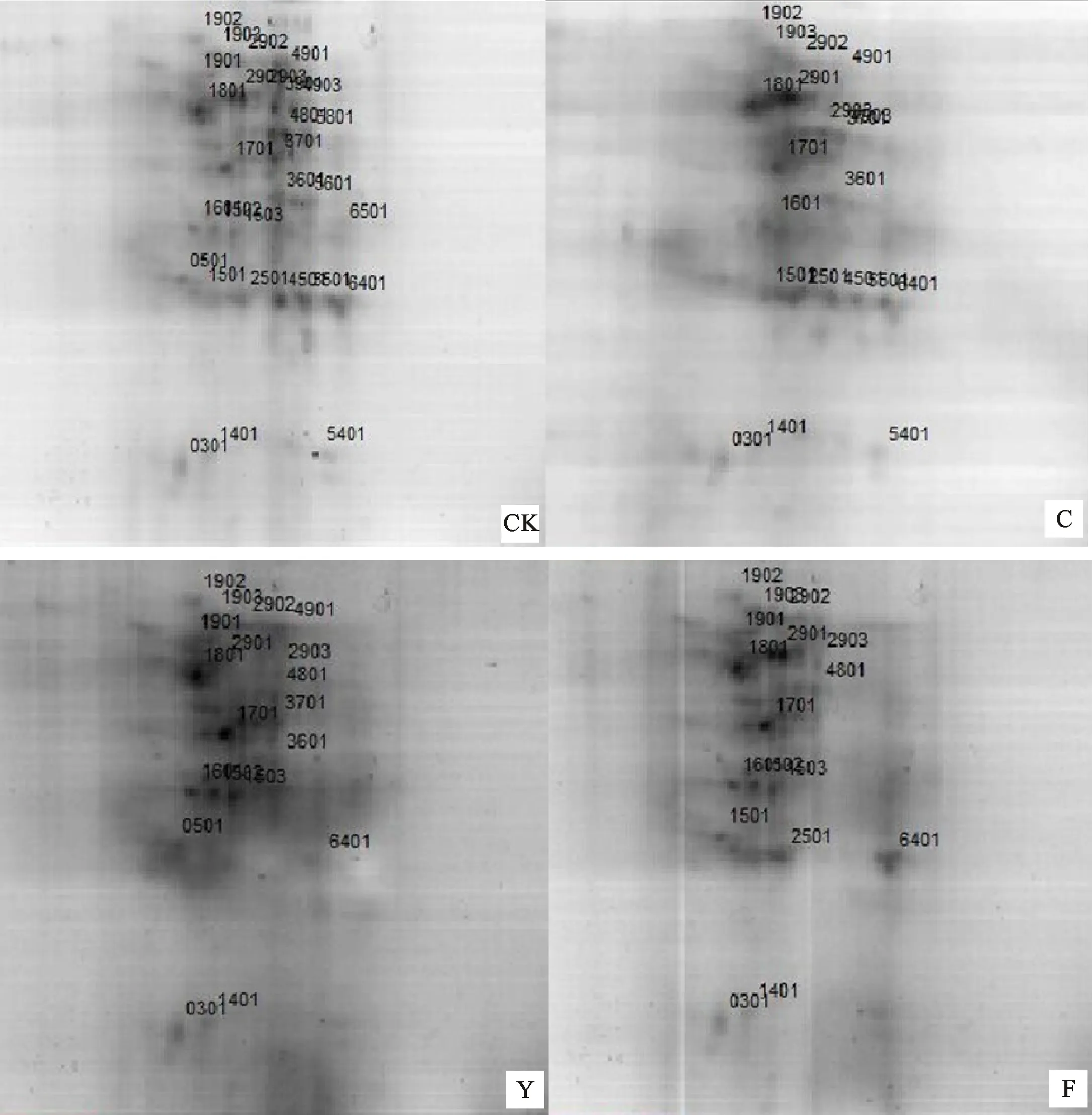
图2 不同处理的蛋白点命名图谱
3 讨 论
3.1 盐胁迫及木炭对小麦幼苗根部形态结构和生理生化的影响
盐胁迫能导致植株矮化,降低干重和鲜重,影响植物的光合作用,使植物提前发育并过早衰老[20],引起作物减产[21],影响植物体内离子运输。这是因为在盐胁迫的应答过程中,高浓度的NaCl改变碳水化合物代谢和能量代谢、离子的膜转运和平衡,使细胞骨架重组和细胞壁重构[22]。研究发现,少量的盐胁迫能促进植物抗氧化酶系统增强,提高SOD、POD和CAT活性;而过高含量的盐分则使植物体内抗氧化酶活性受到抑制。这一现象被很多学者已证实。周丹丹研究表明,5%盐浓度胁迫下,随着盐胁迫时间的延长,渗透调节物质(可溶性糖、脯氨酸和可溶性蛋白质)含量有所下降,SOD、POD、CAT 活性随着胁迫时间的延长先升高后降低[23]。NaCl对冬小麦的抗氧化酶活性的抑制作用与植物体内ROS含量剧增有关[24-25]。而木炭是近几年来迅猛发展起来的对环境无二次污染的新型肥料[15]。木炭的添加能增加植物新生根的数目,提高植物鲜重、株高和叶面积,促进植物器官分化,提高作物产量[14],同时也使植物体内MDA含量下降,显著增加SOD、POD和CAT活性。同时冬小麦在盐胁迫下根部可溶性糖有所积累,这可能是对盐胁迫的适应性表现,或是起信号物质的作用[26],还有待进一步研究。
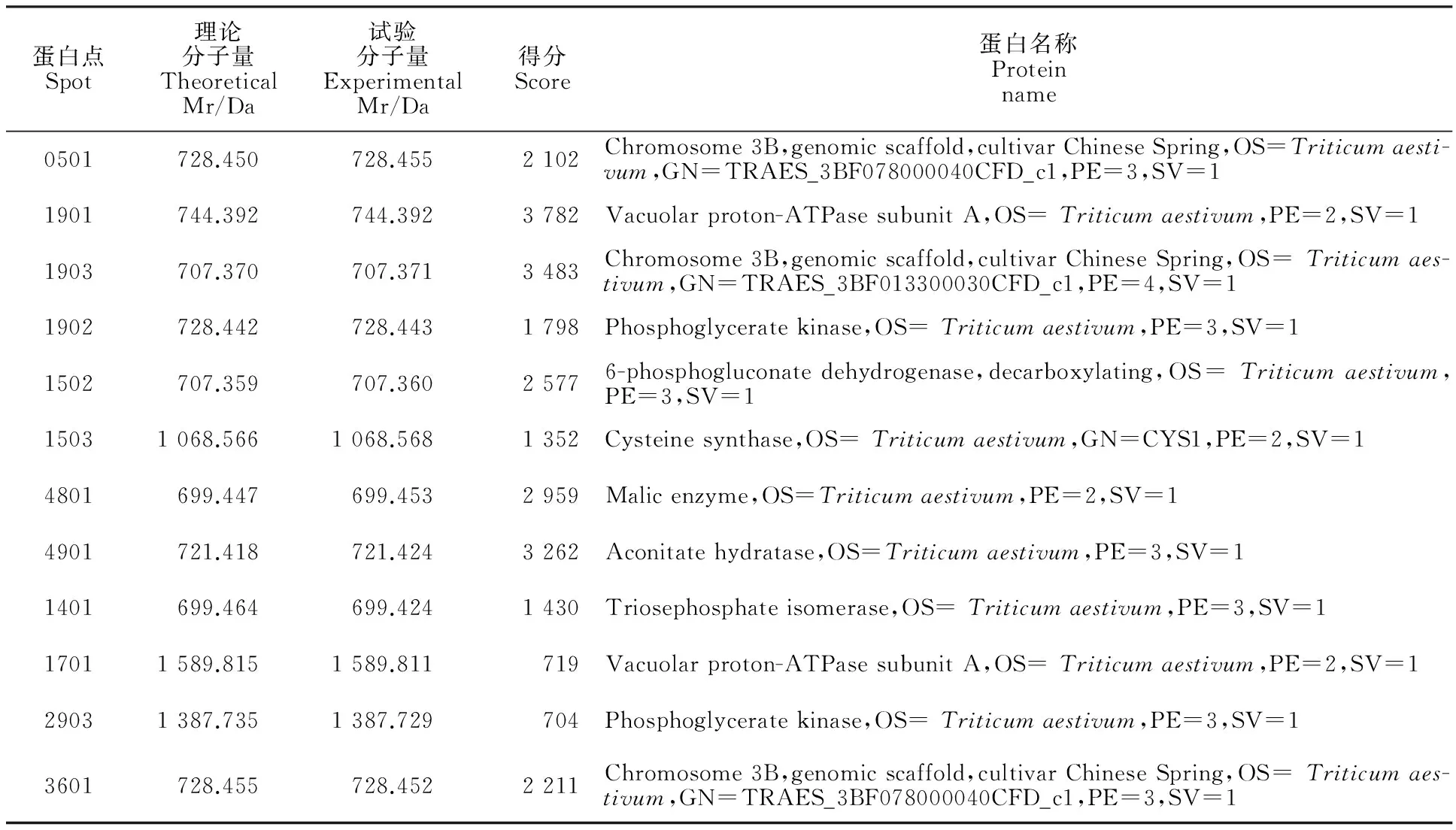
表1 盐胁迫及木炭处理下小麦幼苗根部差异蛋白点的MALDI-TOF-TOF/MS鉴定结果
3.2 木炭对盐胁迫下小麦幼苗根部差异蛋白表达的影响
本实验共成功鉴定出12个蛋白点,按其功能可归为五类蛋白质。
第Ⅰ类为与能量代谢相关蛋白质,即ATP酶(Vacuolar proton-ATPase subunit A,SSP1901和SSP1701 )。ATP酶广泛分布于线粒体内膜、叶绿体类囊体及异养菌和光合菌的质膜上,参与氧化磷酸化和光合磷酸化,在Na+的转运和亚细胞分布以及离子平衡中具有重要作用[27]。ATP合成酶与能量代谢紧密相关。这些蛋白质在盐胁迫下表达下调或不表达,而在木炭处理下有部分上调。研究表明,干旱条件下,ATP合酶表达下调,这将不可避免地影响光合磷酸化的产生,进一步影响卡尔文循环,这可能是干旱胁迫条件下植物净光合速率下降的一个主要原因[28]。
第Ⅱ类为与糖代谢相关蛋白质,包括磷酸甘油醛异构酶(Phosphoglycerate kinase,SSP1902,SSP2903)、磷酸丙糖异构酶(Triosephosphate isomerase,SSP1401)、6-磷酸葡萄糖酸脱氢酶(6-phosphogluconate dehydrogenase,decarboxylating,SSP1502)、苹果酸脱氢酶(Malic enzyme,SSP4801)以及乌头酸水合酶(Aconitate hydratase,SSP4901)。这些蛋白在受到盐胁迫时会表达下调或不表达,而木炭则可缓解这些胁迫,使部分蛋白点恢复上调。葡萄糖-6-磷酸脱氢酶是磷酸戊糖途径的限速酶。磷酸戊糖途径存在于细胞质中,是葡萄糖在体内生成5-磷酸核糖的唯一途径。后者是合成核苷酸辅酶及核酸的主要原料,故在损伤后修复及胁迫处理的细胞中,磷酸戊糖途径比较活跃。该蛋白的特异性表达可能对其修复盐胁迫造成的损伤具有一定作用[27]。磷酸丙糖异构酶是生物体中重要的异构酶之一,存在于多种组织中,其主要参与糖酵解过程,对于 ATP 的储存与生成是必不可少的[28]。研究发现,干旱逆境条件下,磷酸丙糖异构酶的表达量下调,说明在盐胁迫条件下,糖酵解途径受到了阻碍,影响了细胞内糖代谢和能量代谢的正常进行。
第Ⅲ类为与氨基酸代谢相关的蛋白质,如半胱氨酸合成酶(Cysteine synthase,SSP1503),盐胁迫抑制了其表达,而复合处理组则表达上调,表明盐胁迫阻碍了氨基酸相关蛋白质的合成。该蛋白的特异性表达说明盐胁迫下,氨基酸合成明显提高,促进了氮代谢,从而提高了它的耐盐能力[27]。
第Ⅳ类为与遗传物质相关的蛋白质,如染色体的结构蛋白(Chromosome 3B,genomic scaffold,cultivar Chinese Spring,SSP0501,SSP1903和SSP3601)这些蛋白点能够控制遗传表现,使染色质保持一定的结构。
第Ⅴ类为未知功能蛋白质。在后续的研究中,将对其进行进一步探讨。
[1] 杨少辉,季 静,王 罡,等.盐胁迫对植物影响的研究进展[J].分子植物育种,2006,4(Z1):139.
YANG S H,JI J,WANG G,etal.Study on the effects of salt stress on plants [J].MolecularPlantBreeding,2006,4(Z1):139.
[2] DU Z K,LI J M,ZHONG Z C,etal.A proteomic analysis ofArachishypogaealeaf in responses to enhanced ultraviolet-B radiation [J].ActaEcologicaSinica,2014,34(10):2589.
[3] 郭广芳,董 坤,高利艳,等.小麦根蛋白提取与双向电泳方法的优化与应用[J].植物遗传资源学报,2010,11(2):244.
GUO G F,DONG K,GAO L Y,etal.Optimization and application of wheat root protein extraction and two dimensional electrophoresis [J].JournalofPlantGeneticResources,2010,11(2):244.
[4] 阮松林,马华升,王世恒,等.植物蛋白质组学研究进展.Ⅰ.蛋白质组关键技术[J].遗传,2006,28(11):1472.
RUAN S L,MA H S,WANG S H,etal.Research progress of plant proteomics:I.Key technology of protein group [J].Inheritance,2006,28(11):1472.
[5] 白宇杰.杂交水稻盐胁迫响应差异蛋白质组分析 [D].大连:辽宁师范大学,2010:1.
BAI Y J.Differential proteome analysis of salt stress response in hybrid rice [D].Dalian:Liaoning Normal University,2010:1.
[6] 高庆华,曾祥然,贾 霖,等.水稻病程相关蛋白质在逆境胁迫下的表达研究[J].生物化学与生物物理进展,2013,40(11):1140.
GAO Q H,ZENG X R,JIA L,etal.The expression profiling of rice pathogenesisrelated proteins in seedling stage under environmental stresses [J].DevelopmentofBiochemistryandBiologicalPhysics,2013,40(11):1140.
[7] 熊军波.紫花苜蓿响应盐胁迫的蛋白质组研究[D].北京:中国农业科学院,2011:5-6.
XIONG J B.Study on the proteome of the response to salt stress in alfalfa [D].Beijing:Chinese Academy of Agricultural Sciences,2011:5-6.
[8] 陈 明,沈文飚,阮海华,等.一氧化氮对盐胁迫下小麦幼苗根生长和氧化损伤的影响[J].植物生理与分子生物学学报,2004,30(5):569.
CHEN M,SHEN W B,RUAN H H,etal.Effects of nitric oxide on root growth and oxidative damage of wheat seedlings under salt stress [J].JournalofPlantPhysiologyandMolecularBiology,2004,30(5):569.
[9] 吕春晖,牛红军,齐 菲,等.铈对盐胁迫下小麦种子萌发过程中生理指标的影响[J].农业机械化与电气化,2014(6):8.
LÜ C H,NIU H J,QI F,etal.Effects of cerium on physiological indexes of wheat seed germination under salt stress [J].AgriculturalMechanizationandElectrification,2014(6):8.
[10] 温福平,张 檀,张朝晖,等.赤霉素对盐胁迫抑制水稻种子萌发的缓解作用的蛋白质组分析[J].作物学报,2009,35(3):483.
WEN F P,ZHANG T,ZHANG Z H,etal.Proteomic analysis of the effect of GA on the mitigation of rice seed germination under salt stress [J].JournalofCropScience,2009,35(3):483.
[11] 许 健.生物炭对土壤水盐运移的影响[D].杨凌:西北农林科技大学,2016:6-7.
XU J.Study on the influence of biochar experimental on the migration of soil water and salt [D].Yangling:Northwest A&F University,2016:6-7.
[12] 甄晓溪,徐 凡,蒋太英,等.生物炭浸提液对盐胁迫水稻幼苗生长的调节作用机制[J].沈阳农业大学学报,2015,46(4):471.
ZHEN X X,XU F,JIANG T Y,etal.Mechanism of the regulating effect on biochar-leach liquor to rice seedlings under salt stress [J].JournalofShenyangAgriculturalUniversity,2015,46(4):471.
[13] 秦 蓓,王雅琴,唐光木,等.施用棉秆炭对新疆盐渍化土壤理化性质及作物产量的影响[J].新疆农业科学,2016,52(12):2290.
QIN B,WANG Y Q,TANG G M,etal.Effects of applying cotton stalk biochar to Xinjiang saline soil on the physical and chemical properties and crop yield [J].XinjiangAgriculturalSciences,2016,52(12):2290.
[14] AGUILAR-CHVEZ,DAZ-ROJAS M,CRDENAS-AQUINO M D R,etal.Greenhouse gas emissions from a wastewater sludge-amended soil cultivated with wheat(Triticumspp.L.) as affected by different application rates of charcoal [J].SoilBiologyandBiochemistry,2012,52:90.
[16] CHENG Y,MA R L,JIAO Y S,etal.Impact of genotype,plant growth regulators and activated charcoal on embryogenesis induction in microspore culture of pepper(CapsicumannuumL.)[J].SouthAfricanJournalofBotany,2013,88:306.
[17] GROLL J,GRAY V M,MYCOCK D J.Development of cassava(ManihotesculentaCrantz.) somatic embryos during culture with abscisic acid and activated charcoal [J].JournalofPlantPhysiology,2002,159(4):437.
[18] 杨颖丽,杨 宁,王 莱,等.盐胁迫对小麦幼苗生理指标的影响[J].兰州大学学报(自然科学版),2007,43(2):29.
YANG Y L,YANG N,WANG L,etal.Effects of salt stress on physiological indexes of wheat seedlings [J].JournalofLanzhouUniversity(NaturalScienceEdition),2007,43(2):29.
[19] 刘伟霞,潘映红.适用于小麦叶片蛋白质组分析的样品制备方法[J].中国农业科学,2007,40(10):2169.
LIU W X,PAN Y H.Sample preparation method for proteomic analysis of wheat leaves [J].JournalofAgriculturalScience,2007,40(10):2169.
[20] 陈健妙,郑青松,刘兆普,等.两种麻疯树苗对盐胁迫的生理生态响应[J].生态学报,2010,30(4):933.
CHEN J M,ZHENG Q S,LIU Z P.Two kinds ofJatrophasaplingson ecological and physiological response to salt stress [J].JournalofEcology,2010,30(4):933.
[21] 梁洪艳.黑龙江省春小麦耐盐性的研究[D].哈尔滨:东北农业大学,2007:8-9.
LIANG H Y.Study on salt tolerance of spring wheat in Heilongjiang province [D].Harbin:Northeast Agricultural University,2007:8-9.
[22] 刘海臣,杨文军,张春兰,等.非生物胁迫下植物根系的蛋白质组学研究进展[J].河南农业科学,2016,45(1):8.
LIU H C,YANG W J,ZHANG C L,etal.Advances in proteomics of plant roots under abiotic stress [J].HenanAgriculturalScience,2016,45(1):8.
[23] 周丹丹,李存华,杨庆山,等.盐胁迫对朴树叶片渗透调节物质及保护酶系统的影响[D].济南:山东大学,2016:5-8.
ZHOU D D,LI C H,YANG Q S,etal.Effects of salt stress on osmotic adjustment substances and protective enzyme system of Pushu leaves [D].Jinan:Shandong University,2016:5-8.
[24] MAGRINI-BAIR K A,CZERNIK S,PILATH H M,etal.Biomass derived,carbon sequestering,designed fertilizers [J].AnnualEnvironmentScience,2009,3:217.
[25]SINGH R P,JHA P,JHA P N.The plant-growth-promoting bacteriumKlebsiellasp.SBP-8 confers induced systemic tolerance in wheat(Triticumaestivum) under salt stress [J].JournalofPlantPhysiology,2015,184:57.
[26] 刘爱荣,张远兵,张雪平,等.麦冬生长及相关生理代谢对NaCl胁迫的响应[J].草地学报,2016,24(1):93.
LIU A R,ZHANG Y B,ZHANG X P,etal.NaCl stress on growth and physiological metabolism ofOphiopogonjaponicus[J].ActaAgrestiaSinica,2016,24(1):93.
[27] 彭振英.小麦渐渗系耐盐抗旱的蛋白质组学比较研究[D].济南:山东大学,2008:5-8.
PENG Z Y.Comparative study of protein and salt tolerance in wheat gradually infiltration system [D].Jinan:Shandong University,2008:5-8.
[28] 范锋贵.小麦幼苗叶片水分胁迫差异表达蛋白研究及其候选基因克隆[D].杨凌:西北农林科技大学,2012:5-7.
FAN F G.Study on differential expression of water stress in wheat seedling leaves and its candidate gene cloning [D].Yangling:Northwest A&F University,2012:5-7.
Effect of Charcoal on Protein Expression of Wheat Seedling Root under Salt Stress
CHEN Jifang,DUAN Jiangyan
(College of Life Science,Shanxi Normal University,Linfen,Shanxi 041000,China)
Winter wheat Linfen 6339 was used to determine the effect of charcoal on wheat seedling root under salt stress.The wheat seedlings were treated with single factor (salt or charcoal) and two factors (salt and charcoal). The differential expression proteins in wheat roots were analyzed by two-dimensional electrophoresis and mass spectrometry. The results showed that compared with single factor treatment,with the increase of NaCl and charcoal concentration,a total of 12 differentially expressed protein spots were detected in the root system of wheat with abundance variation more than 2 times. The identified 12 protein spots could be classified into 5 categories according to their functions through MALDI-TOF-TOF analysis.Among them,ATP synthase and ATP enzymes belonged to ClassⅠ,which are relative to energy metabolism. Class Ⅱ contained sugar metabolism related proteins,including phosphoglycerate kinase,triosephosphateisomerase,6-phosphogluconate dehydrogenase,malic enzyme and aconitatehydratase. Class Ⅲ contained cysteine synthase belonging to amino acid metabolism related proteins. Class Ⅳ contained structural proteins of chromosome,belonging to hereditary substance related proteins. Class Ⅴ contained unknown function proteins.
Wheat; Root;Salt stress; Charcoal;Protein expression
时间:2017-05-12
2016-12-04
2017-01-12
E-mail:2575814547@qq.com
段江燕(E-mail:506777055@qq.com)
S512.1;S311
A
1009-1041(2017)05-0705-07
网络出版地址:http://kns.cnki.net/kcms/detail/61.1359.S.20170512.2001.038.html
