缢江蓠生长率及其生化组分含量分析
2017-05-30王艳平谢恩义黄翔鹄孙立伟粟文王惠
王艳平 谢恩义 黄翔鹄 孙立伟 粟文 王惠

摘要:【目的】明确缢江蓠(Gracilaria salicornia)的最适宜生态条件,为其人工栽培提供参考依据。【方法】通过室内单因子试验设计,探讨不同温度、光照强度及盐度对缢江蓠生长率、叶绿素a(Chla)、类胡萝卜素(Car)、藻红蛋白(R-PE)、藻蓝蛋白(R-PC)、可溶性蛋白、超氧化物歧化酶(SOD)、丙二醛(MDA)变化的影响。【结果】缢江蓠的适宜生长温度为24~32 ℃,最适生长温度为28 ℃;适宜生长光照强度为3000~9000 lx,最适生长光照强度为6000 lx;适宜生长盐度为20‰~35‰,最适生长盐度为30‰。在适宜的环境条件下,缢江蓠藻体具有较高的生长率,其藻胆蛋白和可溶性蛋白含量较高,MDA含量较低,但SOD活性较高,即受环境胁迫较小,受氧化胁迫较大。【结论】缢江蓠具有能适应高温、低光照强度及中、高盐度环境条件的特性,可在我国南方海域夏季自然条件下栽培生产。
关键词: 缢江蓠;温度;光照强度;盐度;生长;生化组分
中图分类号: S968.434 文献标志码:A 文章编号:2095-1191(2017)06-1099-07
Abstract:【Objective】The most suitable ecological environment for Gracilaria salicornia growth was researched in order to provide reference for its artificial cultivation. 【Measure】Effects of temperature, light intensity and salinity on variation of growth rate, chlorophyll a(Chla), carotenoid(Car), phycoerythrin(R-PE), phycocyanin(R-PC), soluble protein, superoxide dismutase(SOD) and malondialdehyde(MDA) were analyzed by indoor single factor experiment. 【Result】The suitable growth temperature for G. salicornia was 24-32 ℃, and the optimum temperature was 28 ℃; the suitable light intensity was 3000-9000 lx, and the most suitable light intensity for growth was 6000 lx; the suitable salinity was 20‰-35‰ and the optimum salinity for growth was 30‰. Under the suitable conditions, growth rate of G. salicornia was high. And it contained high phycobiliprotein, soluble protein and SOD contents, low MDA content. It was inferred that G. salicornia suffered less from environment stree than oxidative stress. 【Conclusion】G. salicornia can adapt to the environment conditions of high temperature, low light intensity and medium-and-high salinity. Therefore, it can be cultured naturally in summer in the southern sea area of China.
Key words: Gracilaria salicornia; temperature; light intensity; salinity; growth; biochemical composition
\0 引言
【研究意义】缢江蓠(Gracilaria salicornia)隶属于红藻门(Rhodophyta)真红藻纲(Florideae)江蓠目(Gra-
cilariales)江蓠科(Gracilariaceae)江蓠属(Gracilaria),国内主要分布于广东、海南及台湾等地,国外分布于日本、美国等地,其形成的藻场能固定底质、改善水质,为许多动物提供栖息地、护幼场所和食物等,具有重要的生态功能(Phooprong et al.,2007;覃媚等,2016)。此外,缢江蓠是一种可适应高温条件生长的大型经济红藻,兼具食用、饲用和藻胶用等经济价值。随着我国琼胶制造工业的不断发展,琼胶原料的需求也逐年增长,因此发展大规模缢江蓠栽培不仅能为琼胶制造工业提供原料,还能净化沿海海域水质,修复生态环境。【前人研究进展】目前,关于江蓠属可栽培种类适宜生态条件等内容研究已有较多报道,如细基江蓠繁枝变种(Gracilaria tenuistipitata var. Liui)(陈昌生和章景荣,1999;许忠能等,2001)、菊花江蓠(G. lichenoides)(汤坤贤等,2005;徐永健等,2006)、龙须菜(G. lemaneiformis)(Yang et al.,2006;張学成等,2009;Wang et al.,2010;孙雪等,2013)、真江蓠(G. vermiculophylla)(Phooprong et al.,2008;霍元子等,2010)、脆江蓠(G. chouae)(金玉林等,2012;卢晓等,2014)、芋根江蓠(G. blodgettii)(黄中坚等,2014)、智利江蓠(G. chilensis)(陈伟洲等,2015)等,且一致认为温度、光照等物理因子和营养盐、盐度等化学因子及生物因子等是影响江蓠属海藻生长发育的关键环境因素,其研究结果为规模化栽培与生产江蓠提供了可靠的参考依据。【本研究切入点】探索最适合的江蓠生长生态条件是其产业化发展的基础和关键问题,但至今有关环境生态因子对缢江蓠生长等方面的研究尚无报道。【拟解决的关键问题】通过室内单因子试验设计,探讨不同温度、光照及盐度对缢江蓠生长率及生化组分含量的影响,明确缢江蓠的最适宜生态条件,为其人工栽培提供参考依据。
1 材料与方法
1. 1 试验材料
试验所用缢江蓠于2015年4月采自广东省湛江市流沙湾(东经109°56′19.3″,北纬20°26′2.0″),采集的鲜藻放入加冰泡沫箱帶回实验室,将藻体表面的泥沙和杂质以过滤海水清洗干净,再用过滤灭菌的自然海水暂养3 d后开展相关试验。预培养条件:海水温度28 ℃,盐度30‰,光照强度6000 lx,光周期为光亮(L)∶黑暗(D)=12 h∶12 h。
1. 2 试验设计
挑选预培养后生长健康、形态完好的缢江蓠藻体1.0 g(鲜质量)培养于800 mL三角烧瓶中,置于智能型光照培养箱(GXZ-300D)中培养。基于预培养效果分别设计3个生态单因子的不同水平培养试验,试验过程中每隔2 d更换一次灭菌消毒海水并测定鲜藻质量,每处理组均设3个重复,培养14 d后开始进行相关测定分析。
1. 2. 1 温度 试验共设6个温度梯度(16、20、24、28、32和36 ℃)处理组,其他生态因子一致,即盐度30‰,光照强度6000 lx,光周期为光亮(L)∶黑暗(D)=
12 h∶12 h。
1. 2. 2 光照强度 试验设4个光照强度梯度(3000、6000、9000和12000 lx)处理组,其他生态因子一致,即盐度30‰,温度30 ℃,光周期为光亮(L)∶黑暗(D)=
12 h∶12 h。
1. 2. 3 盐度 试验设7个盐度梯度(10‰、15‰、20‰、25‰、30‰、35‰和40‰)处理组,其他生态因子一致,即温度30 ℃,光照强度6000 lx,光周期为光亮(L)∶黑暗(D)=12 h∶12 h。
1. 3 检测指标及方法
(1)藻体相对生长率(RGR):通过测定藻体在培养14 d内其鲜质量(FM)的变化,再根据RGR(%/d)=[1n(Mt/M0)/t]×100进行计算。其中,M0为初始鲜质量,Mt为培养14 d后的鲜质量,称量前用吸水纸吸干藻体水分。(2)叶绿素a(Chla)采用乙醇法进行测定,再参照Porra(2002)的方法计算Chla含量。(3)类胡萝卜素(Car)采用乙醇法进行测定,再根据Parsons和Srickland(1963)的方法计算Car含量。(4)藻红蛋白(R-PE)和藻蓝蛋白(R-PC)含量参考Beer和Eshel(1985)的方法进行测定。(5)可溶性蛋白含量采用考马斯亮蓝G250染色法(张志良和瞿伟菁,2002)进行测定。(6)超氧化物歧化酶(SOD)活性、丙二醛(MDA)含量分别采用黄嘌呤氧化酶法和硫代巴比妥酸法(邹崎,2000)进行测定。
1. 4 统计分析
试验数据采用Excel 2007和SPSS 19.0进行处理及统计分析,并以One-way ANOVA(LSD)和t 检验其差异显著性。
2 结果与分析
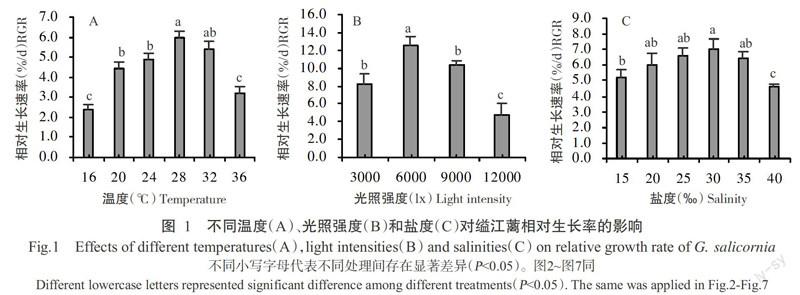
2. 1 温度、光照强度和盐度对缢江蓠生长的影响
2. 1. 1 温度变化对缢江蓠生长的影响 从图1-A可看出,在16~36 ℃范围内,缢江蓠的相对生长率先升高后下降,以28 ℃处理组的相对生长率最高,为生长率曲线的拐点。在16~28 ℃范围内,缢江蓠的生长一直保持增长趋势,其间的相对生长率以20 ℃处理组显著高于16 ℃处理组(P<0.05,下同),从20 ℃上升到28 ℃的相对生长率保持平稳增长;温度超过28 ℃后,缢江蓠的生长随温度升高呈下降趋势,其中36 ℃处理组的相对生长率显著低于32 ℃处理组。
2. 1. 2 光照强度变化对缢江蓠生长的影响 从图1-B可看出,光照强度变化对缢江蓠的生长率有明显影响。在3000~12000 lx范围内,缢江蓠的相对生长率呈先升高后下降的变化趋势,以6000 lx处理组的生长最快,为光照强度变化对缢江蓠生长率曲线的拐点。光照强度为3000~6000 lx时,缢江蓠的生长呈增长趋势;光照强度超过6000 lx后,缢江蓠的生长呈不断下降趋势,以12000 lx处理组的相对生长率最低,与其他处理组的差异均达显著水平。
2. 1. 3 盐度变化对缢江蓠生长的影响 从图1-C可看出,盐度变化对缢江蓠的生长也有明显影响。在15‰~40‰范围内,缢江蓠的相对生长率呈先升高后下降趋势,以30‰处理组的生长最快,显著高于15‰和40‰处理组,但与20‰、25‰和35‰处理组的差异不显著(P>0.05,下同)。40‰处理组的相对生长率最低,与其他处理组差异显著。
2. 2 温度、光照强度和盐度对缢江蓠色素含量的影响
2. 2. 1 温度变化对缢江蓠色素含量的影响 温度变化对缢江蓠色素含量有明显影响(图2-A),在16~36 ℃范围内,缢江蓠的Chla和Car含量均表现为先升高后下降,且以20 ℃为温度对缢江蓠色素含量曲线的拐点。在16~20 ℃范围内,缢江蓠的Chla和Car含量呈升高趋势;超过20 ℃后,Chla和Car含量呈下降趋势;至36 ℃时Chla和Car含量最低,显著低于20 ℃处理组。
2. 2. 2 光照强度变化对缢江蓠色素含量的影响 光照强度变化对缢江蓠的Chla和Car含量也有明显影响(图2-B),在3000~12000 lx范围内,两种色素含量均呈先升高后下降的变化趋势,且以6000 lx为光照强度对缢江蓠色素含量变化曲线的拐点。光照强度升至12000 lx时,缢江蓠的Chla和Car含量均最低,且两种色素含量与其他处理组的差异均达显著水平。
2. 2. 3 盐度变化对缢江蓠色素含量的影响 盐度变化对缢江蓠Chla和Car含量的影响一致,均呈先升高后下降的变化趋势(图2-C),但含量变化曲线的拐点不同,分别为35‰和30‰。缢江蓠的Chla和Car含量均以40‰处理组最低,明显低于其他盐度处理组。
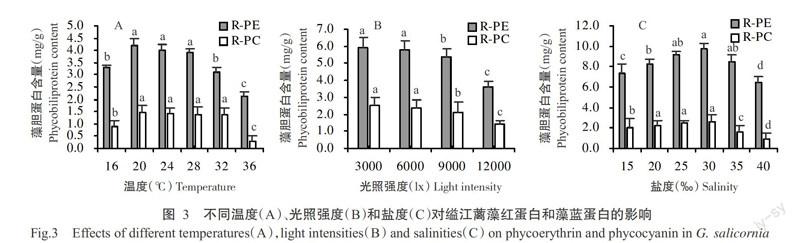
2. 3 温度、光照强度和盐度对缢江蓠藻胆蛋白含量的影响
2. 3. 1 温度变化对缢江蓠藻胆蛋白含量的影响 温度变化对缢江蓠藻胆蛋白含量有明显影响(图3-A),在16~36 ℃范围内,缢江蓠的R-PE和R-PC均呈先升高后下降的变化趋势,且以20 ℃为缢江蓠藻胆蛋白含量曲线的拐点。温度超过20 ℃后,两种藻胆蛋白含量均呈明显的下降趋势;至36 ℃时,两种藻胆蛋白的含量最低,显著低于其他处理组。
2. 3. 2 光照强度变化对缢江蓠藻胆蛋白含量的影响
光照强度变化对缢江蓠藻胆蛋白含量有明显影响(图3-B),随着光照强度的增强,缢江蓠的R-PE和R-PC含量逐渐降低,至12000 lx时两种藻胆蛋白的含量最低,与其他光照强度处理组间的差异达显著水平。
2. 3. 3 盐度变化对缢江蓠藻胆蛋白含量的影响 盐度变化对缢江蓠藻胆蛋白含量变化也有明显影响(图3-C),在15‰~40‰范围内,缢江蓠R-PE和R-PC均呈先升高后下降的变化趋势,且以30‰为缢江蓠藻胆蛋白含量曲线的拐点。当盐度为40‰时,缢江蓠的两种藻胆蛋白含量均显著低于其他盐度处理组。
2. 4 温度、光照强度和盐度对缢江蓠可溶性蛋白含量的影响
2. 4. 1 温度变化对缢江蓠可溶性蛋白含量的影响 温度变化对缢江蓠可溶性蛋白含量有明顯影响(图4-A),在16~36 ℃范围内,缢江蓠可溶性蛋白含量先升高后下降,以20 ℃为可溶性蛋白含量变化曲线的拐点。20 ℃处理组的缢江蓠可溶性蛋白含量显著高于16、32和36 ℃处理组,但与其他处理组间的差异不显著。
2. 4. 2 光照强度变化对缢江蓠可溶性蛋白含量的影响 光照强度变化对缢江蓠可溶性蛋白含量有明显影响(图4-B),随着光照强度的增强,缢江蓠可溶性蛋白含量逐渐降低,其中12000 lx处理组的可溶性蛋白含量最低,显著低于其他光照强度处理组。
2. 4. 3 盐度变化对缢江蓠可溶性蛋白含量的影响 盐度变化对缢江蓠可溶性蛋白含量也有明显影响(图4-C),在15‰~40‰范围内,缢江蓠可溶性蛋白含量呈先升高后下降的变化趋势,以盐度为30‰时的可溶性蛋白含量最高,盐度为40‰时的可溶性蛋白含量最低,二者差异显著。
2. 5 温度、光照强度和盐度对缢江蓠SOD活性和MDA含量的影响
2. 5. 1 温度变化对缢江蓠SOD活性和MDA含量的影响 如图5-A所示,在16~36 ℃范围内,缢江蓠SOD活性随温度的升高呈先下降后上升再下降的变化趋势,其中以32 ℃处理组的SOD活性最高、36 ℃处理组的SOD活性最低,二者差异显著。在16~36 ℃范围内,缢江蓠MDA含量随温度的升高呈先下降后上升的变化趋势(图5-B),以28 ℃处理组的MDA含量最低、36 ℃处理组的MDA含量最高,二者差异显著。
2. 5. 2 光照强度变化对缢江蓠SOD活性和MDA含量的影响 如图6-A所示,在3000~12000 lx范围内,缢江蓠SOD活性随光照强度的增强呈先上升后下降趋势,以9000 lx处理组的SOD活性最高,与其他光照强度处理组差异显著;12000 lx处理组的SOD活性最低,与其他各光照强度处理组的差异也达显著水平。在3000~12000 lx范围内,缢江蓠MDA含量随光照强度的增强而逐渐上升(图6-B),其中以12000 lx处理组的MDA含量最高,与其他光照强度处理组的差异显著。
2. 5. 3 盐度变化对缢江蓠SOD和MDA含量的影响 如图7-A所示,在15‰~40‰的范围内,缢江蓠SOD活性随盐度的升高呈先上升后下降的变化趋势,以30‰处理组的SOD活性最高、40‰处理组的SOD活性最低,二者差异显著。在15‰~40‰的范围内,缢江蓠MDA含量变化趋势与SOD活性的变化趋势恰好相反,随盐度的升高呈先降低后上升趋势(图7-B),以30‰处理组的MDA含量最低、40‰处理组的MDA含量最高,二者差异显著。
3 讨论
3. 1 温度对缢江蓠生长率及生化组分含量的影响
温度对大型海藻的生长、生理活动及地理分布起重要作用(李恒等,2013)。本研究结果表明,在16~36 ℃范围内,缢江蓠的Chla和Car含量均表现为先升高后下降,其原因是低温不利于藻体光合色素合成,但温度过高会影响细胞内酶活性和叶绿素合成,且加速叶绿素分解(钱鲁闽等,2006)。藻体内可溶性蛋白含量变化也反映出温度过高会影响藻体细胞内酶活性,使可溶性蛋白合成减少且消耗速度加快,进而导致其含量减少(潘瑞炽,2001)。藻体SOD活性的变化规律表明过低或过高的温度对藻体均产生胁迫影响,SOD活性较低,既能防止H2O2积累,又避免了超氧阴离子和H2O2反应生成毒性更强的羟基自由基,从而减轻活性氧的伤害。温洋等(2005)认为,SOD可作为鉴定植物耐热性的重要生理指标之一。本研究中,SOD活性以32 ℃处理组的最高,说明缢江蓠具有较好的耐热性。MDA含量可间接反应藻体受胁迫的程度(汪耀富等,1996)。本研究结果表明,缢江蓠MDA含量随温度的升高呈先降低后上升的变化趋势,说明温度过低或过高均会抑制缢江蓠生长。不同江蓠的适合生长温度也不一样,龙须菜的最适生长温度为25 ℃(刘树霞等,2009),细基江蓠繁枝变型的最适温度在25 ℃左右(刘静雯和董双林,2001),真江蓠的最适生长温度为25 ℃(霍元子等,2010),脆江蓠的最适生长温度为25 ℃(金玉林等,2012),芋根江蓠的最适生长温度为33 ℃(黄中坚等,2014),智利江蓠的最适温度为21 ℃(陈伟洲等,2015)。综合本研究结果可知,缢江蓠的适宜生长温度为24~32 ℃,最适温度为28 ℃,即缢江蓠具有能适应较高温度下生长的特性。
3. 2 光照强度对缢江蓠生长及生化组分的影响
光照强度是影响藻类生长的重要生态因子。在一定的光照强度范围内(3000~6000 lx),缢江蓠的相对生长率和光合色素含量均随光照强度的增强而上升,达最大值后二者又逐渐降低,至12000 lx时,其含量为最低值,表明适当的光照强度能促进缢江蓠生长,过高的光照强度则会破坏藻体内的光合色素和酶活性,对藻体产生光抑制或对藻体产生伤害,阻碍其生长。缢江蓠的藻胆蛋白、可溶性蛋白表现出低光照强度处理组的含量较高,说明低光照强度更有利于藻体内生化组分的积累,与缢江蓠分布于中低潮带不喜强光的自然习性相符,是其对环境变化的一种生理适应(林贞贤等,2007)。SOD活性水平可在一定程度上反映藻体耐氧化性,即耐逆本领。本研究结果表明,在过低或过高的光照强度下,缢江蓠SOD活性均较低,可能是在适宜缢江蓠生长低光照强度处理组产生的胁迫较小,高光照强度下藻体发白枯烂而影响SOD活性。MDA含量随光照强度的增强而逐渐增加,表明缢江蓠具有喜低光、不耐强光的特性。综合上述,缢江蓠对光照强度的适应范围为3000~6000 lx,最适光照强度为6000 lx。
3. 3 盐度对缢江蓠生长及生化组分的影响
盐度是影响大型海藻生长的一个重要因素(Yokoya and Oliveira,1992;Phooprong et al.,2007),其高低决定了海水渗透压。盐度过低或过高均会影响藻类的生理活性,从而影响其生长状况(郑兰红,2004)。本研究结果表明,缢江蓠对盐度的适应范围较广,适宜生长的盐度为20‰~35‰,最适生长盐度为30‰,具有耐受较高盐度的特性。盐度15‰和40‰处理组的缢江蓠相对生长率、色素含量、藻胆蛋白含量和可溶性蛋白含量均较低,是由于高盐或低盐胁迫会影响其光合速率和蛋白质合成效率,从而影响其生长过程中的物质积累。为应对低盐或高盐胁迫,缢江蓠SOD活性降低,其原因是逆境胁迫会促进活性氧产生,可直接破坏生物大分子,使酶活性丧失,造成氧化胁迫,即胁迫程度越高,酶活性越低。此外,盐度15‰和40‰处理组的缢江蓠MDA含量明显升高,是因为藻体受低盐或高盐胁迫时,其活性氧清除系统的功能逐渐降低,活性氧积累越来越多,最终促使细胞膜发生膜质过氧化而引发自由基链式反应,形成MDA,使细胞膜流动性下降,膜功能受到伤害。
4 结论
缢江蓠具有能适应高温、低光照强度及中、高盐度环境条件的特性,可在我国南方海域夏季自然条件下栽培生产。
参考文献:
陈昌生,章景荣. 1999. 福建省细基江蓠繁枝变型人工栽培的探讨[J]. 集美大学学报(自然科学版),4(2):65-71. [Chen C S,Zhang J R. 1999. A discussion on the techniques of artificial cultivation of Gracilaria tenuistipitata var. Liui in Fujian[J]. Journal of Jimei University (Natural Science),4(2):65-71.]
陈伟洲,钟志海,刘涛,黄中坚,赖学文. 2015. 光照强度和温度对智利江蓠生长及生化组分的影响[J]. 海洋湖沼通报,(1):28-34. [Chen W Z,Zhong Z H,Liu T,Huang Z J,Lai X W. 2015. Effects of different light intensities and temperature on growth and chemical constituents of Graci-
laria chilensis[J]. Transactions of Oceanology and Limnology,(1):28-34.]
黃中坚,宋志民,杨晓,赖学文,刘涛,陈伟洲. 2014. 生态因子对芋根江蓠的生长及生化组分的影响[J]. 南方水产科学,10(1):27-34. [Huang Z J,Song Z M,Yang X,Lai X W,Liu T,Chen W Z. 2014. Effect of ecological factors on growth and biochemical constituents of Gracilaria blodgettii[J]. South China Fisheries Science,10(1):27-34.]
霍元子,徐姗嫡,张建恒,韩芳,董丽,何培民. 2010. 真江蓠杭州湾海域栽培试验及生态因子对藻体生长的影响[J]. 海洋科学,34(8):23-28. [Huo Y Z,Xu S D,Zhang J H,Han F,Dong L,He P M. 2010. Trial and field cultivation of Gracilaria verrucosa in Hangzhou bay and effects of ecological factors on its growth[J]. Marine Sciences,34(8):23-28.]
金玉林,吴文婷,陈伟洲. 2012. 不同温度和盐度条件对脆江蓠生长及其生化组分的影响[J]. 南方水产科学,8(2):51-57. [Jin Y L,Wu W T,Chen W Z. 2012. Effects of different temperature and salinity on growth and biochemical constituents of Gracilaria chouae[J]. South China Fisheries Science,8(2):51-57.]
李恒,李美真,曹婧,徐智广. 2013. 温度对几种大型海藻硝氮吸收及其生长的影响[J]. 渔业科学进展,34(1):159-165. [Li H,Li M Z,Cao J,Xu Z G. 2013. Effects of temperature on nitrogen uptake and growth in several species of macroalgae[J]. Progress in Fishery Sciences,34(1):159- 165.]
林贞贤,宫相忠,李大鹏. 2007. 光照和营养盐胁迫对龙须菜生长及生化组成的影响[J]. 海洋科学,31(11):22-26. [Lin Z X,Gong X Z,Li D P. 2007. Effects of light and the stress of nutrients deficiency on the growth and levels of chemical constituents of Gracilaria lemaneiformis[J]. Marine Sciences,31(11):22-26.]
刘静雯,董双林. 2001. 光照和温度对细基江蓠繁枝变型的生长及生化组成影响[J]. 青岛海洋大学学报,31(3):332- 338. [Liu J W,Dong S L. 2001. Interactions between light and temperature on growth and levels of chemical constituents of Gracilaria tenuistipitata var. Liui[J]. Journal of Ocean University of Qingdao,31(3):332-338.]
刘树霞,徐军田,蒋栋成. 2009. 温度对经济红藻龙须菜生长及光合作用的影响[J]. 安徽农业科学,37(33):16322- 16324. [Liu S X,Xu J T,Jiang D C. 2009. The effects of temperature on the growth and photosynthesis of economic red macroalga Gracilaria lemaneiformis[J]. Journal of Anhui Agriculture Science,37(33):16322-16324.]
卢晓,李美真,王志刚,胡凡光,吴海一,孙福新. 2014. 光照和温度对脆江蓠的生长和生化组成的影响[J]. 中国水产科学,21(6):1236-1243. [Lu X,Li M Z,Wang Z G,Hu F G,Wu H Y,Sun F X. 2014. Effects of different temperature and illumination intensity on growth and biochemical constituents of Gracilaria chouae[J]. Journal of Fishery Sciences of China,21(6):1236-1243.]
潘瑞熾. 2001. 植物生理学[M]. 第4版. 北京:高等教育出版社. [Pan R C. 2001. Phytophysiology[M]. The 4th Edition. Beijing:Higher Education Press.]
钱鲁闽,徐永健,焦念志. 2006. 环境因子对龙须菜和菊花心江蓠N、P吸收速率的影响[J]. 中国水产科学,13(2):251- 262. [Qian L M,Xu Y J,Jiao N Z. 2006. Effects of en-
vironmental factors on uptake of nitrogen and phosphorus by Gracilaria lemaneiformis and G.1ichevoides[J]. Journal of Fishery Sciences of China,13(2):251-262.]
覃媚,于清武,竺利波,李菲,颜栋美,余炼,高程海. 2016. 三种江蓠共附生细菌多样性及抑菌活性分析[J]. 南方农业学报,47(11):1966-1973. [Qin M,Yu Q W,Zhu L B,Li F,Yan D M,Yu L,Gao C H. 2016. Diversity of epiphytic bacteria of three species of Gracilaria and their bacteriostatic activities[J]. Journal of Southern Agriculture,47(11):1966-1973.]
孙雪,蔡西栗,徐年军. 2013. 海洋红藻龙须菜对2种逆境温度胁迫的应激生理响应[J]. 水生生物学报,37(3):535-540. [Sun X,Cai X L,Xu N J. 2013. Physiological response of marine red algae Gracilaria lemaneiformis of two kinds of adverse temperature stress[J]. Acta Hydrobio-
logica Sinica,37(3):535-540.]
汤坤贤,焦念志,游秀萍,陈敏儿,沈东煜,林亚森,林泗彬. 2005. 菊花江蓠在网箱养殖区的生物修复作用[J]. 中国水产科学,12(2):156-161. [Tang K X,Jiao N Z,You X P,Chen M E,Shen D Y,Lin Y S,Lin S B. 2005. Bioremediation of Gracilaria lichenoides in fish cages-farming areas[J]. Journal of Fishery Sciences of China,12(2):156-161.]
汪耀富,韩锦峰,林学悟. 1996. 烤烟生长前期对干旱胁迫的生理生化影响研究[J]. 作物学报,22(1):117-122. [Wang Y F,Han J F,Lin X W. 1996. Study on physiological and biochemical responses of flue-cured tobacco to drought stress during early growth of the plants[J]. Acta Agronomica Sinica,22(1):117-122.]
温洋,孙吉雄,王代军. 2005. 干旱胁迫对冷地型草坪草生理特性影响的研究[J]. 草原与草坪,(1):43-45. [Wen Y,Sun J X,Wang D J. 2005. Effects of drought stress to the physiological characteristics in cool season turfgrasses[J]. Grassland and Turf,(1):43-45.]
徐永健,王永胜,韦玮. 2006. 多因子交互作用对菊花江蓠氮、磷吸收速率的影响[J]. 水产科学,25(5):222-226. [Xu Y J,Wang Y S,Wei W. 2006. Effects of multi-factor interaction on uptake rates of nitrogen and phosphorus by Gracilaria lichenoides[J]. Fisheries Science,25(5):222- 226.]
许忠能,林小涛,计新丽,王朝晖,黄长江. 2001. 环境因子对细基江蓠繁枝变种氮、磷吸收速率的影响[J]. 应用生态学报,12(3):417-421. [Xu Z N,Lin X T,Ji X L,Wang C H,Huang C J. 2001. Effect of environmental factors on N and P uptake by Gracilaria tenuistipitata var. Liui Zhang et Xia[J]. Chinese Journal of Applied Ecology,12(3):417- 421.]
张学成,费修绠,王广策,林祥志,陈伟洲,隋正红,徐涤,臧晓南. 2009. 江蓠属海藻龙须菜的基础研究与大规模栽培[J]. 中国海洋大学学报,39(5):947-954. [Zhang X C,Fei X G,Wang G C,Lin X Z,Chen W Z,Sui Z H,Xu D,Zang X N. 2009. Genetic studies and large scale cultivation of Gracilaria lemaneiformis[J]. Periodical of Ocean University of China,39(5):947-954.]
张志良,瞿伟菁. 2002. 植物生理学实验指导[M]. 第3版. 北京:高等教育出版社. [Zhang Z L,Qu W J. 2002. Experiment Instruction for Phytophysiology[M]. The 3rd Edieion Beijing: Higher Education Press.]
郑兰红. 2004. 冈村枝管藻(Cladosiphon ckamuranus)培养及实验生态学基础研究[D]. 青岛:中国海洋大学. [Zheng L H. 2004. Primary study of culture and environmental factors on the growth of Cladosiphyon okamuranus[D]. Qingdao:Ocean University of China.]
邹崎. 2000. 植物生理学实验指导[M]. 北京:中国农业出版社. [Zou Q. 2000. Phytophysiology Experiment Instruction [M]. Beijing:China Agriculture Press.]
Beer S,Eshel L A. 1985. Determining phyeoerythrin and phycoeyanin concentrations in aqueous crude extracts of red algae[J]. Marine and Freshwater Research,36(6):785-792.
Parsons T R,Srickland J D H. 1963. Disscussion of spectropho-
tometric determination of marine plant pigments,with revised equalion for ascertaining chlorophylls and carotenoids[J]. Journal of Marine Research,2l(3):155-163.
Phooprong S,Ogawa H,Hayashizaki K. 2007. Photosynthetic and respiratory responses of Gracilaria salicornia(C. Ag.) Dawson(Gracilariales, Rhodophyta) from Thailand and Japan[J]. Journal of Applied Phycology,19(6):795-801.
Phooprong S,Ogawa H,Hayashizaki K. 2008. Photosynthetic and respiratory responses of Gracilaria vermiculophylla (Ohmi) Papenfuss collected from Kumamoto, Shizuoka and Iwate,Japan[J]. Journal of Applied Phycology,20(5):743-750.
Porra R J. 2002. The chequered history of the development and use of simultaneous equations for the accurate determination of chlorophylls a and b[J]. Photosynthesis Research,73(1):149-156.
Wang Z Y,Wang G C,Niu J F,Wang W J,Peng G. 2010. Optimization of conditions for tetraspore release and assessment of photosynthetic activities for different generation branches of Gracilaria lemaneiformis Bory[J]. Chinese Journal of Oceanology and Limnology,28(4):738-748.
Yang Y F,Fei X G,Song J M,Hu H Y,Wang G C,Chung I K. 2006. Growth of Gracilaria lemaneiformis under different cultivation conditions and its effects on nutrient removal in Chinese coastal waters[J]. Aquaculture,254(1-4):248-255.
Yokoya N,Oliveira E. 1992. Effects of salinity on the growth rate,morphology and water content of some brazilian red algae of economic importance[J]. Ciências Marina,18:49-64.
(責任编辑 兰宗宝)
