中国荷斯坦奶牛FASN基因部分序列多态性分析
2017-06-09任卫合王丽萍刘容秀
任卫合,王丽萍,刘容秀,张 丽
(西北民族大学生命科学与工程学院,兰州 730030)
研究简报
中国荷斯坦奶牛FASN基因部分序列多态性分析
任卫合,王丽萍,刘容秀,张 丽
(西北民族大学生命科学与工程学院,兰州 730030)
利用PCR-SSCP技术,对中国荷斯坦奶牛脂肪酸合成酶(fatty acid synthase,FASN)基因第34外显子进行多态性分析。结果显示:该区域存在A、B两种等位基因和AA、BB、AB三种基因型,其中等位基因A和B的频率分别为0.890 8和0.109 2,基因型AA、BB和AB的频率分别为0.908 0、0.011 5和0.195 4。A为优势等位基因,AA为优势基因型。群体遗传学分析发现,荷斯坦奶牛FASN基因的有效等位基因数(Ne)、遗传杂合度(He)、遗传纯合度(Ho)和多态信息含量(PIC)分别为0.184 5、0.195 7、0.804 3和0.175 6。卡方检验表明,该群体已经极显著地偏离了Hardy-Weinberg平衡状态(P<0.01)。说明中国荷斯坦奶牛FASN基因exon34区存在遗传多态性。
FASN基因;基因多态性;PCR-SSCP;中国荷斯坦奶牛
脂肪酸合成酶(fatty acid synthase,FASN)是由FASN基因编码的一种基本代谢酶类,对控制动物体脂沉积和乳脂脂肪酸合成代谢具有重要意义[1]。该酶主要表达在哺乳动物的肝脏和脂肪组织中[2-3],负责催化乙酰辅酶A和丙二酸单酰辅酶A合成一种长链饱和软脂酸[4]。研究发现,脂肪酸合成酶直接受FASN基因表达情况的影响,若FASN基因的碱基序列发生突变,牛全乳中的脂肪酸含量就可能发生变化,对牛的泌乳性能产生影响[5]。Roy等[6]将牛FASN基因定位于19号染色体上,发现其包含42个外显子,编码2 513个氨基酸残基,且含有多个SNPs。研究表明,FASN基因缺失的小鼠胚胎会在其胚胎发育早期死亡,FASN基因突变的小鼠则会在其发育的不同时期死亡,表明脂肪酸合成酶对胚胎发育也有重要作用[7]。褚敏等[8]研究推测,FASN基因不仅影响牛肉脂肪酸的组成及含量,亦对牦牛肉的脂肪量及食用品质有影响,可作为牦牛肉质性状标记辅助选择的有效分析标记。
目前国内对荷斯坦奶牛FASN基因的研究还很少,本研究利用PCR-SSCP技术研究中国荷斯坦奶牛FASN基因第34外显子多态性,旨在丰富荷斯坦奶牛FASN基因库,并为泌乳基因、胚胎发育基因和肉质性状基因的筛选提供分子理论基础。
1 材料与方法
1.1 样品采集及基因组DNA提取
血样采自宁夏贺兰山奶业有限公司的87头中国荷斯坦奶牛,用医用采血管静脉采血,每头10 mL,-20℃保存。采用苯酚-氯仿抽提法从备用冷冻血样中提取总基因组DNA,去离子水溶解稀释,-20℃保存备用。
1.2 引物设计与PCR扩增
根据GenBank中牛FASN基因的mRNA序列(登录号AF285607)设计1对特异性引物,用于扩增荷斯坦奶牛FASN基因序列。引物序列Forward:5′-CCAGACCTTAATTTGCCAATCCTC-3′,Reverse:3′CCCTCTAAAGCCGTCCTCACCA-5′,引物由大连宝生物公司合成,扩增区域从15 906 bp开始,包括了FASN基因外显子34的全部序列,目的片段216 bp。
以提取的DNA为模板,PCR扩增FASN基因。PCR扩增总反应体系为20 μL,其中模板DNA 0.8 μL,上下游引物各0.4 μL,Taq PCR MasterMix 11 μL,ddH2O 7.4 μL。PCR反应程序:94℃预变性4 min;94℃变性30 s,54℃退火30s,72℃延伸30 s,35个循环;最后72℃延伸10 min。PCR产物经1%琼脂糖凝胶电泳检测后置于4℃冷藏保存,以备SSCP分析。
1.3 SSCP检测与等位基因序列的回收测定
取3.0 μL的PCR产物与7.0 μL变性上样缓冲液(0.025%溴酚蓝、98%去离子甲酰胺、10 mmol/L EDTA、0.025%二甲苯氰)混合,98℃变性10 min后再立即冰浴10 min。将变性后的PCR产物点样至12%的非变性聚丙烯酰胺凝胶上(Acr∶Scr=29∶1),160V电压室温电泳4h。银染法显带,并确定其基因型。PCR产物经过SSCP确定基因型后,纯合子PCR产物用试剂盒回收后直接送北京华大公司测序。而杂合子个体参照Hu等[9]介绍的方法进行处理。
1.4 统计分析
利用Popgene32软件计算出FASN基因不同突变位点的等位基因频率、基因型频率、有效等位基因数(Ne)、遗传杂合度(He)、遗传纯合度(Ho)和多态信息含量(PIC),并对该群体进行Hardy-Weinberg平衡检验。
2 结果与分析
2.1 荷斯坦奶牛FASN基因PCR扩增结果
对303头荷斯坦奶牛的DNA样品PCR扩增后,经1%琼脂糖凝胶(100 V,30 min)电泳检测后均得到一条如图1所示的特异性好、条带清晰、长度约为216 bp的产物,与预期的目的片段大小一致,可用于SSCP检测。

图1 荷斯坦奶牛FASN基因exon34区域PCR扩增结果
2.2 PCR-SSCP检测及序列测定结果
中国荷斯坦奶牛FASN基因exon34序列区域共检测到A和B两种等位基因,共形成AA、BB、AB三种基因型(图2),其中等位基因A和B均具纯合型个体。

图2 荷斯坦奶牛FASN基因exon34区域SSCP检测结果
2.3 遗传参数分析
荷斯坦奶牛FASN基因exon34区基因频率、基因型频率以及各遗传多态性指标见表1和表2。等位基因A和B的频率分别为0.890 8和0.109 2,AA、BB和AB基因型频率分别为0.908 0、0.195 4和0.011 5。本研究中,荷斯坦奶牛FASN基因的有效等位基因数(Ne)、遗传杂合度(He)、遗传纯合度(Ho)和多态信息含量(PIC)分别为0.1845、0.1957、0.8043和0.1756。PIC值小于0.25,属于低度多态,说明该位点较为保守。卡方检验表明该群体已经极显著地偏离了Hardy-Weinberg平衡状态(P<0.01)。
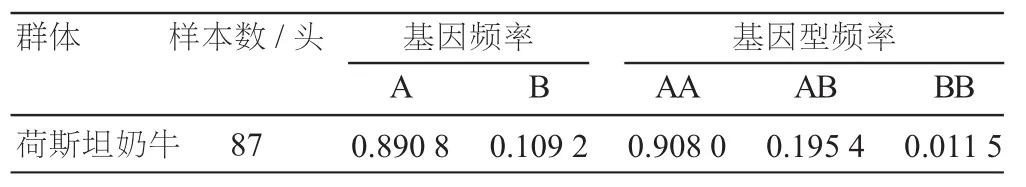
表1 荷斯坦奶牛FASN基因exon34区基因频率和基因型频率
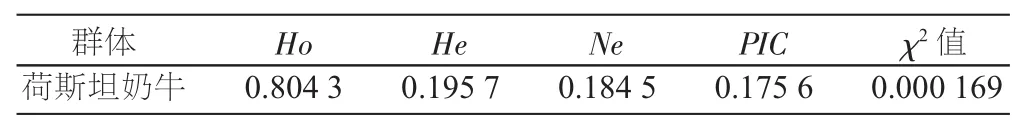
表2 遗传多态性分析
3 讨论
近年来,国内外有关FASN基因的多态性报道相对较少。Abe等[10]在日本黑牛FASN基因编码区发现8个SNPs。Chirala等[7]在牛FASN基因中检测到了5个多态位点。褚敏等[8]在甘南牦牛、天祝白牦牛和大通牦牛FASN基因的第3内含子检测到1个多态位点。欧阳建华等[11]在鸡FASN基因最后1个内含子发现2个多态位点。Abiel[12]在西农萨能奶山羊的FASN基因中检测到3个多态位点。
本研究通过对荷斯坦牛FASN基因exon34区进行PCR-SSCP分析,结果发现A、B两个等位基因,其基因频率分别为0.890 8、0.109 2,A为优势基因,试验群体已经极显著地偏离了Hardy-Weinberg平衡状态(P<0.01),说明该位点受到漂变、选择和引种等因素的影响。这与Roy等[6]和Chirala等[7]的研究结果存在一定的差异;而与马彦南等[5]的研究结果较为一致。造成这种结果的原因可能是试验群体的环境和种群存在差异,也可能是中国荷斯坦牛含有地方黄牛品种血统。本研究在试验群体中仅发现一个个体为BB型,这可能是由于样本量限制的缘故。在遗传多态性分析中,等位基因数(Ne)、多态信息含量(PIC)和杂合度(He)都是群体内遗传变异的测度,其值的高低反映了群体遗传变异的大小,遗传变异大,数值越高。本研究试验群体PIC值小于0.25,呈低度多态,这与武秀香等[2]的研究结果存在一定差异,可能是由于A等位基因对荷斯坦奶牛的生长发育、泌乳性能有正效应,所以在长期的人工育种过程中受到正向选择、处于优势地位的缘故。
[1] 杨永新,王加启,卜登攀,等.牛乳重要营养品质特征的研究进展[J].食品科学,2013,34(1):328-332.
[2] 武秀香,杨章平,王小龙,等.中国荷斯坦奶牛FASN基因遗传多样性及其与泌乳性能的相关性分析[J].中国畜牧杂志,2010,46(1):1-4.
[3] 颜新春,汪以真,许梓荣.动物脂肪酸合成酶(FAS)基因表达的调控[J].动物营养学报,2002,14(2):1-4.
[4] Menendez J A,Lupu R.Fatty acid synthase-catalyzed de novo fatty acid biosynthesis:from anabolic-energy-storage pathway in normal tissues to jack-of-all-trades in cancer cells[J].Archivum Immunologiae Et Therapiae Experimentalis,2004,52(6):414.
[5] 马彦男,贺鹏迦,马永生,等.中国荷斯坦牛FASN基因部分序列PCR-SSCP多态性与泌乳性状的相关性[J].西北农林科技大学学报(自然科学版),2013,41(12):1-6.
[6] Roy R,Gautier M,Hayes H,et al.Assignment1 of the fatty acid synthase(FASN)gene to bovine chromosome 19(19q22)by in situ hybridization and confirmation by somatic cell hybrid mapping[J].Cytogenetics&Cell Genetics,2001,93(1-2):141.
[7] Chirala S S,Chang H,Matzuk M,et al.Fatty acid synthesis is essential in embryonic development:fatty acid synthase null mutants and most ofthe heterozygotes die in utero[J].Proc Natl Acad Sci USA,2003,100(11):6358-6363.
[8] 褚敏,焦斐,吴晓云,等.牦牛FASN基因多态性及其与肉质性状的相关性研究[J].中国草食动物科学,2014,34(1):17-20.
[9] Hu J,Zhou H,Smyth A,et al.Polymorphism of the bovine ADRB3 gene[J].Molecular BiologyReports,2010,37(7):3389-3392.
[10] Abe T,Saburi J,Hasebe H,et al.Novel mutations of the F SNA gene and their effect on fatty acid composition in Japanese Black beef[J]. Biochemical Genetics,2009,47(5):397-411.
[11]欧阳建华,谢亮,刘杰,等.鸡FASN基因2个位点的多样性及其与体重、脂肪沉积性状的相关性[J].畜牧兽医学报,2007,38(1):25-30.
[12]Abiel Berhane Haile.西农萨能奶山羊脂肪酸合酶(FASN)和硬脂酰CoA去饱和酶1(SCD1)基因多态性与乳脂肪酸成分的关联分析[D].陕西杨凌:西北农林科技大学,2015.
S823.2
A
2095-3887(2017)03-0067-03
10.3969/j.issn.2095-3887.2017.03.018
2017-04-13
任卫合(1997-),男,在读本科生。
张丽(1980-),女,副教授,博士。研究方向:牛羊遗传育种。
