丙磺舒在阿维菌素作用棉铃虫细胞中的增效机制研究
2017-06-21王赞永马海昊邓希乐谭辉华周小毛
王赞永,周 勇,马海昊,刘 佳,朱 航,邓希乐,谭辉华,周小毛
(1.广西大学农学院,广西 南宁 530004;2.湖南省农业生物技术研究中心,湖南 长沙 410125)
丙磺舒在阿维菌素作用棉铃虫细胞中的增效机制研究
王赞永1,周 勇2,马海昊2,刘 佳2,朱 航2,邓希乐2,谭辉华1,周小毛2
(1.广西大学农学院,广西 南宁 530004;2.湖南省农业生物技术研究中心,湖南 长沙 410125)
丙磺舒是ABCC转运蛋白抑制剂,用系列浓度阿维菌素和200 μmol/L丙磺舒的食物饲喂棉铃虫,检测中肠组织中阿维菌素的含量。结果表明:与单独使用阿维菌素相比,混合使用丙磺舒后阿维菌素在中肠中的平均驻留时间(MRT)增加了13%,消除半衰期(t1/2z)增加了27%,药物峰值提高了2.8倍。细胞活力测定发现,亚致死剂量阿维菌素与400 μmol/L丙磺舒混用后,棉铃虫细胞的活力降低了67.1%。表明抑制ABCC转运蛋白可以提高阿维菌素在棉铃虫中肠的驻留能力,提高药物对棉铃虫细胞的毒力。
液相色谱-三重四极杆串联质谱(HPLC-MS);棉铃虫;ABC转运蛋白;阿维菌素;丙磺舒
ABC转运蛋白(ATP—binding cassette transporters)是一个包括A-H 8个亚家族的转运蛋白超家族,可以利用水解ATP的能量将药物泵出细胞外,降低细胞内药物浓度,其功能和表达的改变参与细胞多药耐药的形成[1-2]。医学研究发现,ABC转运蛋白对化疗药物的排出作用是肿瘤细胞产生耐药性的重要原因[3],在病原微生物耐药机制研究中发现,微生物可以通过激活细胞内ABC转运蛋白,将药物运出细胞,降低菌体内抗生素浓度[4-5]。近年的研究表明,ABC转运蛋白基因表达量的提高也和害虫对杀虫剂抗药性增强相关[6]。有报道表明,昆虫对来自9种不同化学类别的27种杀虫剂抗性与ABC转运蛋白的运输作用有关[7],其中涉及到的ABC转运蛋白家族集中在包括ABCC、ABCB和ABCG等亚家族[9-10]。
ABC转运蛋白C家族也被称为多药耐药相关蛋白(Multidrug resistance-associated protein,MRP),在细胞排出胞内有毒物质过程中发挥重要功能。丙磺舒(probenecid)学名对-[(二丙氨基)磺酰基]苯甲酸,是一种MRP的竞争性抑制剂,对MPR1-5的药物外排作用均有不同程度的抑制作用[11]。与抗肿瘤药物联用时,能通过抑制肿瘤细胞的外排作用而提高药效[12],临床上将丙磺舒和抗生素联合使用可以延长抗生素的作用时间[13-14]。
棉铃虫是一种世界范围内的主要害虫,长期以来化学农药的广泛使用使棉铃虫的抗药性增强。现有研究认为,昆虫体内存在多种解毒代谢酶系,如酯酶,多功能氧化酶和谷胱甘肽-S-转移酶等,能够有效的降解进入细胞内的药物分子;另外昆虫体内靶标蛋白的突变和表皮穿透能力的改变也是害虫产生抗药性的主要机制[1,15]。最近的研究表明棉铃虫抗药性的增强与体内ABC转运蛋白C家族的表达量上调有关[16],推测ABCC转运蛋白通过加速排出体内有毒物质增强虫体对农药的耐受性,但目前还缺少直接证据证明ABCC转运蛋白参与该过程。笔者拟通过混合使用ABCC转运蛋白抑制剂丙磺舒和阿维菌素,质谱检测抑制ABCC转运蛋白功能后,研究阿维菌素在中肠组织的滞留情况及对棉铃虫细胞的毒力变化,明确ABCC转运蛋白参与昆虫抗药性的作用机制。
1 材料与方法
1.1 主要仪器与试剂
TSQ Endura高效液相色谱-三重四级杆质谱联用仪(美国赛默飞世尔),JY96-MIN超声细胞破碎仪(宁波新芝),Pico17冷冻高速离心机(美国赛默飞世尔),PQX-350H人工气候箱(中仪国科)Labonova Ultra超纯水系统(美国Think-lab)。BHC-1300生物安全柜(苏州博莱尔),HPS160生化培养箱(北京东联哈尔),DSY-L140倒置显微镜(北京长恒荣创)。
阿维菌素标准品(100 mg/L,北京坛墨质检)阿维菌素原药(96.2%,辉丰农化),丙磺舒原药(99.0%,济南凯恩医药)丙酮、乙腈(色谱纯天津大茂),氯化钠,氯化钾磷酸二氢钾磷酸氢二钠,二甲基亚砜(分析纯,国药集团),甲醇,甲酸(色谱纯德国默克Merck),MTT(Solarbio)。
1.2 供试昆虫及细胞
供试棉铃虫Helicoverpa armigera(Hübner)由中国科学院动物研究所提供,一直使用人工饲料培养,对各种农药敏感。饲养条件:光周期为14 h∶10 h(光照∶黑暗),温度(25±2)℃,相对湿度70%。室内人工饲养采用梁革梅[17]所述方法。
棉铃虫胚胎细胞系QB-Ha-E5[18]由青岛农业大学李国勋教授惠赠,该细胞在27℃、含5% FBS的Gibco SF-900II SFM培养基(美国赛默飞世尔)中培养和传代。
1.3 给药及取样方法
按照农药室内生测标准——人工饲料混药法进行[19],首先将阿维菌素和丙磺舒原药用丙酮溶解,分别配制成1 g/L和10 mmol/L的母液,使用时按比例均匀混入制作好的人工饲料中,终浓度为阿维菌素30 mg/L,丙磺舒200 μmol/L,丙酮1%。对照组食物只加入阿维菌素。
混药饲料切成合适大小放入24孔养虫盒中,放入经过饥饿处理8 h的5龄1 d棉铃虫幼虫。自由取食1 h后更换至洁净养虫盒,用正常人工饲料继续饲喂。每组80头虫,正常进食1 h后开始取样,每间隔1 h取样一次,共设10个取样时间点每次取6头棉铃虫,解剖取出中肠,去除肠道食物残留后,称重,-20℃保存,实验重复3次。
1.4 样品前处理
中肠组织解冻后加入1 mLPBS缓冲液(pH值7.4),于冰水浴中超声破碎;之后取600 μL裂解液,加入1 mL乙腈,0.2 g氯化钠,漩涡震荡1 min,12 000 r/min离心10 min,上清液过0.22 μm微孔滤膜用于HPLC-MS检测。
1.5 色谱条件
色谱柱:Hypersil GOLD C18 (2.1×100 mm 1.9Micron);柱温:35℃;流速:0.3 mL/min;进样量:10 μL;流动相:甲醇-水(含0.1%甲酸)。梯度洗脱程序:0~0.78 min,20%~95%甲醇;0.78~3.1 min,95%甲醇;3.1~4.0 min,95%~20%甲醇。
1.6 质谱条件
ESI离子源,正离子扫描模式,扫描范围m/z500~1 000;离子喷雾电压3 500 V;雾化气为N2,压力40 psi;离子传输管温度300℃,喷针温度300℃;检测方式为MRM模式,定性检测离子m/z 896.4/608.1(碰撞电压为46 V,保留时间为3.72 s),定量检测离子m/z 896.4/752.1(碰撞电压为41 V,保留时间为3.72 s);射频透镜电压为298 V。
质谱数据由TSQ Endura工作站收集并计算出峰面积,计算出样品浓度。按下面公式换算成中肠中阿维菌素含量Cn(μg/g)。

其中C为质谱样品测量浓度,v为萃取液体积,M为中肠样品质量。绘制中肠中阿维菌素含量的浓度-时间曲线,通过DAS 2.0软件对获得的数据进行拟合分析,以统计矩法计算药物动力学参数。
1.7 细胞活力检测
采用MTT细胞染色法,测定阿维菌素对棉铃虫细胞的毒力。分别配制含1、5、10、20、40和60 mg/L阿维菌素的昆虫细胞培养基。处理组均加入终浓度400 μmol/L的丙磺舒,对照组加入等体积溶剂。将对数期棉铃虫细胞以1×104个/孔的密度铺96孔板,27℃培养24 h之后将原培养基吸出替换为不同浓度含药物培养基,每个浓度设6个重复。继续培养48 h进行细胞活力测定,测定方法和数据处理参考周青春[20]研究进行。
2 结果与分析
2.1 质谱条件优化
参考侯志广[21]的阿维菌素检测方法,在流动相中加入甲酸,通过ESI离子源、正离子模式在m/z 500~1 000范围内扫描,得到[M+Na]+准分子离子峰m/z=896.4(阿维菌素分子量为887.11),以此作为母离子,进行二级质谱扫描,得到子离子m/z 752.3和m/z 608.1(图2b),选择m/z 752.3作为定量检测离子,对射频透镜电压和碰撞电压进行优化,得到最优射频透镜电压为298 V,最适碰撞电压为41 V。
2.2 色谱条件优化
选择甲醇-水(0.1%甲酸)作为流动相,阿维菌素的准分子离子峰的离子化效果和稳定性均比较好。在流动相中加入体积分数0.1%的甲酸,能有效消除峰形拖尾现象,并且响应值也大幅提高,稳定出峰时间为3.72 s(图2a)。

图2 阿维菌素质谱图(1 000 μg/L标准品)
2.3 分析方法的评价
2.3.1 标准曲线及检测限 准确配制1 000、500、100、10和1 μg/L系列的阿维菌素标准溶液,按上述样品检测条件测定,以进样浓度为横坐标,定量离子的色谱峰平均峰面积为纵坐标,绘制标准曲线(图3),回归方程为Y=1 809.81+874.513X,R2=0.999 4。在该检测条件下,阿维菌素的检出限为0.1 μg/L(S/N=3)定量限为0.3 μg/L(S/N=10),灵敏度较高,适用于中肠组织中低浓度药物的检测。
2.3.2 添加回收率及方法精密度 用正常棉铃虫中肠制取空白样品,按1 000、100和10 mg/L 3个浓度添加阿维菌素标准品,每个浓度5个重复,并经前处理后连续进样。添加回收率及相对标准偏差(n=5)分别为93.6%(RSD为3.0%)、86.1%(RSD为6.7%)、77.0%(RSD为5.2%)。

图3 阿维菌素标准曲线
2.4 棉铃虫中肠中阿维菌素的药物代谢动力学
棉铃虫饲喂药物后,以正常进食的时间为横坐标,以每克中肠组织中阿维菌素含量为纵坐标,绘制阿维菌素在棉铃虫中肠组织中的代谢曲线(图4),比较不同时间棉铃虫中肠阿维菌素含量差异,发现在棉铃虫中肠组织中1~2 h药物浓度快速升高,2 h时达到峰值(肠组织阿维菌素含量为2.4 μg/g),之后药物浓度快速下降,4 h后下降趋于平缓,6 h后阿维菌素基本完成代谢。混合使用丙磺舒后,棉铃虫中肠对药物总体代谢趋势没有显著变化,但药物峰值提高了2.8倍。根据图2,基于药物代谢动力学研究方法[22],使用DAS 2.0软件计算动力学参数(表2)。结果发现混用丙磺舒之后,药物分子在组织中停留时间(MRT)和消除半衰期(t1/2z)各增加了13%和27%。表明使用ABCC转运蛋白抑制剂后,阿维菌素在棉铃虫中肠内的药物峰值和驻留时间均显著提高,推测其对棉铃虫的毒力增强。
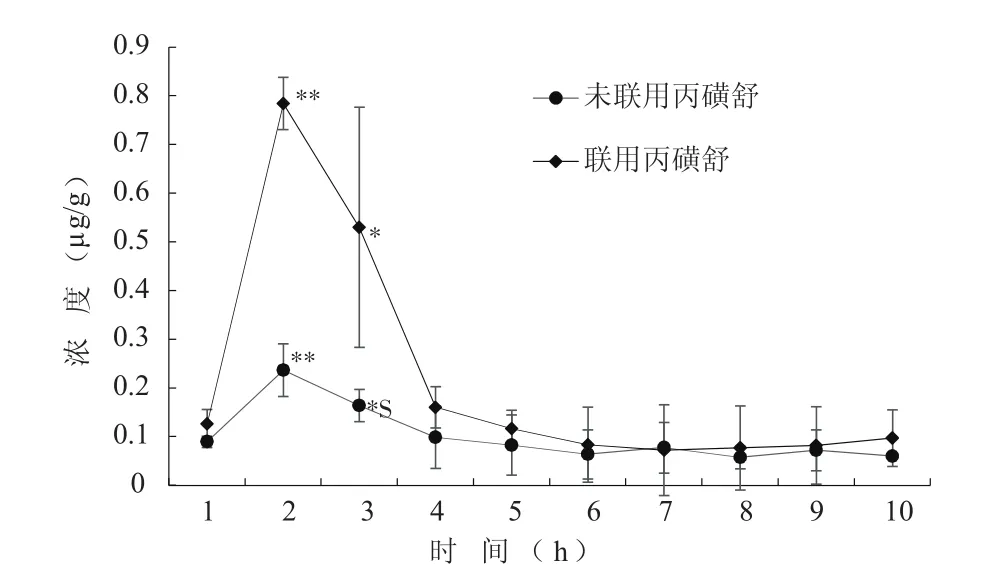
图4 人工饲料混药法给药后棉铃虫中肠中阿维菌素药物浓度-时间曲线
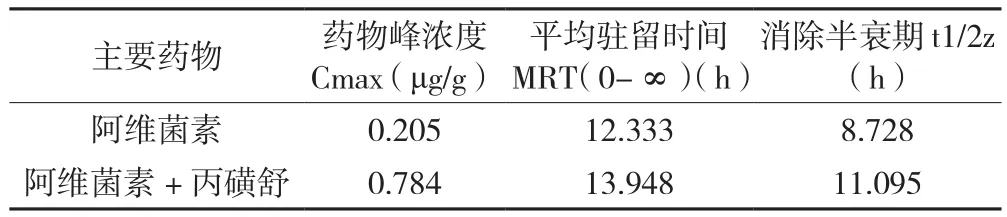
表2 阿维菌素的主要药物代谢动力学参数(统计矩法)
2.5 阿维菌素对棉铃虫的细胞毒力测定
利用不同浓度的阿维菌素处理棉铃虫细胞24 h,MTT法检测细胞活性发现,阿维菌素对棉铃虫胚胎细胞的毒力存在阈剂量效应,当培养基中阿维菌素浓度不超过15 mg/L时细胞活力不受影响,之后随着培养基中阿维菌素药物浓度提高,细胞活力迅速下降。培养基中阿维菌素浓度达到20 mg/L时,细胞活力为23.3%。当培养基中同时添加丙磺舒后,阿维菌素对棉铃虫细胞的毒力存在剂量累积效应,培养基中阿维菌素为1 mg/L时,对细胞没有显著毒力,之后随着药物浓度提高,细胞毒力逐渐增强。EC50为11.971(5.862~18.353)mg/L,毒力方程为y=1.767-1.639lgx (x2=0.156,Sig=0.997)。此时以亚致死剂量阿维菌素处理棉铃虫细胞,发现混合使用丙磺舒后细胞的活力降低了67.1%。表明混用丙磺舒后阿维菌素对棉铃虫细胞的毒力有显著提高。

图5 阿维菌素对棉铃虫细胞毒性MTT染色结果
3 结论与讨论
害虫的抗药性是威胁农业生产安全的重要因素,研究昆虫抗药性作用机制有重要的现实意义。随着研究的深入,昆虫ABC转运蛋白在害虫产生抗药性方面的作用逐渐得到重视,XiaoY等[23]通过RNA干扰抑制ABCC2基因的表达后发现棉铃虫幼虫对阿维菌素的敏感性增加。KM Seong研究发现ABCB49、ABCB50和ABCB65这3个ABC转运蛋白功能异常可以提高果蝇对DTT的抗性[24],禾谷缢管蚜抗药机制研究中发现吡虫啉抗性品系中ABCG20和ABCG23的表达显著高于敏感品系[25]。研究通过混合使用ABCC抑制剂丙磺舒,检测棉铃虫中肠组织对阿维菌素的代谢情况,表明ABCC转运蛋白抑制后,棉铃虫中肠内阿维菌素的药物峰值增加,另外药物在细胞内的驻留时间延长。这和医学研究中通过联用抑制剂能提高血药峰浓度和作用时间的结果是一致的[14,26],充分说明ABCC转运蛋白参与了棉铃虫中肠细胞对阿维菌素的外排作用。
丙磺舒作为ABCC转运蛋白的抑制剂在临床上得到大量应用,与化疗药物联用在人类肺癌,前列腺癌症和乳腺癌治疗中与能极大的提高疗效[12],De Bony报道丙磺舒通过影响肝肾转运,使新一代抗病毒药撷昔洛韦及其代谢物阿昔洛韦的血药峰浓度增加[26]。Leader J P等[27]在研究丙磺舒对昆虫ABC转运蛋白作用机制时采用200 μmol/L,因此笔者也采用200 μmol/L作为棉铃虫中肠的处理浓度,但对昆虫细胞的使用浓度还未有报道。丙磺舒干扰ABCC蛋白的功能,本身对细胞具有一定毒性。Bhaskaracharya A研究发现,丙磺舒对HEK293细胞IC50为203 μmol/L[28],多篇文献基于此浓度对丙磺舒的作用机制进行研究[29-31]。笔者研究比较了丙磺舒对HEK293和棉铃虫细胞毒性的差异,发现2种细胞对丙磺舒耐受能力无显著差异,且基于这一结果,笔者对丙磺舒抑制HEK293和棉铃虫细胞活力进行了检测,结果没有明显差异且200~600 μmol/L的丙磺舒对棉铃虫细胞的毒力无显著差异,因此研究中取400 μmol/L来处理棉铃虫细胞。设置只加入400 μmol/L丙磺舒的孔作为100%活力空白对照,以排除丙磺舒本身毒性的干扰。实验结果表明丙磺舒可以提高药物分子在棉铃虫细胞内的驻留能力,提高药物毒力,有显著的增效作用。丙磺舒或可作为一种增效剂与杀虫剂混合使用以提高药效,降低农药使用量,但其在昆虫上使用的效果,以及环境安全性,还有待进一步研究。
总之,研究通过抑制剂阻断研究,表明ABCC转运蛋白家族参与昆虫细胞对有毒物质的排出作用,同时证明了丙磺舒在和阿维菌素混用时的增效作用,并阐明了增效机制,为基于该作用机制开发新型增效剂提供了条件。
[1] Linda J G,Yannick P,Heiko V,et al. An ABC transporter mutation is correlated with insect resistance to Bacillus thuringiensis Cry1Ac toxin [J]. Plos Genetics,2010,6(12):1-11.
[2] Labbé R,Caveney S,Donly C. Genetic analysis of the xenobiotic resistance-associated ABC gene subfamilies of the Lepidoptera [J]. Insect Molecular Biology,2011,20(2):243.
[3] Ikeda K,Oka M,Yamada Y,et al. Adult T-cell leukemia cells overexpress the multidrug-resistance-protein (MRP) and lung-resistanceprotein (LRP) genes [J]. International Journal of Cancer,1999,82(4):599.
[4] 张 婵,庄肃军,杨丽红,等. 氟康唑体外诱导白假丝酵母菌耐药基因表达的研究[J]. 中国微生态学杂志,2013,25(8):911-913.
[5] Reyes C L,Ward A,Yu J,et al. The structures of MsbA: Insight into ABC transporter-mediated multidrug efflux [J]. FEBS Lett,2006,(580):1042-1048.
[6] Qi W,Ma X,He W,et al. Characterization and expression profiling of ATP-binding cassette transporter genes in the diamondback moth, Plutella xylostella (L.) [J]. Bmc Genomics,2016,17(1):760.
[7] 朱 航,王雅菲,周小毛. 小菜蛾ABCG2mRNA表达特征及茚虫威对其表达量的影响[J]. 植物保护,2015,(6):117-121.
[8] Dermauw W,Van L T. The ABC gene family in arthropods comparative genomics and role in insecticide transport and resistance [J]. Insect Biochemistry & Molecular Biology,2014,45(1):89.
[9] Dermauw W,Osborne E J,Clark R M,et al. A burst of ABC genes in the genome of the polyphagous spider mite Tetranychus urticae [J]. BMC Genomics,2013,14(1):317.
[10] Franco J,Ferreira R C,Ienne S,et al. ABCG-like transporter of Trypanosoma cruzi involved in benznidazole resistance:gene polymorphisms disclose inter-strain intragenic recombination in hybrid isolates [J]. Infection Genetics & Evolution,2015,(31):198-208.
[11] 夏春华,王广基. 丙磺舒对多药耐药相关蛋白的逆转作用及其对药代动力学的影响与应用[J]. 中国药科大学学报,2007,38(5):477-480.
[12] Sirotnak F M,Wendel H G,Bornmann W G B,et al. Coadministration of Probenecid,an Inhibitor of a cMOAT/MRPlike Plasma Membrane ATPase,Greatly Enhanced the Efficacy of a New 10-Deazaaminopterin against Human Solid Tumors in Vivo1 [J]. Clinical Cancer Research,2000,6(9):3705-3712.
[13] Newell A,Herbert E,Vigus J,et al. Gonorrhoea in a south London genitourinary medicine department [J]. International Journal of Std & Aids,2003,14(9):625-629.
[14] Spina S P,D E Jr. Effect of chronic probenecid therapy on cefazolin serum concentrations [J]. Annals of Pharmacotherapy,2003,37(5):621-624.
[15] 刘永杰,沈晋良,杨田堂,等. 氯氟氰菊酯对斜纹夜蛾抗性和敏感种群表皮穿透比较[J]. 中国农业科学,2009,42(7):2386-2391.
[16] Bretschneider A,Heckel D G,Vogel H. Know your ABCs:Characterization and gene expression dynamics of ABC transporters in the polyphagous herbivore Helicoverpa armigera [J]. Insect Biochemistry &Molecular Biology,2016,(72):1-9.
[17] 梁革梅,谭维佳,郭予元. 人工饲养棉铃虫技术的改进[J]. 植物保护,1999,25(2):15-17.
[18] 郑桂玲,李长友,周洪旭,等. 两株棉铃虫胚胎新细胞系的建立及其对杆状病毒侵染的反应[J]. 昆虫学报,2010,(53):167-174.
[19] NY/T 1154.10-2008,农药室内生物测定准则杀虫剂第10部分:人工饲料混药法[S].
[20] 周青春,洪华珠. 利用MTT法测定杀虫剂对细胞的毒力[J]. 植物保护学报,1996,23(4):343-348.
[21] 侯志广,方永睿,嘧东林,等. 高效液相色谱-三重串联质谱法同时检测稻田中阿维菌素与茚虫威的残留量[J]. 农药,2015,(10):744-747.
[22] 邢晓玲,王妲妲,袁小秋,等. 甲砜霉素及HP-β-CD甲砜霉素在家兔体内的药代动力学比较研究[J]. 湖南农业科学,2009,(12):11-14.
[23] Xiao Y,Liu K,Zhang D,et al. Resistance to Bacillus thuringiensis Mediated by an ABC Transporter Mutation Increases Susceptibility to Toxins from other Bacteria in an Invasive Insect [J]. Plos Pathogens,2016,12(2):1-20.
[24] Seong K M,Sun W,Clark J M,et al. Splice form variant and amino acid changes in MDR49 confers DDT resistance in transgenic Drosophila [J]. Sci Rep,2016,(6):23355.
[25] 康新乐,李玉婷,王 康,等. 禾谷缢管蚜ABC转运蛋白基因的克隆、分子特性及表达分析[J]. 昆虫学报,2015,58(6):593-602.
[26] De B F,Tod M,Bidault R,et al. Multiple interactions of cimetidine and probenecid with valaciclovir and its metabolite acyclovir [J]. Antimicrob Agents Chemother,2002,46(2):458.
[27] Leader J P,O'Donnell M. J. Transepithelial transport of fluorescent p-glycoprotein and MRP2 substrates by insect Malpighian tubules:confocal microscopic analysis of secreted fluid droplets [J]. Journal of Experimental Biology,2005,(208):4363-4376.
[28] Bhaskaracharya A,Daoung P,Jalilian I,et al. Probenecid Blocks Human P2X7 Receptor-Induced Dye Uptake via a Pannexin-1 Independent Mechanism [J]. Plos One,2014,9(3):1-8.
[29] Watanabe I,Tatebe J,Namba S,et al. Activation of aryl hydrocarbon receptor mediates indoxyl sulfate-induced monocyte chemoattractant protein-1 expression in human umbilical vein endothelial cells [J]. Circulation Journal Official Journal of the Japanese Circulation Society,2013,77(1):224-230.
[30] Nzila A,Mberu E,Bray P,et al. Chemosensitization of Plasmodium falciparum by Probenecid In Vitro [J]. Antimicrobial Agents & Chemotherapy,2003,47(7):2108.
[31] Yu C P,Sweet D H,Peng Y H,et al. Effects of nonsteroidal antiinflammatory drugs on the renal excretion of indoxyl sulfate,a nephrocardiovascular toxin,in rats [J]. European Journal of Pharmaceutical Sciences,2017,(101):66-70.
(责任编辑:肖彦资)
TheSynergismMechanismofProbenecidonAvermectinsTreatingHelicoverpaArmigeraCell
WANG Zan-yong1,ZHOU Yong2,Ma Hai-hao2,LIU Jia2,ZHU Hang2,DENG Xi-le2,TAN Hui-hua1,ZHOU Xiao-mao2
(1. College of Agriculture, Guangxi Univeristy, Nanning 530004,PRC; 2. Hunan Agricultural Biotechnology Research Center, Changsha 410125, PRC)
Probenecid is inhibitor of ABC transport. In this research, we use artificial feed contain 30 mg/L avermectins and 200 μmol/ L probenecid to feed Helicoverpa armigera and detect the content of avermectins in Helicoverpa armigera midgut. The results showed that compared with the using avermectin alone, after mixed with probenecid, the average dwell time (MRT) of avermectins in midgut increased by 13%, the Half-life (t1/2 z) increased by 27%, and Peak concentration increased 2.8 times. It was found that after using combination of sublethal dose avermectins and 400 μmol/L probenecid, Ha cell viability reduced by 67.1% by cell vitality determination. These phenomenon indicated that inhibition of ABC transporters can improve intestinal resident ability of avermectins in the Ha midgut, improve drug toxicity to the cells of Ha.
liquid chromatography-triple quadrupole tandem mass spectrometry(HPLC-MS); Helicoverpa armigera; ABC transporters; avermectins; probenecid
S482
A
1006-060X(2017)05-0064-05
10.16498/j.cnki.hnnykx.2017.005.018
2017-03-12
国家自然科学基金(31371966)
王赞永(1987-),男,湖南长沙市人,硕士研究生,研究方向为农药毒理与有害生物抗药性。
谭辉华,周小毛
