苹果植株应答连作障碍差异蛋白质组学分析
2017-06-05倪伟倪蔚茹赵静王增辉毛志泉陈学森沈向
倪伟 倪蔚茹 赵静 王增辉 毛志泉+陈学森+沈向


摘要: 為明确不同苹果品种应对连作障碍时差异表达的蛋白,在蛋白表达水平上揭示苹果应答连作障碍的机制,本研究选择在重茬地中表现不同的两个苹果品种(抗重茬能力强的富士2001和抗重茬能力弱的首富1号)作为试材,利用重茬土进行盆栽试验,运用蛋白质非标记定量技术(label-free)鉴定两个苹果品种叶片中差异表达的蛋白,将差异蛋白与 GO、COG等数据库进行比对,分析不同苹果品种差异蛋白的功能和分类。结果表明,两个苹果品种在重茬土栽植条件下主要鉴定得到43种差异或特异表达的蛋白,主要分为4类,包括光合作用相关蛋白(1种)、植物能量及物质代谢相关蛋白(17种)、植物防御和抗逆性相关蛋白(8种)、其他功能蛋白(17种),其中,植物能量及物质代谢、植物防御和抗逆性相关蛋白表达量具有较大差异。不同苹果品种在连作障碍反应相关蛋白的表达量方面存在差异,抗重茬能力强的品种蛋白表达量明显高于抗重茬能力弱的品种,推测参与防御反应的苹果过敏原蛋白和病程相关蛋白是苹果植株应对连作障碍的关键蛋白。抗重茬能力强的品种通过加强多种不同途径的代谢,增强植株对于连作障碍的抗逆性,减少连作障碍对植物的伤害。
关键词:苹果;连作障碍;蛋白质非标记定量技术;蛋白质组学
中图分类号:S661.1:Q786文献标识号:A文章编号:1001-4942(2017)05-0001-09
Differential Proteomics Analysis on Apple in Response to
Continuous Cropping Obstacle
Ni Wei1, Ni Weiru1, Zhao Jing2, Wang Zenghui1, Mao Zhiquan1, Chen Xuesen1, Shen Xiang1
(1.College of Horticulture Science and Engineering, Shandong Agricultural University/State Key Laboratory of
Crop Biology, Taian 271018,China;2.College of Horticulture, Anhui Agricultural University, Hefei 230000, China)
AbstractThis research aims to explore the mechanism of apple in response to continuous cropping obstacle at the protein expression level by confirming the differential expression proteins of different apple cultivars answering continuous cropping. Stronger replanting-tolerant cultivar Fuji 2001 and weaker replanting-tolerant cultivar Shoufu 1 were grown in pots with replanted soils. Under the same cultivation and management conditions, label-free technology was used to identify the differential expression proteins in the leaves of apple which had different resistance abilities to continuous cropping. The differential expression proteins were compared with GO, COG and other databases to analyze their function and classification. The results showed that 43 kinds of differential or particular expression proteins were identified from two apple cultivars, and were mainly divided into four categories, including one protein related to photosynthesis, seventeen proteins referring to energy and carbohydrate metabolism, eight proteins related to plant defense and resistance, and seventeen proteins with other functions. There were great differences among the proteins of energy and carbohydrate metabolism and plant defense and resistance. Different apple cultivars had different protein expression levels related to continuous cropping. Furthermore, the stronger replanting-tolerant apple cultivar had higher protein expression level than the weaker one. It was inferred that the allergen proteins and pathogenesis related proteins, which joined in defensive reaction, were the key proteins of apple in response to continuous cropping. The stronger replanting-tolerant apple cultivar enhanced various different metabolic pathways to strengthen the resistance to continuous cropping obstacle and reduce the harm to plants.
KeywordsApple; Continuous cropping obstacle; Label-free technology; Proteomics
果园连作障碍又称再植病、重茬病,主要表现为树体生活力降低,枝条生长量小,树冠小,产量低,严重影响果园的更新和可持续发展。研究表明不同苹果品种对于连作障碍的表现不同,但作用机理尚不明确。因此研究不同苹果品种在重茬土中的差异蛋白,从蛋白质水平上研究减轻连作障碍的机理,对于降低连作障碍对苹果植株的影响、实现果园的持续发展有重要意义。目前普遍认为连作障碍产生的原因是综合性的,主要由土壤理化性质的破坏、土壤养分的失衡或缺失、微生物种群改变或根系分泌物的毒害等因素引起[1]。有学者研究发现,在不同苹果砧木资源中存在抗重茬障碍的遗传基因,希望能够利用Malus sieversii等不同的苹果种质获得抗重茬障碍基因,并筛选得到抗重茬砧木[2]。有研究表明,太子参在连作、轮作不同种植方式下,叶片中的蛋白质表达出现差异,连作条件下植物衰老或病害发生相关的五种蛋白在叶片中的表达量均有不同程度的上调,并认为这是太子参连作病害发生和提前黄化的主要原因[2]。而蛋白质非标记定量技术(label-free)检测差异蛋白时在肽段的数量、蛋白质的覆盖率和分析通量方面具有较大优势,且不受样品来源和数量的限制。有研究采用蛋白质非标记定量等技术研究烟草青枯菌致病性,比较分析了非致病和致病青枯菌侵染后烟草的差异蛋白质,并进一步得到显著性量变差异蛋白质163个,富集分析结果表明它们与糖代谢相关或主要定位于细胞外,这对于进一步研究青枯菌侵染烟草的生物学机制有重要作用[3]。目前对于苹果连作障碍的研究大多为植物生理生化方面的研究,蛋白质层面的研究极少。本试验以在重茬地中生长表现不同的两个苹果品种为材料,运用蛋白质非标记定量技术研究苹果中与抗重茬相关的差异蛋白,获得两个苹果品种的差异蛋白图谱,分析差异蛋白的类别和功能,以便能进一步从蛋白质水平上找到解决苹果连作障碍的理论依据。
1材料与方法
1.1试材及取样
试验材料为在重茬地中生长表现较好的富士2001(Fuji 2001)和表现较差的首富1号(Shoufu 1)两个苹果品种[3]。供试植株为以平邑甜茶(M. hupehensis)为基砧、分别以富士2001和首富1号为接穗的两年生植株。选择生长整齐一致的幼苗栽植,重茬土取自泰安市岱岳区滩清湾村20年生苹果园,收集距果树树干1 m之内、深 5~40 cm范围内的土壤,采取多点随机取样的方式以保证试验的准确性,混匀过筛后装在外径35 cm、内径30 cm的透气塑料花盆中,每盆装土10 kg。试验于2015年在山东农业大学观赏果树试验站进行。3月26日将富士2001和首富1号的幼苗栽在装有重茬土的盆中,每个处理10盆,每盆种植3株,进行统一管理。于10月21日采集样品,选取同一生长部位正常生长的苹果叶片,液氮保存带回,-80℃存放,以干冰包装运输到北京邦菲生物科技有限公司进行蛋白质非标记定量试验,每个处理重复3次。
1.2蛋白提取
取适量样品用液氮研磨成粉末状,加入1 mL预冷BPP提取缓冲液,室温涡旋振荡10 min,加入1 mL Tris饱和酚,室温涡旋振荡10 min,4℃、15 000×g离心15 min;转移上清液,并加入等体积BPP提取缓冲液,室温涡旋振荡5 min,4℃、15 000×g离心15 min,重复抽提一次。取1 mL上清液,加入5 mL过饱和硫酸铵甲醇溶液(即AM沉淀剂),于-20℃沉淀过夜,过夜的溶液于4℃、15 000×g离心15 min。加入1 mL冰冷甲醇,用剪过的枪头充分打碎蛋白,于4℃、15 000×g离心5 min,弃上清液;加入1 mL冰冷丙酮,用枪头充分搅碎蛋白块,于4℃、12 000×g离心5 min,弃上清液重复上述操作;蛋白沉淀室温下自然风干约5~8 min,加入适量脲裂解液;将溶解完全的蛋白液于20℃、20 000×g离心30 min,转移上清液至离心管中,4℃或-20℃保存备用。
1.3酶解
取50 μL提取的蛋白质样品,加入 DTT至终浓度为100 mmol/L,沸水浴5 min,冷却至室温。加入200 μL UA buffer(8 mol/L Urea,150 mmol/L Tris-HCl pH值8.0)混匀,转入10 kD超滤离心管,14 000×g离心15 min,弃滤液。加入200 μL UA buffer,14 000×g離心15 min,弃滤液。加入100 μL IAA(50 mmol/L IAA in UA),600 r/min振荡1 min,室温避光30 min,14 000×g离心10 min,弃滤液。加入100 μL UA buffer,14 000×g离心10 min,弃滤液,重复2次。加入100 μL 100 mmol/L NH4HCO3,14 000×g离心10 min,弃滤液,重复2次。加入40 μL Trypsin buffer(4 μL Trypsin in 40 μL NH4HCO3),600 r/min振荡1 min,37℃放置16~18 h。换新收集管,14 000×g离心10 min,取滤液,测定在280 nm处的吸光度值,对蛋白质的肽段进行定量分析。
1.4酶解产物的LC-MS/MS分析
蛋白质FASP酶解后,按照定量结果取2 μg酶解后产物进行LC-MS/MS分析。采用纳升流速HPLC液相系统EASY-nLC1000进行分离。液相A液为0.1%甲酸乙腈水溶液(乙腈为2%),B液为0.1%甲酸乙腈水溶液(乙腈为84%)。色谱柱Thermo EASY column SC200 150 μm×100 mm (RP-C18)以100%的A液平衡。样品由自动进样器上样到Thermo EASY column SC001 traps 150 μm×20 mm (RP-C18)(Thermo),再经色谱柱分离,流速为300 nL/min。相关液相梯度如下:0~105 min,B液线性梯度从0~45%;105~110 min,B液线性梯度从45%~100%;110~120 min,B液维持在100%。酶解产物经毛细管高效液相色谱分离后用Q-Exactive质谱仪(Thermo Finnigan)进行质谱分析。分析时长:120 min;检测方式:正离子;母离子扫描范围:300~1 800 m/z;多肽和多肽碎片的质量电荷比按照下列方法采集:MS1在M/Z 200时分辨率为70 000;AGC target:3e6;一级Maximum IT:10 ms;Number of scan ranges:1;Dynamic exclusion:40.0 s。每次全扫描(full scan)后采集10个碎片图谱,二级在M/Z 200时分辨率为17 500,MS/MS Activation Type:HCD;Isolation window:2 m/z;Microscans:1;二级Maximum IT:60 ms;Normalized collision energy:30 eV;Underfill ratio:0.1%。
1.5Maxquant的非标记分析
每个样品重复3次质谱分析,原始文件导入Maxquant软件(版本号1.3.0.5)进行查库,进行LFQ非标记定量分析。搜索使用的数据库为Malus mill。主要参数如下,Main search ppm:6;Missed cleavage:2;MS/MS tolerance ppm:20;De-Isotopic: TRUE;enzyme:Trypsin;database:uniprot_rat_35702_20151029.fasta;Fixed modification:Carbamidomethyl(C);Variable modification: Oxidation(M),Acetyl (Protein N-term);Decoy database pattern: reverse;Lable free quantification(LFQ):TRUE;LFQ min ratio count:1;Match between runs:2 min;Peptide FDR:0.01;Protein FDR:0.01。
1.6Perseus的统计学和生物信息学分析
Maxquant所得的查库文件使用Perseus软件进行分析,Perseus软件版本号为1.3.0.5。
2结果与分析
2.1肽段和蛋白质分析
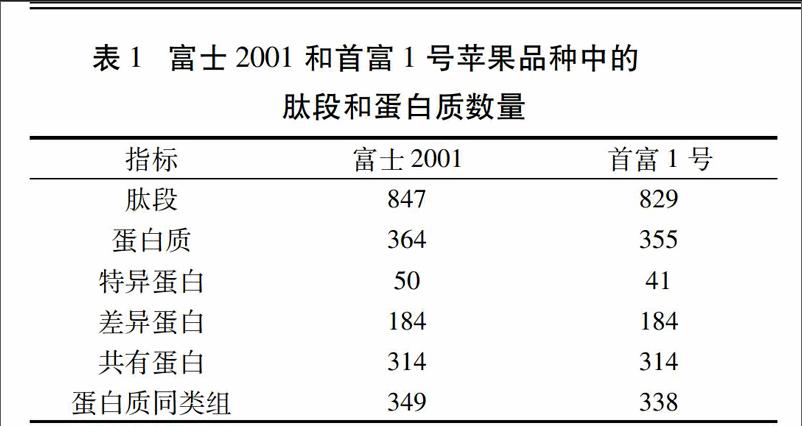
富士2001和首富1号肽段和蛋白质鉴定结果见表1。可以看出,富士2001和首富1号中鉴定得到的肽段数分别为847和829,而蛋白质总数分别为364和355,对应的蛋白质同类组数为349和338。本试验共鉴定得到了184个差异蛋白(差异倍数≥1.5或差异倍数≤0.667),其中与首富1号相比,富士2001中144个为上调蛋白,40个为下调蛋白。富士2001和首富1号鉴定到的共有蛋白为314个,富士2001中存在而首富1号中未检测到的特异蛋白为50个,首富1号中的特异蛋白数量为41个。这些结果表明抗重茬表现不同的两个苹果品种在蛋白质表达方面也具有较大的差异。
2.2蛋白质组GO功能分类
基因本体论(Gene Ontology,GO)数据库通过建立一套具有动态形式的控制字集,来解释真核基因及蛋白质在细胞内所扮演的角色,从而全面描述生物体中基因和基因产物的属性[4]。结合GO数据库对两个苹果品种的差异蛋白进行功能分类,从宏观上认识抗重茬表现不同的苹果品种蛋白质功能分布特征。富士2001和首富1号表达量差异蛋白的GO分类统计如图1所示,差异表达蛋白在细胞组分、分子功能和生物学过程三大类中分别包含了13、15个和23个功能小类。在生物学过程这一类中,生物学调控和代谢过程占的比例较高;在细胞组分中,胞内区域成分和细胞骨架占的比例较大;在分子功能中,催化活性和结合活性占的比例较高。富士2001特异蛋白GO分类统计如图2所示,特异蛋白在生物学过程、细胞组分和分子功能三类分别包含14、6个和7个功能小类。在生物学过程这一类中,刺激应答、生物学调控、代谢过程、应答生物性刺激占的比例较高;在细胞组分中,膜、细胞成分、胞外区域成分占的比例较大;在分子功能中,催化活性和结合活性占的比例较大。首富1号特异蛋白GO分类统计如图3所示,特异蛋白在生物学过程、细胞组分和分子功能三类分别包含12、6个和10个功能小类。在生物学过程这一类中,细胞代谢过程、生物学调控和代谢过程的比例较高;在细胞组分中,类核占的比例较大;在分子功能中,基因结合转录因子活性、催化活性和转移酶活性占的比例较大。
2.3蛋白质组COG功能分类
在生物体内,蛋白质不能独立地行使功能,而是不同蛋白质相互协调完成一系列生化反应以行使其生物学功能[5]。COG(Cluster of Orthologous Groups of proteins 蛋白相邻类的聚簇)是对蛋白质进行直系同源分类的数据库。将两个苹果品种差异蛋白与COG 数据库进行比对,预测蛋白质的功能并进行分类统计。如图4所示,两个品种表达量差异蛋白根据功能可分为19个大类,其中一般功能预测包含的差异蛋白数量最多,为30个,其次是次生代谢物的生物合成、运输和分解(19)、细胞骨架(18)、能量的产生和转化(17),其他类别的差异蛋白数量各不相同。如图5所示,富士2001特异蛋白根据功能可分为8个大类,主要为次生代谢物的生物合成、运输和分解(13),其他种类的特异蛋白数量均较低。如图6所示,
2.4差异蛋白的表达变化及功能分类
对质谱分析得到的两个苹果品种的表达量差异蛋白和特异蛋白进行数据分析、序列比對、结果注释和文献检索鉴定,主要得到43种差异或特异表达蛋白,在表达量差异蛋白中共鉴定得到25种蛋白存在显著性差异,富士2001中特异蛋白为13种,首富1号中特异蛋白为9种(过敏原蛋白类和病程相关蛋白类在表达量差异蛋白和特异蛋白中都鉴定得到)。这些蛋白按照功能主要分为4类,具体的差异蛋白表达量见。
第1类:包括1种与光合作用相关的蛋白,为叶绿素a/b结合蛋白Ⅰ型,在富士2001中表达量显著上调。
第2类:与植物能量及代谢相关的蛋白为17种。包括糖酸组分代谢有关的蛋白,除山梨醇6-磷酸脱氢酶和部分苹果酸酶在富士2001中显著下调外,其余苹果酸脱氢酶、果糖激酶、葡萄糖激酶均在富士2001中显著上调,而山梨醇脱氢酶在富士2001中特异表达。类黄酮代谢相关的黄酮醇合酶、二氢黄酮醇-4-还原酶、花青素合成酶、花青素还原酶均在富士2001中显著上调,而二磷酸尿核苷葡萄糖-类黄酮3-O-葡糖基转移酶、查尔酮合成酶仅在首富1号中特异表达。抗坏血酸代谢相关的除L-半乳糖酶在富士2001中显著下降外,其余L-半乳糖-1-磷酸酶、GDP-D-甘露糖焦磷酸酶、单脱氢抗坏血酸还原酶均在富士2001中显著上升,与能量代谢有关的糖基转移酶只在首富1号中特异表达。
第3类:与植物防御、抗逆性相关的蛋白有8种。其中病程相关类(pathogenesis-related,PR)蛋白分别为病程相关蛋白、β-1,3-葡聚糖酶、类甜蛋白和苹果过敏原蛋白和其他防御相关蛋白如防御素、多酚氧化酶、脱水蛋白和自噬相关蛋白。除少量过敏原蛋白和病程相关蛋白在富士2001中显著下调和在首富1号中特异表达外,其余的多酚氧化酶、防御素均在富士2001中显著上调,而脱水蛋白、自噬相关蛋白、类甜蛋白、β-1,3-葡聚糖酶仅在富士2001中特异表达。
第4类:其他功能蛋白包含17种,分别在植物的生物合成与调控、细胞定位与识别、细胞信号传递等功能发挥不同的作用,通过多种代谢途径提高植物的抗逆能力。
3讨论与结论
3.1光合作用相关蛋白对苹果抗重茬的影响
叶片通过光合作用进行物质积累和能量代谢,通过光信号传导过程调节植物各种生理生化代谢途径,而蛋白质正是这些生理生化反应的最直接参与者[6]。有研究表明植物在受到逆境胁迫时,类囊体的结构会遭到破坏,引起膜脂过氧化和叶绿素含量的减低,导致光合速率下降,同样苹果植株在受到连作障碍胁迫时,光合速率也会下降[7]。刘淑芹等[8]在番茄重茬土中套作分蘖洋葱后,相比于对照,番茄植株中10种与光合作用相关的蛋白显著上升,明显减低了连作障碍对番茄生长的影响。在本试验中仅检测到一种与光合作用相关的蛋白,即叶绿素a/b结合蛋白Ⅰ型,在富士2001中的表达量显著高于首富1号,绿色植物通过捕光叶绿素结合蛋白接收太阳能来同化CO2获得能量。分析原因是因为富士2001在重茬土中的光合作用能力较强,预示生成更多的总糖、还原糖等碳水化合物,营养物质储存更加丰富,有利于增强对不良环境的抵抗能力,这与我们先前在重茬土中测量苹果植株净光合速率的结果基本一致。但由于本试验中未鉴定出其他关于光合作用的差异蛋白,其他有关蛋白是否在苹果抗连作障碍中发挥重要作用,还有待于进一步研究。
3.2防御、抗逆相关蛋白对苹果抗重茬的影响
在本试验中苹果抗重茬能力主要体现在两个方面:一方面是植株体内与自身代谢活动或者能量转化相关蛋白的表达量存在显著差异,另一方面是能提高植株抗逆性、防御外界环境生物或非生物胁迫相关蛋白的表达量存在显著差异。
在植株自身的代谢活动中,糖酸代谢具有重要的作用。林茂兹[2]对太子参连作与轮作的多糖和人参皂苷Rb1含量的比较结果说明太子参连作导致其品质急剧下降。有研究发现一定浓度的草酸和苹果酸能够促进平邑甜茶幼苗的生物量增长、光合作用等,减轻连作障碍对幼苗生长的影响[9,10]。在本试验中共鉴定得到5种糖酸代谢相关的差异表达蛋白,分別参与植物三羧酸、糖酵解等不同的代谢途径,蛋白分别为苹果酸酶、苹果酸脱氢酶、果糖激酶、葡萄糖激酶、山梨醇脱氢酶、山梨醇6-磷酸脱氢酶,还有1种能量代谢相关的蛋白为糖基转移酶。除山梨醇6-磷酸脱氢酶和部分苹果酸酶在富士2001中显著下调外,其余苹果酸脱氢酶、果糖激酶、葡萄糖激酶均在富士2001中显著上调,山梨醇脱氢酶在富士2001中特异表达,而糖基转移酶在首富1号中特异表达。糖基转移酶具有多种形式,大量分布在生物体内,是合成糖甙键的关键酶,对于合成糖蛋白、糖脂及其他次生代谢产物有重要作用,糖基化作用还可以抑制和排除生物体内大量内生和外生有毒物质的合成与代谢[11]。这些结果表明在应答重茬胁迫的过程中,苹果植株可能通过糖酸类化合物及能量的代谢反应为植株抗重茬障碍实现物质基础。
抗坏血酸在植物体内具有重要的抗氧化作用,能够清除自身的活性氧和自由基,减少逆境胁迫对于植株的伤害,保证正常的代谢功能,植物体能自主合成抗坏血酸,若植物体内抗坏血酸代谢不足或受阻,植物的抗逆性和生长发育都会受到限制[12]。潘德灼等[13]研究发现抗坏血酸代谢相关酶在龙眼果皮采后褐变过程中表达量不断下调,导致细胞中积累较多的活性氧,从而加速了果皮的衰老。本试验中共鉴定得到4种抗坏血酸代谢相关蛋白,分别为L-半乳糖酶、L-半乳糖-1-磷酸酶、GDP-D-甘露糖焦磷酸酶、单脱氢抗坏血酸还原酶,除L-半乳糖酶在富士2001中显著下调外,其余L-半乳糖-1-磷酸酶、GDP-D-甘露糖焦磷酸酶、单脱氢抗坏血酸还原酶均在富士2001中显著上调。表明在重茬土逆境胁迫中,苹果植株可能通过抗坏血酸代谢途径减低活性氧的浓度,从而减轻自由基对植株的伤害,所以抗坏血酸参与的抗氧化过程也是苹果应对连作障碍的重要防御反应。
类黄酮生物合成途径在植物体内普遍存在,能够产生大量的次生代谢产物,清除体内自由基,减少膜脂过氧化程度,在植物着色物质、防御素方面有重要的作用[14]。有研究证明在杨梅中花色苷合成与糖代谢途径中关键蛋白表达上调是果肉花色苷合成和颜色形成的重要生理机制[15,16]。在本研究中,共鉴定得到6类与类黄酮代谢相关的蛋白,黄酮醇合酶、二氢黄酮醇-4-还原酶、花青素合成酶、花青素还原酶均在富士2001中显著上调,而二磷酸尿核苷葡萄糖-类黄酮3-O-葡糖基转移酶、查尔酮合成酶仅在首富1号中特异表达。这都表明了苹果植株在重茬土中能够通过增加体内类黄酮代谢相关酶的表达,代谢生成不同的次生代谢产物,提高植物的抗氧化作用,从而抵御外界不利环境对于植株生长的胁迫,减少连作障碍对植株生长的不良影响。
病程相关(pathogenesis-related,PR)蛋白是一类在植物中具有广泛抗性的可溶性蛋白[17],根据PR蛋白的植物来源、氨基酸序列同源性以及生化功能等,可将PR蛋白分为17类,其中包括富含甘氨酸的病程相关蛋白(PR-1)、β-1,3-葡聚糖酶(PR-2)、几丁质酶(PR-3)、类甜蛋白(PR-5)、过氧化物酶类(PR-9)、过敏反应相关的PR蛋白(PR-10)等[18,19]。本试验中共鉴定到四类PR蛋白,分别为病程相关蛋白、β-1,3-葡聚糖酶、类甜蛋白和苹果过敏原蛋白。除少量过敏原蛋白和病程相关蛋白在富士2001显著下调和在首富1号中特异表达外,其余的多酚氧化酶、防御素均在富士2001中显著上调,这些PR蛋白对于苹果植株应对生物和非生物胁迫具有重要作用。张彩霞等[20]用轮纹病菌侵染苹果枝条,发现枝条中的病程相关类蛋白表达显著上升,表明其在苹果枝条应答轮纹病菌的胁迫反应中发挥了重要作用,参与了枝条表皮细胞的抗病反应。
如上所述,苹果植株在应对连作障碍的胁迫过程中,多个代谢途径的蛋白差异表达或特异表达,包括光合作用、糖类及能量代谢、抗坏血酸代谢、类黄酮代谢及防御、抗逆性反应等相关蛋白。推测参与防御反应的苹果过敏原蛋白和病程相关蛋白是苹果植株应对连作障碍的关键蛋白,提高了苹果对外界环境的抵御能力,能够应对生物和非生物胁迫,这与刘淑芹等[8]的研究結果相近。本试验从蛋白质水平上初步了解了苹果植株应对连作障碍胁迫时不同抗性相关蛋白的差异表达情况,可为进一步提高苹果连作障碍抗性的种质改良提供理论基础。
参考文献:
[1]Wilson S, Andrews P, Nair T S. Non-fumigant management of apple replant disease[J]. Scientia Horticulturae, 2004, 102(2): 221-231.
[2]林茂兹. 太子参连作障碍及其分子生态学机制研究[D].福州:福建农林大学,2010.
[3]倪蔚茹,王安然,贺锡燕,等.重茬土对相同砧木不同苹果品种生理指标及叶片抗氧化酶活性的影响[J].中国农业科学,2016,49(18):3597-3607.
[4]贾新平,孙晓波,邓衍明,等.鸟巢蕨转录组高通量测序及分析[J]. 园艺学报, 2014, 41(11):2329-2341.
[5]孙倩倩.外源褪黑素对番茄果实采后成熟的影响[D]. 北京:中国农业大学,2016.
[6]蔡永占, 周普雄, 李佛琳,等. 不同气候环境中团棵期烟草叶片蛋白质组学分析[J].中国农业科学, 2013, 46(4): 859-870.
[7]尹承苗,张先富,胡艳丽,等.不同浓度有机物料发酵流体对连作苹果幼树叶片光合荧光参数和根系抗氧化酶活性的影响[J].中国农业科学, 2014, 47(9):1847-1857.
[8]刘淑芹,周新刚,吴凤芝,等.分蘖洋葱-番茄套作系统中番茄叶片差异蛋白表达分析[J].中国农业科学, 2016, 49(5):919-932.
[9]李俊芝,王功帅,胡艳丽,等.几种低分子量有机酸对连作平邑甜茶幼苗光合与根系生长的影响[J]. 园艺学报, 2014, 41(12):2489-2496.
[10]王红燕,佟海英,黄苏珍,等.柠檬酸和草酸对Pb 胁迫下马蔺生长和生理的影响[J]. 生态学杂志, 2010, 29 (7):1340-1346.
[11]Hong Z L, Zhang Z M, Olson J M, et al. A novel UDP-glucose transferase is part of the callose synthase complex and interacts with phragmoplastin at the forming cell plate[J]. Plant Cell, 2001, 13:769-780.
[12]盖瑞,郭小静,张贞,等.不同花粉对苹果果实中抗坏血酸相关酶活性的影响[J]. 林业科学,2014, 50 (11):36-43.
[13]潘德灼,盖永红,吕晓杰,等.龙眼果实采后果皮褐变过程差异表达蛋白分析[J]. 果树学报, 2016, 33(5):542-551.
[14]张丽,罗海波,姜丽,等. 果实成熟衰老过程中蛋白质组学研究进展[J]. 植物生理学报, 2011, 47(9): 861-871.
[15]Abrahams S,Lee E,Walker A R,et al. The Arabidopsis TDS4 gene encodes leucoanthocyanidin dioxygenase(LDOX)and is essential for proanthocyanidin synthesis and vacuole development[J]. The Plant Journal,2003, 35 (5):624-636.
[16]张泽煌 ,陈义勇,钟秋珍,等.红果肉与白果肉杨梅花青苷和糖代谢途径的差异蛋白研究[J].园艺学报, 2013, 40(12):2391-2400.
[17]Soh H C, Park A R, Park S, et al.Comparative analysis of pathogenesis-related protein 10 (PR10) genes between fungal resistant and susceptible peppers[J]. European Journal of Plant Pathology, 2012, 132: 37-48.
[18]Loon L C,Rep M,Pieterse C M.Significance of inducible defense-related proteins in infected plants[J]. Annual Review of Phytopatholoy,2006, 44: 135-162.
[19]林茂兹, 张志兴, 林争春,等. 太子参连作障碍蛋白差异表达分析[J]. 草业学报, 2010, 19(6): 197-207.
[20]张彩霞,田义,张利义,等.苹果枝条表皮应答轮纹病菌侵染的蛋白质组学分析[J]. 植物病理学报, 2015, 45(3):280-287.
