落羽杉与墨西哥落羽杉3个杂交后代品系的生长和杂种优势分析
2017-06-05王丹碧凌子然胡李娟殷云龙王中生
王丹碧, 余 华, 凌子然, 胡李娟, 殷云龙, 王中生,①
〔1. 南京大学: a. 生命科学学院, b. 国际地球系统科学研究所, 江苏 南京 210023; 2. 江苏省中国科学院植物研究所(南京中山植物园), 江苏 南京 210014〕
落羽杉与墨西哥落羽杉3个杂交后代品系的生长和杂种优势分析
王丹碧1a, 余 华1b, 凌子然1a, 胡李娟2, 殷云龙2, 王中生1a,①
〔1. 南京大学: a. 生命科学学院, b. 国际地球系统科学研究所, 江苏 南京 210023; 2. 江苏省中国科学院植物研究所(南京中山植物园), 江苏 南京 210014〕
以落羽杉〔Taxodiumdistichum(Linn.) Rich.〕和墨西哥落羽杉(T.mucronatumTenore)及其3个杂交后代品系‘中山杉405’(‘Zhongshansha 405’)、‘中山杉406’(‘Zhongshansha 406’)和‘中山杉407’(‘Zhongshansha 407’)2年生盆栽扦插苗为研究对象,对株高和地径的年增长量、总叶面积、株高和地径的增长量动态变化、株高和地径的生长期进行了比较分析,并根据地径增长量分析了3个杂交后代品系的杂种优势。结果表明:3个杂交后代品系的年株高增长量介于父本与母本之间但无显著差异,而三者的年地径增长量和总叶面积则明显高于父本和母本。墨西哥落羽杉与3个杂交后代品系的株高和地径生长期均一致,分别为5月11日至10月5日和6月1日至11月9日;而落羽杉的株高和地径生长期分别为5月11日至8月10日和6月8日至11月16日。5月份至10月份,墨西哥落羽杉的株高持续增长,而3个杂交后代品系的株高不连续增长,落羽杉的株高则在8月10日之后停止生长;生长初期和生长末期,3个杂交后代品系的地径增长量均高于父本和母本;秋季3个杂交后代品系均能快速进行地径生长,而此时父本和母本的地径增长量均下降。除夏季的少数时间段外,‘中山杉405’在整个生长季都表现出明显的杂种优势,而‘中山杉406’和‘中山杉407’在整个生长季总体上表现出杂种优势,且它们的杂种优势最大值均出现在生长后期,说明供试的3个杂交后代品系在秋季也能快速生长。研究结果显示:供试的3个杂交后代品系均遗传了落羽杉地径生长迅速和墨西哥落羽杉株高生长期长的特性,秋季3个杂交后代品系的地径和株高增长量总体上均高于父本和母本,表现出明显的杂种优势。
落羽杉; 墨西哥落羽杉; ‘中山杉’; 株高; 地径; 杂种优势
生长是作物产量的决定因子。树木的生长可以定义为时间序列上植株量度的增加[1],最常用的量度指标为树高和树径(地径或胸径),这些指标较易测量且与材积、生物量密切相关。这些指标在单位时间上的变化(即生长速率)可以表述为相对生长速率或者绝对生长速率。相对生长速率(即单位时间某量度指标相对于其最初值的变化)常用于研究草本植物,但不适用于树木,原因是成长过程中非光合器官(树干、树枝和根)占植株总生物量的比例逐渐增加,树木的相对生长速率会随着树木的成长而快速递减[2]。林学和生态学中的生长预测模型大多数是基于绝对生长速率[3],绝对生长速率为前后2次采样期间树木某量度指标的变化[4]。树木生长是许多复杂的生理、形态和生物量分配过程综合作用的最终结果,这些过程受遗传、发育和环境因子的共同影响。研究树木的生长速率不仅可以在个体层次上量化其生长表现和生长策略,还能了解植株整体对环境的适应性[5]。
杂种优势普遍存在于生物界,是指杂交后代在生长、繁殖、适应性或者产量等方面优于亲本的中值或2个亲本的现象[6]。利用杂种优势原理培育林木优良品种已在杨属(PopulusLinn.)、落叶松属(LarixMill.)和桉属(EucalyptusL’Her.)等属的树种中广泛应用[6-8];例如,日本落叶松〔Larixkaempferi(Lamb.) Carr.〕与长白落叶松(L.olgensisHenry)的杂种生长超亲优势明显[8]。落羽杉属(TaxodiumRich.)杂交育种为中国首创,已有近50年的历史[9]9-10,[10-11],积累了丰富的品种。‘中山杉’(Taxodiumhybrid ‘Zhongshansha’)是江苏省中国科学院植物研究所从落羽杉〔T.distichum(Linn.) Rich.〕与墨西哥落羽杉(T.mucronatumTenore)杂交组合中选育出来的优良无性系,具有速生、耐盐碱和耐水湿等特点[11]。近年来,‘中山杉’在云南滇池、安徽巢湖和三峡库区等地的湿地生态系统恢复工程中发挥了巨大作用[12-14]。虽然落羽杉属植物的育种和推广已经取得许多阶段性成果,但相关研究目前主要集中在造林以及耐盐碱、耐淹水和外部形态变异等方面[10-15],对于亲本和杂交后代的生长速率、节律以及杂种生长优势等方面还缺乏必要的了解。
作者以生长在相同环境中的同龄落羽杉和墨西哥落羽杉及其3个杂交后代品系为研究对象,通过在整个生长季对株高生长和地径生长的连续测量,试图重建株高和地径生长轨迹,量化生长初期、快速生长期和生长末期的株高和地径的增长量,并计算杂交后代的杂种优势;在此基础上,比较3个杂交后代品系和亲本的生长策略和节律,以期分析和总结3个杂交后代品系速生丰产超双亲的基础。
1 材料和方法
1.1 材料
实验地点为江苏省中国科学院植物研究所(地理坐标为东经118°48′、北纬32°07′)的露天苗圃。以父本落羽杉和母本墨西哥落羽杉及其3个杂交后代品系‘中山杉405’(‘Zhongshansha 405’)、‘中山杉406’(‘Zhongshansha 406’)和‘中山杉407’(‘Zhongshansha 407’)为研究对象。于2010年4月中旬将亲本及其3个杂交后代品系的2年生扦插苗移入塑料盆(上口径30 cm、下口径20 cm、高度25 cm)中定植,每盆1株。每个亲本及杂交后代品系16~20盆。根据植株生长状态和天气情况定期定量浇水施肥。
1.2 方法
1.2.1 株高和地径的测量 分别于2010年5月4日、5月18日、6月1日、6月15日、6月29日、7月13日、7月27日、8月10日、8月24日、9月7日、9月21日、10月5日、10月19日、11月2日和11月16日,选择同一个时段,用不锈钢卷尺(精度0.1 cm)测量植株的株高,用游标卡尺(精度0.01 mm)测量地径。为了更准确地确定株高和地径生长的开始期和结束期,分别在5月11日、11月9日和11月23日增加3次观测。为保证地径测量位置的一致,首次测量时用记号笔标定地径测量的位置,在每一样株的3个不同方向测量地径,结果取平均值。
1.2.2 植株总叶面积的测量 由于植株总叶面积较大,每个亲本及杂交后代品系各随机选3株样株,用子样法[16]测量样株的比叶面积(specific leaf area,SLA)。于2010年11月30日,在样株的上、中、下层各取1个小枝,每个小枝50~100片小叶,用叶形纸称重法[17]测量小枝上所有样叶的总叶面积,然后将样叶和该植株剩余的所有叶片分别置于80 ℃烘箱中烘干7 d,根据样叶的总叶面积与烘干后质量的比值计算样叶的SLA,然后依据每个样株的叶片总质量与SLA的乘积估算植株的总叶面积。
1.3 数据计算和统计分析
计算每个亲本及杂交后代品系样株株高和地径各次测量值的平均值,然后计算前后2次测量值的变化值(即前后2次测量值的差值);逐次累加平均株高或平均地径的变化值,获得年株高增长量或年地径增长量。
将前后2次平均株高和平均地径出现变化的日期确定为生长的开始期,将前后2次平均株高和平均地径停止变化的日期确定为生长的结束期。由于树木的收缩以及测量误差,变化值出现负值时亦记为停止变化,记为“0”。
各树种的株高(地径)增长量根据公式“株高(地径)增长量=2周后的平均株高(平均地径)-2周前的平均株高(平均地径)”[4]计算。然后,根据地径增长量计算杂交后代品系的杂种优势,计算公式为“杂种优势=〔(杂交后代品系的地径增长量均值-亲本的地径增长量均值)/亲本的地径增长量均值〕×100%”。
使用SPSS 16.0统计分析软件对相关实验数据进行统计和分析,通过协方差分析(analysis of covariance,ANCOVA)消除落羽杉和墨西哥落羽杉及其3个杂交后代品系的初始地径值对地径增长的影响;采用Duncan’s新复极差法检验亲本与3个杂交后代品系的年株高增长量和年地径增长量的差异显著性。
2 结果和分析
2.1 地径增长量的协方差分析
落羽杉和墨西哥落羽杉及其3个杂交后代品系地径增长量的协方差分析结果见表1。由表1可以看出:初始地径的均方与初始地径和地径增长量相互作用的均方接近,而地径增长量的均方最小(0.017),说明初始地径以及初始地径和地径增长量相互作用对地径增长量的影响稍大,但差异未达到显著水平(P>0.05)。
表1 落羽杉和墨西哥落羽杉及其3个杂交后代品系地径增长量的协方差分析结果
Table 1 Result of analysis of covariance on basal diameter increment ofTaxodiumdistichum(Linn.) Rich.,T.mucronatumTenore and their three hybrid progeny lines

变异来源Sourceofvariation自由度Degreeoffreedom平方和Sumofsquares均方MeansquareP值Pvalue初始地径Initialbasaldiameter40.2230.0560.642地径增长量Basaldiameterincrement10.0170.0170.661初始地径×地径增长量Initialbasaldiameter×basaldiameterincrement40.2120.0530.663误差Error827.2360.088总计Total917.688
2.2 年株高增长量、年地径增长量和总叶面积的比较分析
落羽杉和墨西哥落羽杉及其3个杂交后代品系‘中山杉405’、‘中山杉406’和‘中山杉407’的年株高增长量、年地径增长量和总叶面积的比较结果见表2。
由表2可见:墨西哥落羽杉的年株高增长量(34.44 cm)高于落羽杉的年株高增长量(25.33 cm),而‘中山杉405’、‘中山杉406’和‘中山杉407’的年株高增长量分别为25.85、26.81和28.86 cm,介于父本和母本之间,但各亲本及其杂交后代品系间的年株高增长量无显著差异(P>0.05)。
由表2还可见:‘中山杉407’的年地径增长量最大,为15.02 mm,墨西哥落羽杉最小,为11.88 mm,且3个杂交后代品系的年地径增长量均高于亲本,其中,‘中山杉406’和‘中山杉407’的年地径增长量显著高于亲本(P≤0.05),而‘中山杉405’的年地径增长量显著高于墨西哥落羽杉,但与落羽杉无显著差异。3个杂交后代品系保持了落羽杉地径快速增长和干形通直的优良特性。落羽杉的年地径增长量显著高于墨西哥落羽杉,说明落羽杉地径的生长优于墨西哥落羽杉。
另外,3个杂交后代品系的总叶面积均大于落羽杉和墨西哥落羽杉,其中,‘中山杉406’的总叶面积最大,为0.89 m2,而墨西哥落羽杉的总叶面积最小,为0.49 m2,二者间差异达显著水平,而双亲与另2个杂交后代品系的总叶面积的差异则均不显著。
相关性分析结果表明:落羽杉和墨西哥落羽杉及其3个杂交后代品系的年地径增长量与总叶面积呈正相关,相关系数为0.86。



树种Treespecies年株高增长量/cmAnnualheightincrement年地径增长量/mmAnnualbasaldiameterincrement总叶面积/m2Totalleafarea中山杉405Zhongshansha40525.85±2.21a15.02±0.72b0.80±0.19ab中山杉406Zhongshansha40626.81±2.42a17.79±0.77a0.89±0.08a中山杉407Zhongshansha40728.86±2.18a17.93±0.71a0.70±0.08ab墨西哥落羽杉T.mucronatum34.44±2.19a11.88±0.70c0.49±0.09b落羽杉T.distichum25.33±2.62a14.23±0.51b0.54±0.04ab
1)同列中不同的小写字母表示差异显著(P≤0.05) Different lowercases in the same column indicate the significant difference (P≤0.05).
2.3 株高和地径生长期的比较
落羽杉和墨西哥落羽杉及其3个杂交后代品系的株高和地径生长期的比较结果见表3。
由表3可见:落羽杉和墨西哥落羽杉及其3个杂交后代品系的株高生长期均从5月11日开始,而仅落羽杉的株高生长期于8月10日结束,墨西哥落羽杉及3个杂交后代品系的株高生长期均于10月5日结束,较落羽杉的株高生长期长约2个月。
表3 落羽杉和墨西哥落羽杉及其3个杂交后代品系的株高和地径生长期的比较
Table 3 Comparison on growth periods of height and basal diameter ofTaxodiumdistichum(Linn.) Rich.,T.mucronatumTenore and their three hybrid progeny lines

树种Treespecies生长期(MM-DD)Growthperiod(MM-DD)株高Height地径Basaldiameter中山杉405Zhongshansha40505-11—10-0506-01—11-09中山杉406Zhongshansha40605-11—10-0506-01—11-09中山杉407Zhongshansha40705-11—10-0506-01—11-09墨西哥落羽杉T.mucronatum05-11—10-0506-01—11-09落羽杉T.distichum05-11—08-1006-08—11-16
从表3还可见:墨西哥落羽杉及3个杂交后代品系的地径生长期均于6月1日开始、11月9日结束,仅落羽杉的地径生长期开始时间和结束时间均较晚,于6月8日开始、11月16日结束。
2.4 株高增长量的动态变化
落羽杉和墨西哥落羽杉及其3个杂交后代品系株高增长量的动态变化见图1。由图1可见:5月份至10月份墨西哥落羽杉的株高持续增长,而3个杂交后代品系的株高则不连续增长。
8月10日之后落羽杉的株高生长停止,7月27日其株高增长量最低,仅0.36 cm。落羽杉在6月15日和8月10日的株高增长量较大,分别为5.28和5.56 cm。7月高温对墨西哥落羽杉的株高生长有明显影响,导致其株高增长量最低,7月27日其株高增长量仅0.38 cm,与其株高生长趋于停止时(10月19日)的株高增长量(0.31 cm)接近;此外,墨西哥落羽杉在各时期的株高增长量均较大,最大值(5.19 cm)出现在10月5日。‘中山杉405’在6月15日和9月21日的株高增长量较大,分别为3.40和3.35 cm。‘中山杉406’和‘中山杉407’的株高增长量最大值分别出现在6月15日和6月29日,分别为4.97和5.30 cm。
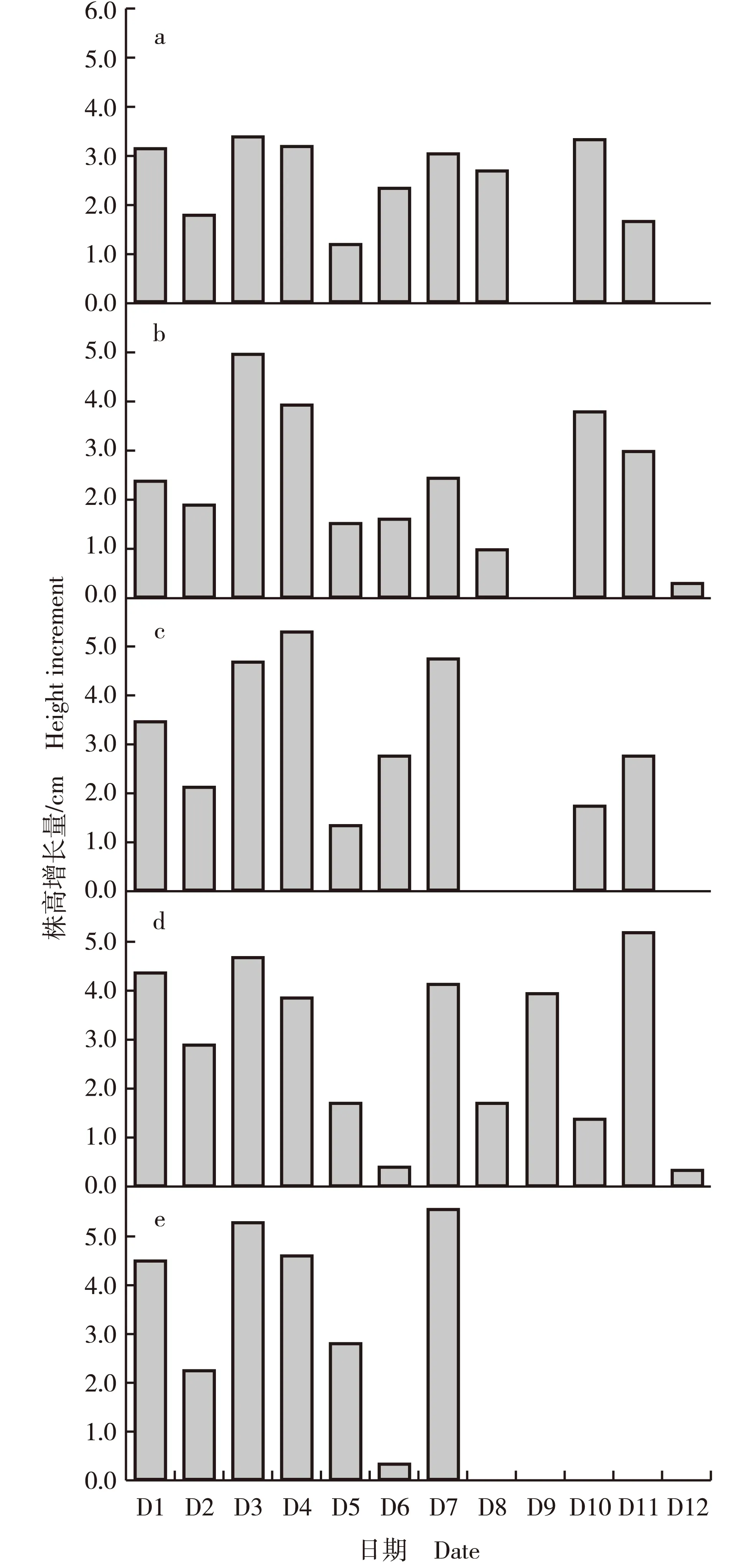
D1: 5月18日May 18th; D2: 6月1日June 1st; D3: 6月15日June 15th; D4: 6月29日June 29th; D5: 7月13日July 13th; D6: 7月27日July 27th; D7: 8月10日August 10th; D8: 8月24日August 24th; D9: 9月7日September 7th; D10: 9月21日September 21st; D11: 10月5日October 5th; D12: 10月19日October 19th.a: ‘中山杉405’‘Zhongshansha 405’; b: ‘中山杉406’‘Zhongshansha 406’; c: ‘中山杉407’‘Zhongshansha 407’; d: 墨西哥落羽杉T. mucronatum Tenore; e: 落羽杉T. distichum (Linn.) Rich.图1 落羽杉和墨西哥落羽杉及其3个杂交后代品系株高增长量的动态变化Fig. 1 Dynamic change in height increment of Taxodium distichum (Linn.) Rich., T. mucronatum Tenore and their three hybrid progeny lines
5月18日,落羽杉和墨西哥落羽杉的株高增长量分别为4.47和4.34 cm,而3个杂交后代品系的株高增长量为2.39~3.45 cm,落羽杉和墨西哥落羽杉的株高增长量均大于3个杂交后代品系。7月27日,落羽杉和墨西哥落羽杉的株高增长量分别为0.36和0.38 cm,而‘中山杉405’、‘中山杉406’和‘中山杉407’的株高增长量分别为2.35、1.61和2.73 cm,说明7月高温对落羽杉和墨西哥落羽杉株高生长的影响大于对3个杂交后代品系的影响。8月10日以后,落羽杉的株高生长停止,墨西哥落羽杉的株高持续旺盛生长,例如,10月5日墨西哥落羽杉的株高增长量达5.19 cm;与墨西哥落羽杉相似,秋季3个杂交后代品系的株高生长旺盛,例如,9月21日‘中山杉405’、‘中山杉406’和‘中山杉407’的株高增长量分别为3.35、3.78和1.75 cm,10月5日3个杂交后代品系的株高增长量分别为1.65、3.00和2.76 cm。
2.5 地径增长量的动态变化
落羽杉和墨西哥落羽杉及其3个杂交后代品系地径增长量的动态变化见图2。
由图2可以看出:生长初期,‘中山杉405’、‘中山杉406’和‘中山杉407’的地径增长量均大于落羽杉和墨西哥落羽杉,例如,6月份3个杂交后代品系的地径增长量总体上大于落羽杉和墨西哥落羽杉。但随生长期的延续,亲本和3个杂交后代品系的地径增长量变化趋势发生改变。7月13日,‘中山杉406’的地径增长量大于墨西哥落羽杉,而‘中山杉407’的地径增长量则大于双亲;7月27日,墨西哥落羽杉的地径增长量最小,其次为‘中山杉405’,说明7月高温对二者的地径生长影响较大;8月10日,双亲和3个杂交后代品系的地径增长量接近,均略低于或等于1 mm;8月24日,3个杂交后代品系的地径增长量均大于双亲。

D1: 6月1日June 1st; D2: 6月15日June 15th; D3: 6月29日June 29th; D4: 7月13日July 13th; D5: 7月27日July 27th; D6: 8月10日August 10th; D7: 8月24日August 24th; D8: 9月7日September 7th; D9: 9月21日September 21st; D10: 10月5日October 5th; D11: 10月19日October 19th; D12: 11月2日November 2nd.a: ‘中山杉405’‘Zhongshansha 405’; b: ‘中山杉406’‘Zhongshansha 406’; c: ‘中山杉407’‘Zhongshansha 407’; d: 墨西哥落羽杉T. mucronatum Tenore; e: 落羽杉T. distichum (Linn.) Rich.图2 落羽杉和墨西哥落羽杉及其3个杂交后代品系地径增长量的动态变化Fig. 2 Dynamic change in basal diameter increment of Taxodium distichum (Linn.) Rich., T. mucronatum Tenore and their three hybrid progeny lines
进入地径快速生长期(9月份)之后,落羽杉和墨西哥落羽杉与‘中山杉405’、‘中山杉406’和‘中山杉407’的地径增长量的差异增大,例如,9月7日和9月21日,‘中山杉405’的地径增长量相近,分别为2.26和2.23 mm;9月21日,‘中山杉406’和‘中山杉407’的地径增长量分别高达3.51和3.13 mm,大于落羽杉的地径增长量(2.68 mm),更远大于墨西哥落羽杉的地径增长量(1.62 mm);10月份,3个杂交后代品系的地径增长量接近于7月份,但双亲的地径增长量均低于7月份。说明即使在秋季,3个杂交后代品系的地径也能快速生长,而此时双亲的地径增长量均下降。10月份和11月份,落羽杉的地径增长量较小,说明此阶段其地径生长较慢,这也是落叶树种对逐渐降低的气温的适应性表现之一。
6月28日至10月5日,落羽杉的地径增长量均大于墨西哥落羽杉,其中,9月7日、9月21日和10月5日,落羽杉的地径增长量分别比墨西哥落羽杉高53.8%、65.4%和70.0%。说明在8月10日株高生长停止之后,落羽杉将资源更集中地分配在地径生长上,并且其地径生长优于墨西哥落羽杉。
2.6 杂交后代的杂种优势比较
在树木的相对生长方程(allometric equation)中,地径增长对生物量的贡献最大,因此地径增长常用于指示树木整株的生长[1]。以地径增长量为指标,对‘中山杉405’、‘中山杉406’和‘中山杉407’的杂种优势进行分析,结果见表4。
由表4可见:总体上看,3个杂交后代品系在整个生长季均表现出杂种优势。除6月15日、7月13日和7月27日外,‘中山杉405’在其他时期都表现出明显的杂种优势,其中,在快速生长期(9月7日)其杂种优势为35.3%,在生长末期(10月19日和11月2日)其杂种优势明显,分别达到44.8%和95.9%。除7月13日外,‘中山杉406’在整个生长季均表现出杂种优势,在生长初期(6月15日)其杂种优势相对较低,为16.9%;在高温期(7月27日)其杂种优势升至36.7%;在快速生长期其杂种优势更明显,9月7日、9月21日和10月5日其杂种优势分别为61.0%、63.3%和70.2%,10月19日其杂种优势高达72.4%。与‘中山杉406’相似,除8月10日外,‘中山杉407’在整个生长季均表现出杂种优势,在生长初期其杂种优势略小,但在快速生长期其杂种优势明显,8月24日、9月7日和9月21日其杂种优
表4 落羽杉和墨西哥落羽杉的3个杂交后代品系杂种优势的动态变化势分别为50.8%、61.6%和45.6%;10月19日‘中山杉407’的杂种优势最大,达113.0%;即使在生长末期(11月2日)其杂种优势还保持在45.2%。
Table 4 Dynamic change in heterosis of three hybrid progeny lines fromTaxodiumdistichum(Linn.) Rich. andT.mucronatumTenore

树种Treespecies不同日期(MM-DD)的杂种优势/% Heterosisatdifferentdates(MM-DD)06-1506-2907-1307-2708-1008-2409-0709-2110-0510-1911-02中山杉405Zhongshansha405-9.453.1-30.0-3.29.611.435.33.515.344.895.9中山杉406Zhongshansha40616.912.9-3.936.73.96.761.063.370.272.438.1中山杉407Zhongshansha40711.241.113.521.0-0.150.861.645.643.2113.045.2
研究结果显示:3个杂交后代品系最大的杂种优势均在生长末期,推测由于该时期亲本的地径增长量都很低,特别是落羽杉为落叶性树种,10月中下旬树叶衰老变黄,地径生长趋于停止。而进入10月份后,3个杂交后代品系继续保持着不低于7月份的地径增长量,而此阶段无论是墨西哥落羽杉还是落羽杉的地径增长量均低于7月份。说明3个杂交后代品系在秋季也能继续进行快速生长,表现出持续的生长优势。
3 讨论和结论
落羽杉和墨西哥落羽杉的株高和地径增长量差异较大,这与落羽杉为落叶性树种,而墨西哥落羽杉为半常绿性树种有关。本研究结果显示:生长在中国江苏南京的墨西哥落羽杉还保持着株高生长连续及株高生长期延长至秋季等特性,这与其原产地(美国德克萨斯州西南部、墨西哥和危地马拉[9]2)的热带气候特征相吻合。供试的3个杂交后代品系均遗传了墨西哥落羽杉株高生长期长的特性,秋季还能进行旺盛的株高生长。南京每年6月下旬到7月中旬为梅雨季节,之后即进入高温期,且南京夏季高温较同纬度其他地区高[18]。半常绿性的墨西哥落羽杉的优势为叶片功能期长,但在本研究中,墨西哥落羽杉最不耐南京的夏季高温,表现为其株高和地径增长量在7月中下旬最低,推测原因为墨西哥落羽杉的原产地为河谷或河岸带植被,自然状态下几乎全部分布在河岸、溪流或泉水边,偶尔也分布于有浅层地下水的平原地区[9]2。本研究的供试苗木均为盆植,花盆较大的表面积与体积之比及容器表面吸收太阳辐射均会导致盆内土壤温度以4 ℃·h-1~10 ℃·h-1的速率升高[19],盆内土壤累积的高温可能损伤根系并阻滞生长,尤其是7月下旬午间气温高达40.55 ℃[20]23,因此,墨西哥落羽杉根系不适应高温,表现出最低的生长速率。此外,墨西哥落羽杉的地径增长量在整个生长季均没有出现明显高峰,呈现平缓的生长节奏,这也是其对原产地热带气候的适应性表现之一。落羽杉为落叶性树种,整个生长季其株高和地径增长呈现明显峰值,尤其是8月10日株高生长停止以后,光合同化产物较为集中分配至地径生长,出现了地径增长量的峰值,甚至在9月21日略高于杂交后代品系‘中山杉405’,更较墨西哥落羽杉高65%。供试3个杂交后代品系的地径增长也继承了其父本落羽杉快速生长的特性,例如,9月21日,‘中山杉406’的地径增长量高达3.51 mm。与此相比,同为落叶树种的华北落叶松(Larixprincipis-rupprechtiiMayr)当年播种苗的地径月净生长量峰值只接近0.50 mm[21],说明落羽杉和墨西哥落羽杉的3个杂交后代品系对光能和热量等资源的利用更佳。
本研究中,供试的3个杂交后代品系在耐高温、总叶面积、地径生长、株高生长和生长期等方面具有杂种优势。3个杂交后代品系植株的总叶面积均大于双亲,因而能够截获更多的光能,使单株光合效率提高,杂种优势通过总叶面积的变化直接影响冠层的光合作用。此外,生长期是决定植物总生长量的重要因子。3个杂交后代品系的株高生长期比父本落羽杉长近2个月。胡李娟等[20]17-18的研究结果表明:进入10月份之后,‘中山杉405’、‘中山杉406’和‘中山杉407’继续保持着与9月份(光合高峰期)同等的光合速率水平,促进了这3个杂交后代品系在快速生长期和生长末期的干物质积累,并为此时期显著的株高生长和地径生长提供了物质基础。此外,3个杂交后代品系于生长末期还能旺盛生长,说明这3个杂交后代品系充分利用了延长的生长季,这也是三者表现出杂种优势的原因之一。落叶松〔Larixgmelinii(Rupr.) Kuzen.〕杂交后代的生长期也比亲本长[22],因此,推测生长期延长也是供试3个杂交后代品系的株高和地径大于双亲的基础原因之一。
综合分析结果表明:供试的3个杂交后代品系结合了父本落羽杉和母本墨西哥落羽杉的生长特性,既继承了落羽杉地径扩展生长迅速的特性,也保持了墨西哥落羽杉生长期长的特性,在生长末期还能保持旺盛生长,因此,3个杂交后代品系的株高和地径生长均超过亲本,表现出明显的杂种优势。
[1] BOWMAN D M J S, BRIENEN R J W, GLOOR E, et al. Detecting trends in tree growth: not so simple[J]. Trends in Plant Science, 2013, 18: 11-17.
[2] MENCUCCINI M, MARTNEZ-VILATA J, VANDERKLEIN D, et al. Size-mediated ageing reduces vigor in trees[J]. Ecology Letters, 2005, 10: 1183-1190.
[3] LAROCQUE G R, MARSHALL P L. Evaluating the impact of competition using relative growth rate in red pine (PinusresinosaAit.) stands[J]. Forest Ecology and Management, 1993, 58: 65-83.
[4] POMMERENING A, MUSZTA A. Relative plant growth revisited: towards a mathematical standardization of separate approaches[J]. Ecological Modeling, 2016, 320: 383-392.
[5] YU H. Fast growing ofAcaciamangium: a result of efficient light utilization and carbon assimilation[D]. Singapore: School of Biological Sciences, National University of Singapore, 2002.
[6] MARRON N, CEULEMANS R. Genetic variation of leaf traits related to productivity in aPopulusdeltoides×Populusnigrafamily[J]. Canadian Journal of Forest Research, 2006, 36: 390-400.
[7] 叶金山, 王章荣. 杂种马褂木杂种优势的遗传分析[J]. 林业科学, 2002, 38(4): 67-71.
[8] 王景章, 丁振芳. 落叶松杂种优势及其利用的研究[J]. 遗传, 1989, 11(4): 1-4.
[9] 殷云龙, 於朝广. 中山杉: 落羽杉属树木杂交选育[M]. 中国林业出版社, 2005.
[10] 於朝广, 殷云龙. 落羽杉属树木种间杂交选育研究进展[J]. 江苏林业科技, 2008, 35(2): 39-46.
[11] 於朝广, 殷云龙, 徐建华. 落羽杉属4个新品种[J]. 林业科学, 2011, 47(5): 181-182.
[12] 韩亚平, 徐 杉, 朱 勇. 滇池湖滨湿地中山杉和黑杨的光合特性[J]. 林业科技开发, 2012, 26(2): 57-59.
[13] 马 林, 杨红明, 钟 华, 等. 中山杉引种研究进展及其在昆明地区的应用现状[J]. 林业调查规划, 2011, 36(1): 19-25.
[14] 殷云龙, 於朝广, 华建峰, 等. 重庆万州三峡库区消落带中山杉造林试验[J]. 林业科技开发, 2014, 28(2): 117-121.
[15] 杨美凌, 殷云龙, 方炎明, 等. 落羽杉属种类、栽培变种及杂种的外部形态变异及亲缘关系研究[J]. 植物资源与环境学报, 2010, 19(2): 40-47.
[16] WELLS R, MEREDITH W R, Jr, WILLIFORD J R. Heterosis in upland cotton. Ⅱ. Relationship of leaf area to plant photosynthesis[J]. Crop Science, 1988, 28: 522-525.
[17] YU H, ONG B L. Photosynthesis and antioxidant enzymes of phyllodes ofAcaciamangium[J]. Plant Science, 2000, 159: 107-115.
[18] 竺可桢. 南京之气候[M]. 南京: 东南大学出版社, 2014: 42.
[19] KOZLOWSKI T T, KRAMER P J, PALLARDY S G. The Physiological Ecology of Woody Plants[M]. San Diego: Academic Press, 1990: 208.
[20] 胡李娟. 杂交中山杉405、406、407及亲本的光合生理特性研究[D]. 南京: 江苏省中国科学院植物研究所, 2012.
[21] 董太祥. 华北落叶松播种苗当年生长规律的研究[J]. 河北农业大学学报, 1994, 17(1): 44-49.
[22] BALTUNIS B S, GREENWOOD M S. Variation in lateral shoots elongation patterns and hybrid vigor in full-sib families and interspecific hybrids of larch[J]. Tree Physiology, 1999, 19: 131-136.
(责任编辑: 张明霞)
Analyses on growth and heterosis of three hybrid progeny lines fromTaxodiumdistichumandT.mucronatum
WANG Danbi1a, YU Hua1b, LING Ziran1a, HU Lijuan2, YIN Yunlong2, WANG Zhongsheng1a,①
(1. Nanjing University: a. School of Life Sciences, b. International Institute for Earth System Science, Nanjing 210023, China; 2. Institute of Botany, Jiangsu Province and Chinese Academy of Sciences, Nanjing 210014, China),
J.PlantResour. &Environ., 2017, 26(1): 55-62
Taking two-year-old potted cutting seedlings ofTaxodiumdistichum(Linn.) Rich.,T.mucronatumTenore and their three hybrid progeny lines of ‘Zhongshansha 405’, ‘Zhongshansha 406’ and ‘Zhongshansha 407’ as research objects, annual increments of height and basal diameter, total leaf area, dynamic changes in increments of height and basal diameter, and growth periods of height and basal diameter were compared and analyzed, and heterosis of three hybrid progeny lines were analyzed according to basal diameter increment. The results show that annual height increments of three hybrid progeny lines are between male and female parents, but there is no significant difference, while their annual basal diameter increments and total leaf areas are obviously higher than those of male and female parents. Growth periods of height and basal diameter ofT.mucronatumare consistent with those of three hybrid progeny lines, which are from May 11th to October 5th and from June 1st to November 9th, respectively, while those ofT.distichumare from May 11th to August 10th and from June 8th to November 16th, respectively. From May to October, height ofT.mucronatumkeeps growing, while that of three hybrid progeny lines grows discontinuously, and that ofT.distichumstops growing after August 10th. During early and late growth periods, basal diameter increments of three hybrid progeny lines are higher than those of male and female parents. In autumn, basal diameter keeps increasing quickly in three hybrid progeny lines, while it decreases in male and female parents at the meantime. Except a few time segments in summer, ‘Zhongshansha 405’ exhibits obvious heterosis over the whole growing season. By contrast, ‘Zhongshansha 406’ and ‘Zhongshansha 407’ generally show heterosis over the whole growing season. For all three hybrid progeny lines, their highest heterosis values appear in late growth period, indicating that they also grow fast in autumn. This study shows that these three hybrid progeny lines tested inherit characteristics of fast growth of basal diameter ofT.distichumand long growth period of height ofT.mucronatum. In autumn, increments of both height and basal diameter of three hybrid progeny lines are totally higher than those of male and female parents, indicating obvious heterosis.
Taxodiumdistichum(Linn.) Rich.;TaxodiummucronatumTenore;Taxodiumhybrid ‘Zhongshansha’; height; basal diameter; heterosis
2016-05-02
江苏省创新能力建设计划(科技设施类)(BM2015019)
王丹碧(1992—),女,贵州黔南州人,硕士研究生,主要从事湿地生态学方面的研究。
①通信作者E-mail: wangzs@nju.edu.cn
S718.46; S791.34
A
1674-7895(2017)01-0055-08
10.3969/j.issn.1674-7895.2017.01.07
