甘蓝型油菜花粉母细胞减数分裂行为观察
2017-06-21李东昊
李东昊,姜 玲,阮 颖
(湖南农业大学生物科学技术学院,湖南 长沙 410128)
甘蓝型油菜花粉母细胞减数分裂行为观察
李东昊,姜 玲,阮 颖
(湖南农业大学生物科学技术学院,湖南 长沙 410128)
以甘蓝型油菜“湘油15号”为材料,采用改良的卡宝品红染色液对其进行花粉母细胞减数分裂过程的细胞学观察。结果表明:当甘蓝型油菜花蕾长度小于1.5 mm时,花粉母细胞大多处于减数分裂时期;而花蕾长度介于1.5~2.5 mm时,花粉母细胞大多已发育为单核花粉粒;相较于碱性品红和Gimesa染色液,改良的卡宝品红染色液着色效果更佳,核质区分明显;甘蓝型油菜花粉母细胞在经历减数分裂Ⅰ和减数分裂Ⅱ后形成四分体,期间存在两次胞质分裂。
甘蓝型油菜;花粉母细胞;减数分裂
减数分裂是有性生殖的个体形成雌雄配子的一种特殊分裂方式[1],对遗传变异、生物进化等有着重要的生物学意义[2-3]。同时,它也是单倍体、多倍体遗传育种[4-5]以及辐射诱变育种和败育性研究[6]的理论基础。
甘蓝型油菜(Brassica napus L.)系十字花科芸薹属植物,是由二倍体的白菜型油菜(Brassia campestris L.)和甘蓝(Brassica oleracea L.)自然杂交而成的异源四倍体[7],在长江中下游流域广为栽培,是非常重要的油料作物[8-9]。油菜减数分裂过程中雌雄配子发育的正常与否直接关系到产量的高低[10]。因此,笔者通过改良的卡宝品红染色法对甘蓝型油菜花粉母细胞减数分裂过程进行了较为完整的细胞学观察,以期为研究甘蓝型油菜减数分裂过程中染色体上组蛋白甲基化修饰的变化情况打下基础。
1 材料与方法
1.1 试验材料
供试甘蓝型油菜品种为“湘油15号”,由湖南农业大学植物发育与表观遗传调控实验室提供。
1.2 试验方法
1.2.1 花蕾长度与小孢子母细胞发育情况观察 油菜现蕾后,分别选取花蕾长7.0、6.5、6.0、5.5、5.0、4.5、4.0、3.0、2.5、2.0、1.5、1.0和小于1.0 mm的花进行染色和细胞学观察。
1.2.2 改良品红染色液制备 首先,制备原液A,将3 g碱性品红溶于100 mL 70%酒精。然后,取原液A 10 mL加入到90 mL 5%石炭酸水溶液中制成原液B。接着,取原液B 55 mL,加入6 mL冰醋酸和6 mL福尔马林(38%的甲醛)制成原液C。染色液工作液配制:取原液C 10 mL,加入80 mL 45%冰醋酸,再加入1.8 g山梨醇配成10%浓度的卡宝品红染色液,静置14 d,备用。
1.2.3 花粉母细胞减数分裂过程的观察 油菜现蕾开花后,采集处于不同发育时期的花蕾;将采集的材料用卡诺固定液(酒精︰冰醋酸=3︰1)固定24 h,使用1 mol/L盐酸60℃解离5 min;分别用Gimesa染色液、传统的碱性品红染色液和改良的卡宝品红染色液进行染色(30 min),在显微镜下观察并拍摄,比较3种染色液的着色效果。
2 结果与分析
2.1 花蕾长度与雄配子体发育时期的关系
从图1中可以看出,当花蕾长度为1.5~2.5 mm时,约80%的花粉母细胞已发育为单胞花粉粒;而花蕾小于1.5 mm时,85%的花粉母细胞正处于减数分裂时期。因此,试验选取花蕾小于1.5 mm的花粉母细胞进行细胞学观察。
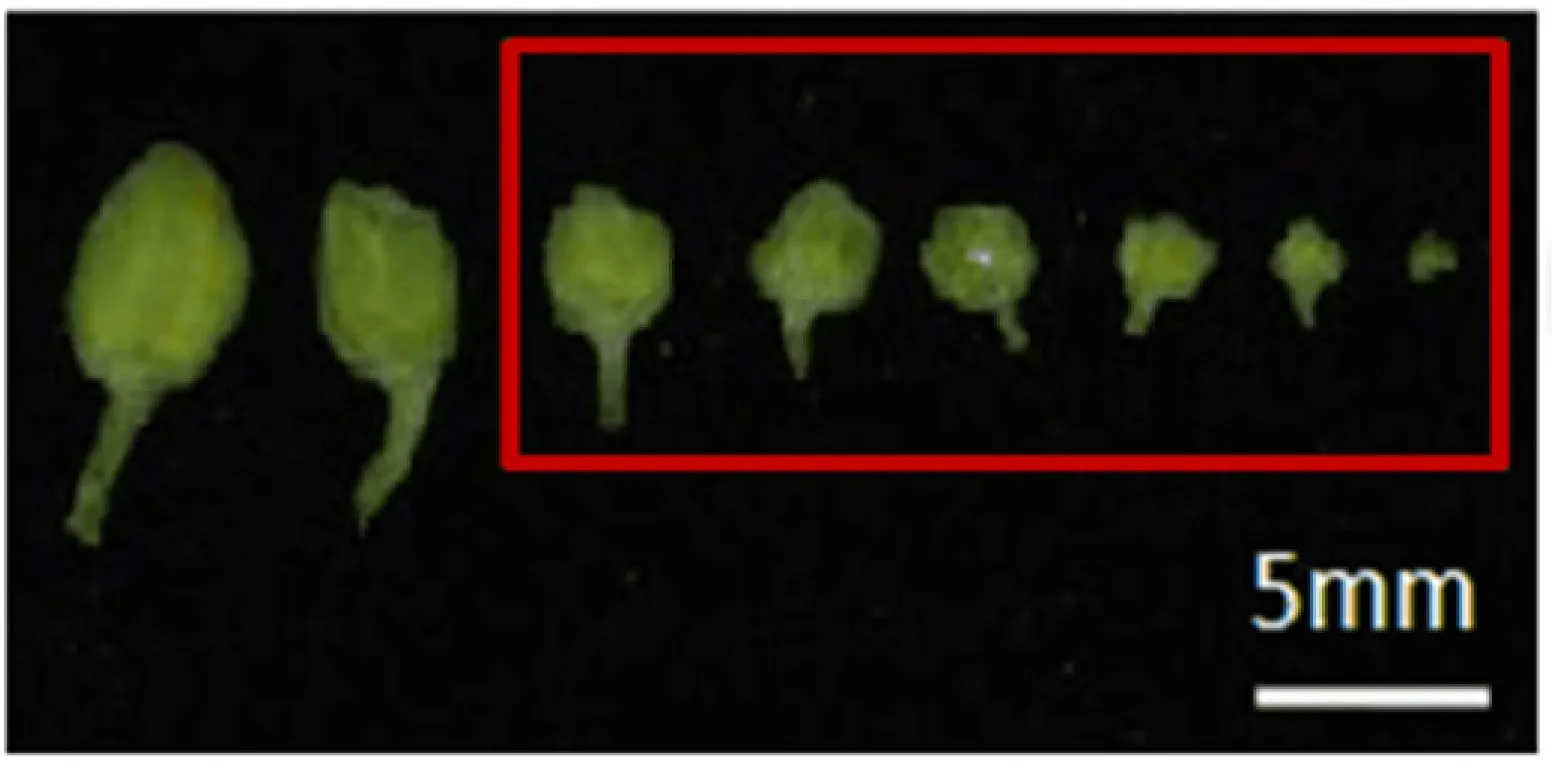
图1 3种不同染色液不同长度的花蕾
2.2 3种不同染色液着色效果的比较
如图2所示,Gimesa染色液着色不明显,而传统的碱性品红染色液,尽管其着色较深,可以很好地观察到染色质位置,但是缺点也很明显,即在一些时期,不能很好地区分染色体状态;相较于前两者,改良后的卡宝品红染色液可以较好地着色,以便清晰地观察花粉粒形态和染色体行为。
2.3 甘蓝型油菜花粉母细胞减数分裂过程观察
对小于1.5 mm的花蕾进行染色和显微观察发现,跟大多数植物一样,甘蓝型油菜“湘油15号”的花粉母细胞历经了分裂间期、减数分裂Ⅰ、减数分裂Ⅱ以及2次胞质分裂后,最终形成四分体。
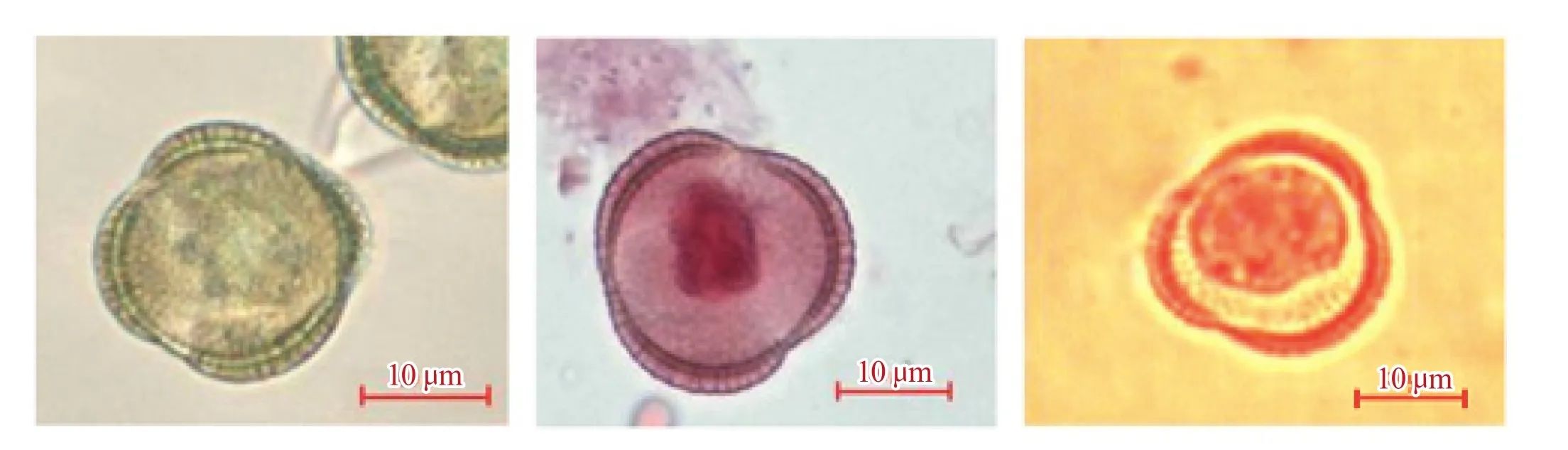
图2 3种不同染色液着色效果的比较
花粉母细胞首先进入减数分裂间期,此时细胞形态多为近似圆形。进入减Ⅰ前期:细胞核开始消失,染色质加深浓缩形成染色体,此时可以较为清楚地看到联会的同源染色体(图3A)。减Ⅰ中期:细胞核膜与核仁消失,纺锤体出现,染色体集中出现在细胞质中间的赤道板上,此时染色体排布有重叠现象不利于染色体的观察(图3B)。减Ⅰ后期:核内同源染色体由纺锤丝牵引,各自移向两极,此时可以观察到核内两侧有染色体的聚集(图3C)。减Ⅰ末期:染色体解压缩,染色质逐渐向中间聚集,细胞质分裂1次,形成2个大小均等的细胞质(图3D)。二分体时期:减数第一次分裂后,核内染色质不进行复制,经短暂的间期后,分裂在2个子细胞的染色质再次浓缩形成染色体(图3E)。减Ⅱ中期:各姐妹染色单体向细胞中部聚集,排布于赤道板上(图3F)。减Ⅱ后期:姐妹染色单体由纺锤体牵引向两侧聚集(图3G)。四分体时期:两侧聚集的染色体再次解浓缩,核膜核仁出现,细胞质进行第二次分裂,小孢子细胞质外形成胼胝质共同包围形成四分体,至此减数分裂完成(图3H)。
此后小孢子进一步发育直至形成具有细胞外壁的单核花粉粒。游离状态下的单核花粉粒,形状近似椭圆,细胞核大且集中,胞质浓厚,未观察到明显的液泡。
3 讨 论
甘蓝型油菜,尤其是在自然界稳定遗传的甘蓝型油菜[11]。在新合成的双二倍体中,稳定的染色体配对通常是由遗传控制(例如小麦、燕麦)的,或者是由染色体结构改变而获得的(如棉花)[12-15]。尽管碱性品红染色液染色效果出众,但是对染色体与细胞质的区分效果较差。而改良后的卡宝品红染色液能有效地解决这一问题,使染色体着色明显强于细胞质,在观察时很容易将两者区分开来。
表观遗传学(epigenetics)是不同于传统遗传学的一个学科分支,其研究的主要内容是由非基因序列改变而引起的,在发育和细胞增殖过程中可以稳定传递的基因或者蛋白表达的变化[16-17]。染色体上组蛋白发生的共价化学修饰(甲基化,乙酰化等)也属于表观遗传学的范畴[18]。因此,观察花粉母细胞的减数分裂行为,有助于开展甘蓝型油菜的表观遗传学研究。试验结果显示,在减数分裂Ⅰ和Ⅱ的前期、中期和后期可以很好地观察到染色体的分布情况,为研究甘蓝型油菜染色体上组蛋白甲基化修饰的变化情况打下了基础。

图3 “湘油15号”花粉母细胞减数分裂过程
[1] 李再云,刘后利. 甘蓝型油菜减数分裂中的染色体行为观察[J]. 华中农业大学学报,1994,13(4):418-421.
[2] 吕 琳,何聪芬,董迎卯,等. 木立芦荟小孢子母细胞减数分裂与花粉育性关系的初步研究[J]. 遗传,2005,27(3):429-434.
[3] 冯九焕,卢永根,刘向东,等. 水稻花粉发育过程及其分期[J]. 中国水稻科学,2001,15(1):21-28.
[4] 张晓伟,高睦枪,原玉香,等. 人工合成甘蓝型油菜研究[J]. 河南农业科学,2001,(2):7-10.
[5] 钱 伟. 甘蓝型油菜与白菜型油菜亚基因组问杂种优势的研究[D].武汉:华中农业大学,2003.
[6] 叶纨芝,曹家树,余小林,等. 白菜细胞核雄性不育两用系的细胞学观察[J]. 细胞生物学杂志,2004,26:516-522.
[7] 傅廷栋. 杂交油菜的育种与利用[M]. 武汉:湖北科技出版社,2000.
[8] 李培武,赵永国,张 文,等. 中国甘蓝型油菜硫苷含量及组份分析[J]. 中国农业科学,2005,38(7):1346-1352.
[9] Liu Z Q,Adamczyk K,Manzanarex-dauleux M,et al. Mapping PrBn and other quantitative trait loci responsible for the control of homeologous chromosome pairing in oilseed rape(Brassica napus L.)haploids[J]. Genetics,2006,174:1583-1596.
[10] 余凤群,傅廷栋. 甘蓝型油菜几个雄性不育系的细胞形态学研究[J].武汉植物学研究,1990,8(3):119-216.
[11] 杨光圣,瞿 波,傅廷栋. 三个甘蓝型油菜隐性细胞核雄性不育系小孢子发生的细胞学研究[J]. 华中农业大学学报,1999,18(5):520-523.
[12] Rajhathy T,Thomas H. Genetic control of chromosome pairing in hexaploid oats[J]. Nature New Biology,1972,239:217-219.
[13] Mursal I E. Endrizzi J E. A reexamintiton of the diploid like meiotic behavior of polyploid cotton[J]. Theor Appl Genet,1976,47:171-178.
[14] Jenczewski E,Eber F,Grimaud A,et al. A major gene controlling homeologous pairing in oilseed rape(Brassica napus)haploids[J]. Genetics,2003,164:645-653.
[15] 李 俊,方小平. 人工合成甘蓝型油菜减数分裂中的染色体行为观察[J]. 江西农业学报,2010,22(1):1-4.
[16] 董玉玮,侯进慧,朱必才,等. 表观遗传学的相关概念和研究进展[J].生物学杂志,2005,22(1):1-3.
[17] 李光雷,喻树迅,范术丽. 表观遗传学研究进展[J]. 生物技术通报,2011,1(1):41-49.
[18] 于 红. 表观遗传学:生物细胞非编码RNA调控的研究进展[J].遗传,2009,31(11):1077-1086.
(责任编辑:成 平)
ObservationofPollenMotherCellMeiosisinBrassica napus
LI Dong-hao,JIANG Ling,RUAN Ying
(College of Biological Science and Technology, Hunan Agricultural University, Changsha 410128, PRC)
The cytoplasmic observation of pollen mother cell meiosis in Brassica napus Xiangyou 15 was carried out with the modified carbol fuchsin. The results indicated that when the length of buds is less than 1.5 mm, pollen mother cells mostly are in the meiosis period; and while the length of buds are between 1.5 mm and 2.5 mm, pollen mother cells mostly develop into mononuclear pollen grains. Compared with alkaline fuchsin and Gimesa, the modified carbol fuchusin is effective in pigmentation, with clear nucleus-cytoplasm distinction. And after the meiosisⅠand meiosisⅡ, pollen mother cells of Brassica napus develop into tetrads, along with cytokinesis twice.
Brassica napus; polen mother cell; meiosis
Q942.4
A
1006-060X(2017)05-0001-03
10.16498/j.cnki.hnnykx.2017.005.001
2017-03-05
作物种质创新与资源利用国家重点实验室科学基金开放项目(12KFXM10)
李东昊(1992-),男,湖南长沙市人,硕士研究生,专业方向:植物发育与表观遗传。
阮 颖
