镉胁迫水稻潜热通量与冠层结构参数变化遥感分析
2017-06-05罗力兵刘湘南吴伶刘美玲
罗力兵,刘湘南,吴伶,刘美玲
(中国地质大学(北京)信息工程学院,北京 100083)
镉胁迫水稻潜热通量与冠层结构参数变化遥感分析
罗力兵,刘湘南*,吴伶,刘美玲
(中国地质大学(北京)信息工程学院,北京 100083)
为了更准确地监测农田水稻镉污染状况,结合水稻冠层能量平衡和结构参数LAI(Leafarea index),构建水稻生理功能敏感指数Pf(Physiological function sensitivity index),分析水稻镉胁迫水平:首先利用遥感影像数据和实测数据,基于能量平衡模型模拟实验区水稻潜热通量LE(Latentheat flux),然后结合LE与LAI构建水稻生理功能敏感指数Pf,最后根据Pf识别水稻镉胁迫。结果表明,镉胁迫越重水稻LE、LAI和Pf值越小,LE可将水稻镉胁迫区分为重度和轻度/中度两个水平;LAI可将水稻镉胁迫区分为中度/重度和轻度胁迫两个水平;Pf能更好地将水稻镉胁迫区分为轻度、中度和重度三个水平,其区分准确度分别为70.59%、69.05%和84.79%。Pf能有效诊断并定量分析水稻镉污染胁迫状况,为作物重金属污染监测提供重要参考。
镉胁迫;能量平衡;潜热通量;LAI;胁迫指数
农业重金属污染是重大生态环境问题之一,直接影响着农业生产和粮食安全。如何准确快速获取某区域的水稻重金属污染信息,成为研究者目前关注的重点。传统获取水稻重金属污染信息的主要方法是采样和化验,其成本高并且难以大面积获取。重金属胁迫会抑制水稻的生理生长,从而导致不同重金属胁迫下水稻生理功能特征和生理要素特征有所差异[1]。利用遥感方法可以大面积获取水稻的生理参量,从而实现重金属胁迫大面积监测。目前利用遥感监测水稻重金属胁迫的研究主要是基于可见光、近红外、微波和热红外构建水稻叶绿素含量、叶面积指数、生物量[2]和生理功能[3]等水稻重金属胁迫敏感指数来监测重金属污染,然而这些研究侧重从生理要素的角度监测水稻重金属胁迫,并未从水稻生长机理的角度结合生理过程和生化参数。为了准确获取水稻重金属胁迫程度和分布信息,本文通过结合水稻冠层能量平衡和结构参数LAI(Leaf area index)的变化来分析水稻镉污染胁迫。
蒸腾作用是植被独有的生理机制,是植被与外界能量交换的主要生理方式,也是植被能量释放的关键途径。镉胁迫会干扰植被叶片气孔开放,抑制植被蒸腾速率,当植被蒸腾速率受到抑制则冠层辐射能量增加,反之则减少[4]。镉胁迫也会导致水稻LAI产生差异,LAI是重要的水稻冠层结构参数,其可以表征水稻的生理活性和水稻冠层能量交换[5]。本研究基于研究区Landsat-8影像数据、实测数据和气象数据,利用能量平衡模型和经验模型分别模拟水稻潜热通量和反演水稻LAI,最后结合水稻冠层结构对水稻生理功能的影响,构建水稻生理功能镉胁迫指数,进而分析水稻镉污染程度。
1 材料与方法
1.1 实验设计
我国湖南省湘江流域重金属污染严重,其中镉、锌、铜、镍、铅等均超出国家标准[6],本研究区位于湘江流域典型重金属污染区——湖南省株洲地区。在研究区选取A、B、C三个实验区,每个实验区面积均为200m2,水稻品种为博优9083,土壤为红壤,显酸性,母质为第四纪红土红壤,系亚热带季风气候,雨量充沛、光热充足,在人工管理下水田土壤肥沃,灌溉充足。野外重金属含量样本化验结果见表1。实验区A、B和C土壤镉含量均超出国家二级标准,C区最为严重,根据化验结果将A、B、C依次定为轻度镉污染区、中度镉污染区和重度镉污染区。研究区与试验区地理位置如图1所示,其中a图为中国地理图,b图为株洲市,c图为研究区图,c图中的A、B和C分别表示试验区地理位置分布。
1.2 实验数据
研究数据主要分为影像数据、环境数据和实测数据。影像数据为2014年9月4日株洲区域Landsat-8影像数据,包括可见光、近红外和热红外波段;环境数据为同日株洲气象站观测的气温值和风速值;实测数据为2014年9月所测成熟期水稻数据,主要包括LAI、水稻株高、土壤重金属含量。LAI数据采集:在研究区稻田中分别均匀采集30组LAI值,取其平均值作为样地LAI值,测量仪器为AccuPAR LP-80植被冠层分析仪,测量方式为垂直测量。土壤重金属含量是在野外试验区分别均匀采集多个土壤样本后,在实验室化验获取(原子吸收法)。
1.3 方法
1.3.1 基于能量平衡模型的水稻潜热通量模拟
基于能量平衡模型模拟地表蒸散的遥感方法,把遥感数据反演的地表参数以及大气参数输入模型中,计算潜热通量。目前基于能量平衡模型计算蒸散的方法主要有单层模型、双层模型[7]和多层模型[8]。本研究的重点是利用水稻生理功能的变化分析水稻镉污染胁迫,为突出水稻生理要素对水稻生理功能的影响,研究时段选择在水稻成熟期,能量平衡模型选用单层模型。水稻冠层能量平衡是水稻自身生理功能调节与外在环境因素共同作用的结果,其外在环境影响因素主要有湿度、气温和风速,内在影响因素主要有蒸腾作用和光合作用[9]。本研究的水稻品种、生长环境和管理措施均相似,主要差异为试验区之间土壤镉含量和水稻冠层结构差异。本文基于能量平衡模型,在典型环境条件下模拟水稻潜热通量,模拟镉胁迫对水稻生理过程的影响。能量平衡方程为:
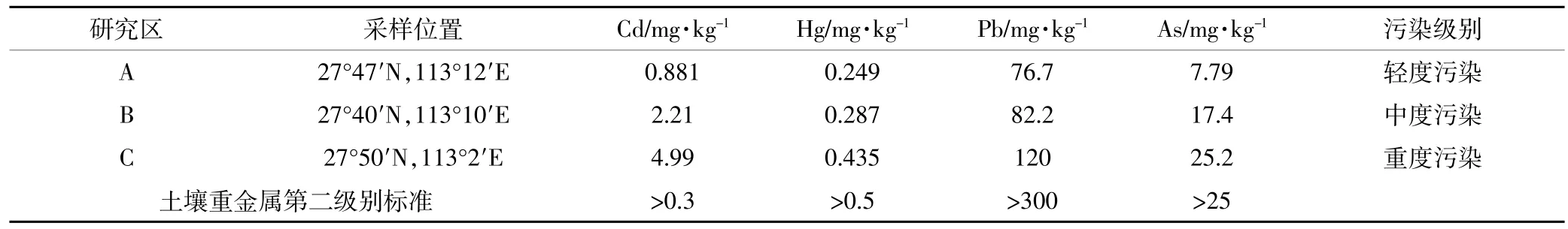
表1 实验区土壤重金属含量Table 1 Heavymetal content in soilofexperimentalarea
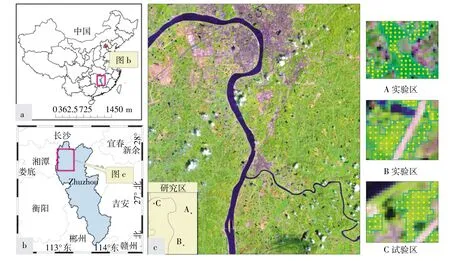
图1 研究区地理位置和实验区分布Figure 1 Location of the study area and the distribution ofexperimentalsite

式中:LE表示潜热通量;Rn代表地表净辐射能量,是各种波长的辐射能在地表的收支平衡;H表示显热通量;G表示土壤热通量。
吸收的能量主要来源于太阳辐射和太阳散射辐射,释放的能量主要是植被自身辐射、显热通量、土壤热通量和潜热通量。存储的能量占总能量比例小,一般可以忽略。
Rn与冠层辐射能量求取方程为:
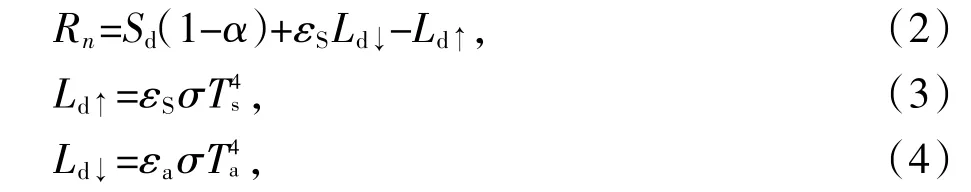
式中:Sd是太阳总辐射,Sd=Gsc×cosθ×dr×τsw(Gsc是太阳常量1367W·m-2;cosθ表示太阳天顶角余弦值;dr为日地距离因子,数值可以在Landsat-8头文件中获取;τsw为大气透过率);α是地表反照率[10];εS是地表比辐射率;Ld↓是地表下行辐射;Ld↑水稻冠层辐射能量或称地表上行辐射;σ为斯蒂芬-玻耳兹曼常数;εa是空气比辐射率[11];Ts是水稻冠层温度,K,由于温度是本研究的主要参数,反演方法将在下文详细介绍;Ta为空气温度,本文取值为株洲气象站观测的日平均气温299.15 K。
显热通量(H)计算公式如下:

式中:ρ是空气密度,取值1.131 kg·m-3;Cp表示空气定压比热容,其值为1004 J·kg-1·K-1,ΔT表示冠层温度和空气温度之差,K;ra为空气动力学阻抗[12-14],s·m-1。
土壤热通量(G)计算公式如下:

式中:Γc=0.315,Γs=0.05,分别表示裸土和完全植被覆盖时潜热通量占净辐射的比例;fc表示植被覆盖度。
1.3.2 温度反演
目前地表温度反演方法主要有单窗算法、劈窗算法和辐射传输方程法,其中辐射传输方程法比较适合用于水温反演,由于Landsat-8热红外波段定标系数仍不稳定,美国地质勘探局(United States Geological Survey,USGS)提倡用单波段来反演温度。本文温度计算采用由Jimenez-Munoz等[16]提出的单通道算法SC(Single-channelMethod),主要方程如下:
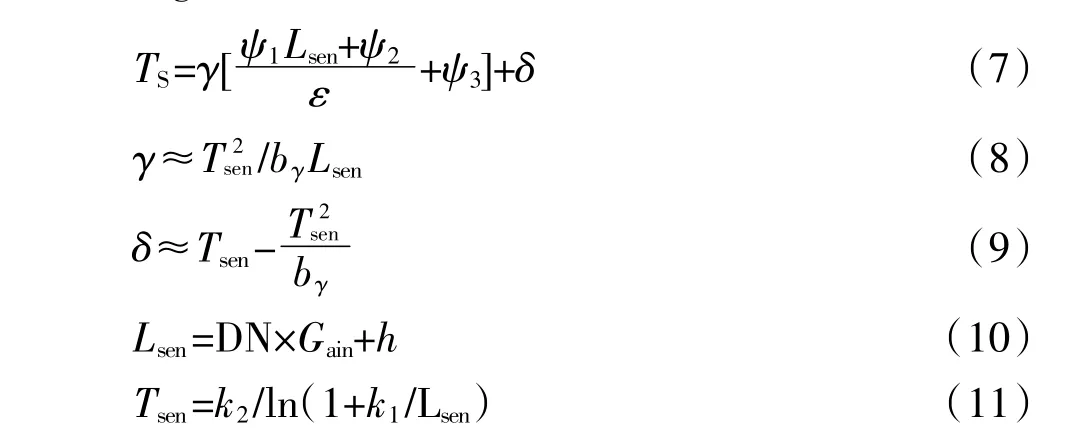
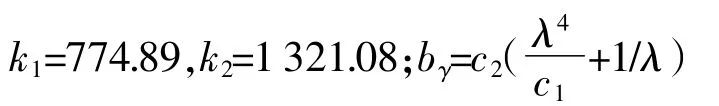

ω为大气含水量,取值2.4 g·cm-2,参考覃志豪等[18]大气估算研究结果。
1.3.3 水稻LAI反演
遥感反演LAI的主要方法有经验模型、半经验模型和物理模型等。本文采用经验模型,首先将地面实测LAI与相应试验区Landsat-8影像植被指数GBNDVI进行指数函数拟合,再根据拟合方程反演LAI[19],主要公式如下:

式中:NIR表示近红外波段,Green表示绿波段,Blue表示蓝波段;y表示LAI的值,x表示指数GBNDVI的值。
1.3.4 水稻生理功能敏感指数构建
水稻生理功能敏感指数的构建主要是基于水稻LE和LAI。在水稻生长过程中,镉污染胁迫会抑制水稻生理功能,影响水稻能量平衡,导致水稻能量吸收减少,从而抑制水稻营养器官的生长,具体体现在LAI、叶绿素含量和生物量等生理要素。水稻生理要素又会影响水稻生理功能,镉污染越重,则水稻的生理功能越弱,生理要素越小,随着水稻的生长其差异越明显[2]。由于LAI是水稻重要的冠层结构参数,其可以表征水稻的生理活性和影响水稻冠层能量交换[5]。蒸腾作用是水稻与外界能量交换的重要方式,能很好地表征水稻生理功能强弱。针对镉胁迫特性,结合水稻LE与LAI构建水稻生理功能镉胁迫指数Pf,根据该指数差异可区分不同镉胁迫,其计算式如下:式中:LAImax表示轻度镉胁迫下最大叶面积指数值,LEmax表示轻度镉胁迫下最大潜热通量值。

2 结果分析
2.1 镉胁迫对水稻LE的影响
根据能量平衡模型模拟三个试验区水稻LE,结果如图2所示。轻度胁迫与中度胁迫下水稻LE分布范围相近,但轻度胁迫水平的均值、中位数、下四分位数和上四分位数的统计值比中度胁迫高,说明在本研究的观测尺度下中度镉胁迫对水稻LE响应并不明显,而重度胁迫对水稻LE的影响比较明显,从而可以很好区分水稻重度胁迫与其他胁迫。据此将图2中虚线值定为重度胁迫与其他胁迫阈值,LE值为375 W·m-2。从试验区LE整体分布规律可知,镉胁迫越高水稻LE越小,证明镉胁迫会抑制水稻蒸腾作用,从而导致蒸腾散热减少。由于受镉胁迫的影响,水稻冠层能量平衡中其他组分能量会有所差异,其中冠层辐射能量如图3所示,其与镉胁迫正相关,主要源于蒸腾受到抑制,水稻要以辐射的方式散失多余的热量。
2.2 镉胁迫对LAI与指数Pf结果的影响
试验区LAI与镉胁迫敏感分析如图4所示。中度胁迫与重度胁迫下水稻LAI值不易区分,但中度胁迫的统计值高于重度胁迫LAI值,轻度胁迫下水稻的LAI值高于中度与重度胁迫,说明在本研究观测尺度下基于LAI能很好区分轻度镉胁迫与其他镉胁迫,将图4中虚线值定为轻度胁迫与其他胁迫的阈值,LAI值为4.51。从试验区LAI整体分布规律可知,镉胁迫越重LAI值越低。由于试验区之间主要差异为镉含量,故LAI的差异能间接反映镉长期胁迫的结果。

图2 LE与镉胁迫敏感性分析Figure 2 Sensitivity analysisof LE to cadmium stress
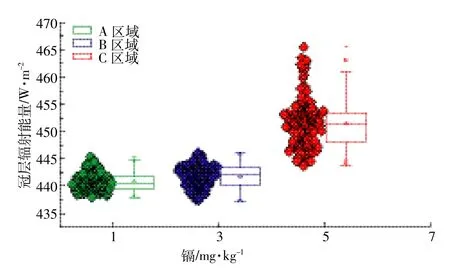
图3 冠层辐射与镉胁迫敏感性分析Figure 3 Sensitiveanalysisof canopy radiation to cadmium stress
根据LE与LAI求取水稻生理功能镉胁迫敏感指数Pf,结果如图5所示。不同镉胁迫下试验区指数分布差异比较明显,能较好区分不同镉胁迫。根据试验区之间Pf的分布,将该图中线c和d的值定为不同胁迫水平之间的阈值(分别为0.44和0.61),则胁迫水平范围为Pf(重度胁迫)<0.44<Pf(中度胁迫)<0.61<Pf(轻度胁迫),根据划分的区间范围计算得轻度胁迫、中度胁迫和重度胁迫的区分准确度分别为70.59%、69.05%和84.79%。从试验区Pf整体分布规律可知,镉胁迫越高,指数Pf值越小,基于指数Pf能很好区分不同镉胁迫。
2.3 镉胁迫对研究区水稻LE、LAI和Pf的影响

图4 LAI与镉胁迫敏感性分析Figure4 Sensitive analysisof LAIto cadmium stress

图5 Pf与镉胁迫敏感性分析Figure 5 Sensitive analysisof Pfto cadmium stress
为实现镉胁迫大面积监测,先将研究区遥感影像进行监督分类,统计得水稻区域为15.81%。根据能量平衡模型模拟整个研究区水稻LE,根据试验区不同胁迫LE阈值(图2),将胁迫水平划分为LE(重度胁迫)<375W·m-2<LE(轻度/中度胁迫),面积分别占研究区总面积的4.15%和11.66%。研究区LE分布如图6a所示,其中重度镉胁迫主要分布在湘江附近。根据拟合方程计算研究区水稻LAI,根据试验区所得不同胁迫LAI阈值(图4),将胁迫水平划分为LAI(中度/重度胁迫)<4.51<LAI(轻度胁迫),其面积分别占研究区总面积的10.71%和5.10%,分布如图6b所示,其中轻度镉胁迫区域主要分布在远离湘江的区域。根据研究区水稻LE与LAI计算指数Pf,根据试验区指数Pf所得不同胁迫阈值(图5),将胁迫水平划分为Pf(重度胁迫)<0.44<Pf(中度胁迫)<0.61<Pf(轻度胁迫),其面积分别为研究区总面积的3.79%、6.28%和5.73%,分布如图6c所示,与LE与LAI值相比,指数Pf对区域的识别更加清晰,其中重度镉胁迫区域主要分布在湘江附近,随着与湘江距离变大,水稻由重度镉胁迫渐变为轻度镉胁迫。
3 讨论
本研究中研究区水稻品种相同、生长环境和管理措施均相似,所以实验区之间主要差异为土壤镉含量和水稻冠层结构差异。目前Liu等[3]从水稻冠气温差的角度说明镉胁迫对水稻生理功能变化的影响,但没有从能量平衡的角度深入分析镉胁迫对水稻蒸腾散热的影响。镉胁迫会干扰植被叶片气孔开放,从而抑制植被蒸腾速率,在植被冠层能量平衡中主要体现为蒸腾散热减弱,本研究通过模拟不同镉胁迫下水稻LE,发现镉胁迫越重水稻LE越小(图2),证明镉胁迫会抑制水稻蒸腾作用。如今很多学者通过水稻冠层温度来间接感应水稻生理功能强弱[3-4],其原理主要是水稻生理功能受到抑制,水稻要以辐射的方式散失多余的热量,所以镉胁迫越重,冠层辐射越强(图3),符合水稻蒸腾作用被抑制的能量平衡规律。
镉胁迫会抑制水稻生理生长,从而导致水稻的生理参数发生改变,所以试验区镉胁迫越重LAI值越小(图4)。由于LAI也是水稻重要的冠层结构参数,可以表征水稻的生理活性和影响水稻冠层能量交换。本研究基于水稻冠层结构差异与土壤镉含量差异构建水稻生理功能镉胁迫指数Pf(图5),镉胁迫越高,该指数值越小,并能很好区分不同镉胁迫。研究区指数Pf值空间分布如图6c所示,湘江是镉胁迫水源污染的重要来源,所以镉胁迫区域主要分布在湘江附近,随着与湘江距离变大,水稻由重度镉胁迫渐变为轻度镉胁迫。
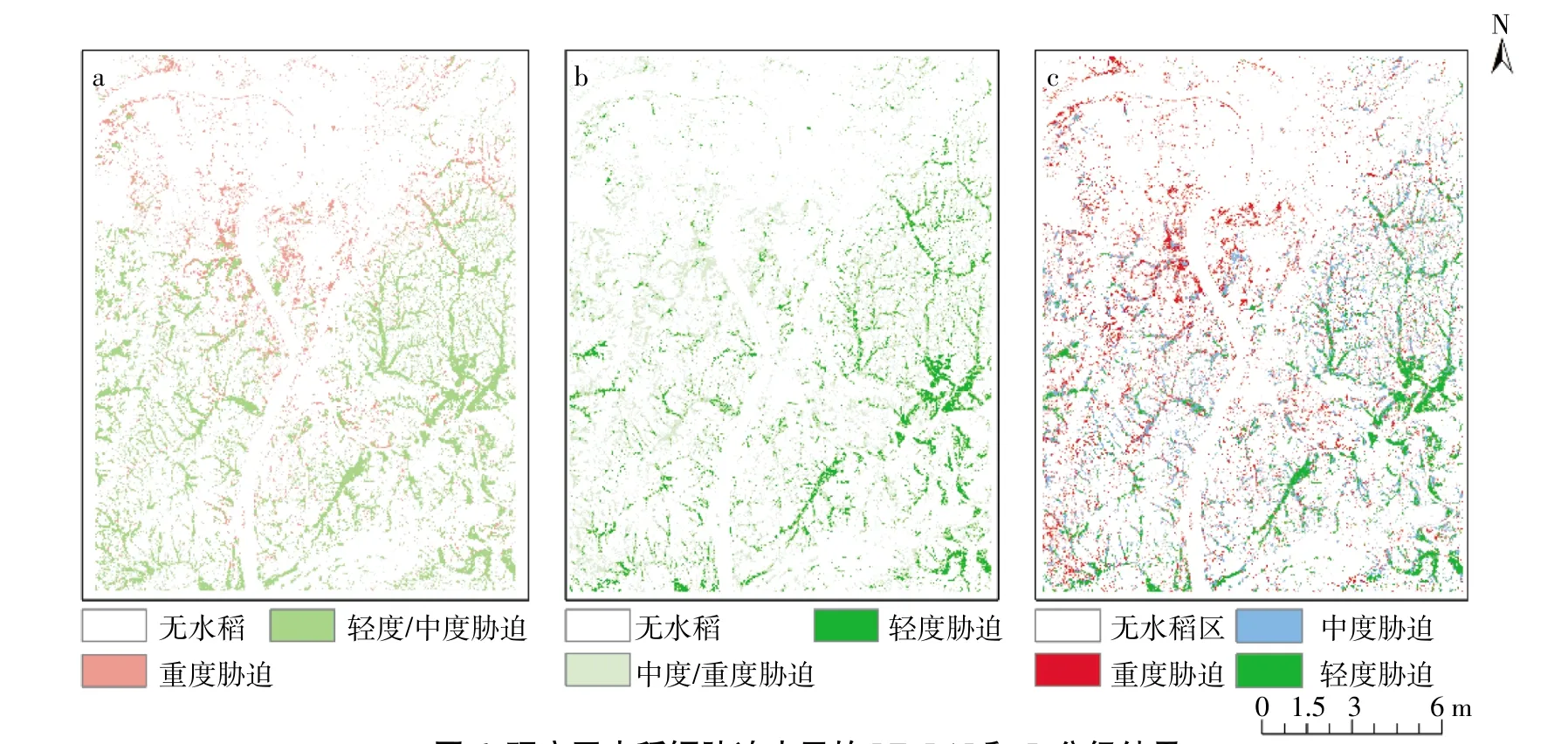
图6 研究区水稻镉胁迫水平的LE、LAI和Pf分级结果Figure 6 Cadmium stress levelof rice estimated by LE,LAIand Pf
本研究提供了一种结合遥感热红外、可见光和近红外从水稻生理功能的角度分析水稻镉污染胁迫的新思路。研究中温度和LAI的反演误差主要来源于遥感数据和反演方法,后续研究中将从遥感反演方法和数据方面研究如何提高温度与LAI反演精度。本文研究方法适合于环境因素差异不大的水稻镉胁迫监测,在自然条件下,环境因素对水稻生理功能的影响较大,环境因素与镉胁迫对水稻生理功能的影响关系还需进一步研究。
4 结论
(1)镉胁迫会抑制水稻蒸腾作用,水稻生理功能胁迫指数Pf能较好区分不同镉胁迫,其对轻度、中度和重度镉胁迫区分的准确度分别为70.59%、69.05%和84.79%。
(2)研究区内重度镉污染区域主要分布在湘江附近,随着与湘江距离变大,水稻由重度镉污染渐变为轻度镉污染,说明镉的污染源主要来自湘江,符合镉污染分布规律。
[1]张军,束文圣.植物对重金属镉的耐受机制[J].植物生理与分子生物学学报,2006,32(1):1-8.
ZHANG Jun,SHUWen-sheng.Studies on the tolerancemechanism of cadmium to plants[J].Journal of Plant Physiology and Molecular Biology,2006,32(1):1-8.
[2]赵利婷,刘湘南,丁超,等.遥感同化WOFOST模型动态监测水稻重金属污染胁迫[J].农业环境科学学报,2015,34(2):248-256.
ZHAO Li-ting,LIU Xiang-nan,DING Chao,et al.Study on dynamic monitoring of heavymetalpollution stress in ricebyWOFOSTmodel[J]. JournalofAgro-EnvironmentScience,2015,34(2):248-256.
[3]Liu M L,Li X N,Zhang B Y,et al.Regional heavymetal pollution in cropsby integrating physiological function variabilitywith,spatio-temporal stability usingmulti-temporal thermal remote sensing[J].International Journal of Applied Earth Observation&Geoinformation,2016, 51:91-102.
[4]凌军,张拴勤,潘家亮,等.植物蒸腾作用对红外辐射特征的影响研究[J].光谱学与光谱分析,2012,32(7):1775-1779.
LING Jun,ZHANG Shuan-qin,PAN Jia-liang,et al.Effects of plant transpiration on infrared radiation characteristics[J].Spectroscopy and SpectralAnalysis,2012,32(7):1775-1779.
[5]Moran M S,Clarke TR,Inoue Y,etal.Estimating crop water deficitusing the relation between surface-air temperatureand spectralvegetation index[J].Remote SensingofEnvironment,1994,49(3):246-263.
[6]陈咏淑,吴甫成,吕焕哲,等.近20年来湘江水质变化分析[J].长江流域资源与环境,2004,13(5):508-512.
CHEN Yong-shu,WU Fu-cheng,LÜHuan-zhe,etal.Analysisof Xiangjiang Riverwaterquality change in recent20 years[J].Resourcesand Environment in the Yangtze Basin,2004,13(5):508-512.
[7]Liang SL.Narrowband to broadband conversions of land surface albedo I:Algorithms[J].Remote Sensing of Environment,2001,76(2):213-238.
[8]Shuttleworth W J,Gurney R J.The theoretical relationship between foliage temperature and canopy resistance in sparse crops[J].Quarterly Journalof the RoyalMeteorologicalSociety,1990,116(492):497-519.
[9]季劲钧,胡玉春.一个植物冠层物理传输和生理生长过程的多层模式[J].气候与环境研究,1999,4(2):25-37.
JIJin-jun,HU Yu-chun.A Multilayermodel of plant canopy physical transportand physiological growth processes[J].Climatic and EnvironmentalResearch,1999,4(2):25-37.
[10]刘昌明.土壤-植物-大气系统水分运行的界面过程研究[J].地理学报,1997,52(4):366-373.
LIUChang-ming.Interfacialprocessesofsoil-plant-atmospherewatersystem[J].Acta Geographica Sinica,1997,52(4):366-373.
[11]BastiaanssenW GM,MenentiM,Feddes R A,etal.A remote sensing surface energy balance algorithm for land(SEBAL):1.Formulation[J]. JournalofHydrology,1998,212/213(1/2/3/4):198-212.
[12]Brutsaert W.Evaporation into the atmosphere[M].Netherlands:Springer,1982.
[13]Businger JA.A noteon the businger-dyer profiles[J].Boundary-Layer Meteorology,1988,42(1):145-151.
[14]Paulson CA.Themathematical representation ofwind speed and temperature profiles in theunstableatmospheric surface layer[J].Journalof Applied Meteorology,1987,9(9):857-861.
[15]FriedlM A.Modeling land surface fluxesusing a sparse canopymodel and radiometric surface temperaturemeasurements[J].Journal of GeophysicalResearch Atmospheres,1995,1002(D12):25435-25446.
[16]Jimenez-Munoz JC,Cristobal J,Sobrino JA,etal.Revision of the single-channel algorithm for land surface temperature retrieval from landsatthermal-infrared data[J].IEEETransactionson Geoscience&Remote Sensing,2009,47(1):339-349.
[17]Jimenez-Munoz JC,Sobrino JA,Skokovic D,etal.Land surface temperature retrievalmethods from landsat-8 thermal infrared sensor data [J].IEEEGeoscience&Remote Sensing Letters,2014,11(10):1840-1843.
[18]覃志豪,LIWen-juan,ZHANGMing-hua,等.单窗算法的大气参数估计方法[J].国土资源遥感,2003,15(2):37-43.
QIN Zhi-hao,LIWen-juan,ZHANG Ming-hua,et al.Optimization method ofsingle-channelalgorithm for atmospheric parameters[J].Remote Sensingof Land and Resources,2003,15(2):37-42.
[19]Wang FM,Huang JF,Tang Y L,et al.New vegetation index and its application in estimating leafarea index of rice[J].Rice Science,2007, 14(3):195-203.
Cadm ium stressanalysisbased on the changes in latent heat flux and canopy structuralparam eter of rice using remotely sensed data
LUO Li-bing,LIUXiang-nan*,WU Ling,LIUMei-ling
(Schoolof Information Engineering,China University ofGeosciences(Beijing),Beijing100083,China)
A physiological function sensitivity index(Pf),created by combining the rice canopy energy balancemodeland the structural parameter Leaf Area Index(LAI),is proposed in order to analyze the cadmium stress levelof rice.Firstly,the latentheat flux(LE)of rice was simulated based on an energy balancemodel using remote sensing and in situmeasured data,and then the Pf,which can reflect the physiological function of rice,was developed using LE and the LAI.Finally,the cadmium stress in rice was evaluated using the Pf.The results show thathigher cadmium stress generally corresponds to smaller valuesof LE,the LAI,and the Pf.LE could only distinguish high cadmium stress from other levels(medium/low)and the LAIcould only discriminate high andmedium cadmium stress from low stress;however,the Pfcould differentiate the three levels of cadmium stress of rice.The classification accuracies of Pffor high,medium,and low stress were 70.59%,69.05%,and 84.79%,respectively.Pfcan effectively discriminate different cadmium stress levels;therefore,it is an important reference formonitoring heavymetalstressof crops.
cadmium stress;energy balance;latentheat flux;LAI;stress index
S511
A
1672-2043(2017)05-0832-07
10.11654/jaes.2016-1583
2016-12-10
罗力兵(1989—),男,贵州铜仁人,硕士研究生,主要从事遥感信息分析与地学应用研究。E-mail:cugbllb@163.com
*通信作者:刘湘南E-mail:liuxn@cugb.edu.cn
国家自然科学基金项目(41371407);中央高校基本科研业务费专项资金(2-9-2015-107)
Project supported:The National Natural Science Foundation of China(41371407);The Fundamental Research Funds for the Central Universities(2-9-2015-107)
罗力兵,刘湘南,吴伶,等.镉胁迫水稻潜热通量与冠层结构参数变化遥感分析[J].农业环境科学学报,2017,36(5):832-838.
LUO Li-bing,LIU Xiang-nan,WU Ling,etal.Cadmium stressanalysisbased on the changes in latentheat flux and canopy structuralparameter of rice using remotely sensed data[J].Journalof Agro-EnvironmentScience,2017,36(5):832-838.
