26份割手密种质材料的抗旱性差异评价
2017-05-30王天菊王先宏杨清辉
王天菊 王先宏 杨清辉

摘 要 以26份割手密为试验材料,在正常供水和干旱胁迫处理下,测定伸长初期的13项形态指标、生理指标及光合参数,利用相关分析、聚类分析和模糊隶属函数分析法对其抗旱性差异进行综合评价。结果表明:干旱胁迫后,不同材料之间的抗旱指标存在明显差异,其抗旱性不同。相关分析表明,不同基因型割手密的染色体数与抗旱性呈极显著负相关,叶面积与抗旱性呈显著负相关。模糊隶属函数和聚类分析可将26份不同基因型割手密分为3类,其中高抗旱类型1份(2013-13),中抗旱类型14份,不抗旱类型11份,为割手密抗旱材料筛选及甘蔗抗旱基因发掘奠定基础。
关键词 甘蔗;割手密;抗旱鉴定;模糊隶属函数法
中图分类号 S566.1 文献标识码 A
Comprehensive Evaluation on Drought Resistance Difference
of Twenty-six Saccharum spontaneum L. Accessions
WANG Tianju, WANG Xianhong*, YANG Qinghui*
College of Agriculture and Biological Technology, Yunnan Agriculture University / Sugarcane
Research Institution, Yunnan Agricultural University, Kunming, Yunnan 650201, China
Abstract Twenty-six S. spontaneum accessions were used as the materials under the normal water and drought stress treatment, 13 morphological indices, physiological indices and photosynthetic parameters were measured at early elongation stage. Comprehensive evaluation of drought resistance by the methods of correlation, clustering, fuzzy membership function analysis were undertaken. The results showed that the drought resistance index of different materials were obviously different. The drought resistance of S. spontaneum accessions were also different under the drought stress treatment. Correlation analysis showed the relative values of chromosome numbers and drought resistance were highly significantly negatively correlated. And the relative values of leaf area and drought resistance were highly significantly negatively correlated. Membership function and cluster analysis showed that the twenty-six accessions could be divided into three categories, including one accession as the high drought-resistant type(2013-13), 14 accessions as the drought-resistant type and 11 accessions as the drought-sensitive type. The study would lay the foundation for exploring drought-resistant genes to improve new available sugarcane varieties.
Key words Sugarcane; Saccharum spontaneum L.; drought resistance identification; Fuzz membership function
doi 10.3969/j.issn.1000-2561.2017.09.011
中國目前大约90%蔗区在缺水无灌溉条件的丘陵旱坡地[1-2],干旱已成为制约中国甘蔗生产的主要因素之一。生产实践证明,培育甘蔗高产抗旱新品种是提高水分利用效率最重要、最有效的措施之一[3-4]。割手密(Saccharum spontaneum L.)具有旱生植物所具有的特征,又是甘蔗育种中最有价值的资源植物[5-6],利用割手密的抗旱特性培育抗旱性强、水分利用率高的新品种,对中国甘蔗产业发展具有非常重要的意义。
近年来国内外学者从形态、生理生化的角度,对割手密的抗旱性鉴定指标、评价方法和抗旱种质(品种)筛选做了较多研究。在干旱胁迫下,割手密叶片的束缚水含量、丙二醛含量、脯氨酸含量、可溶性糖含量、质膜透性、超氧化物歧化酶活性、过氧化物酶活性均升高,而株高长速、功能叶重、根系活力、根/冠比、叶片含水量、失水速率、叶绿素含量、过氧化氢酶活性和净光合速率均下降[7-10],这些变化使割手密能够长期适应热带和亚热带的生长环境。因此上述指标均可作为鉴定割手密抗旱能力较可靠的依据。割手密抗旱性评价方法有模糊隶属函数法[9-11]、田间自然鉴定法[12]、判别分析法[8]和聚类分析法[8,13],其中模糊隶属函数法能够较准确评价割手密的抗旱性,应用较多。采用这些方法,国内学者已经鉴定出川79-1-15等一批抗旱性较强的野生资源[10,12],筛选出云割F3 95/128等一批抗旱性优越的割手密后代及其血缘的种质创新材料,拓宽了甘蔗抗旱育种的遗传基础[7-9,11,13]。
割手密是一种基因型多样的植物,其染色体数(2n)在40~128之间,染色体基数为8,其中以2n=64、80、96、112和128等5种染色体类型为主[14-117],然而针对国内外不同基因型割手密抗旱鉴定与筛选的研究尚未见报道。本研究在割手密资源调查、植物学性状分析以及核型分析等研究基础上[16-20],对26份不同基因型割手密材料通过测定干旱胁迫条件下生长发育、形态性状和生理生化的变化,以抗旱隶属函数为依据,采用综合抗旱性评价的方法进行抗旱性筛选和评价,以期为甘蔗抗旱育种提供新的种质资源。
1 材料与方法
1.1 材料
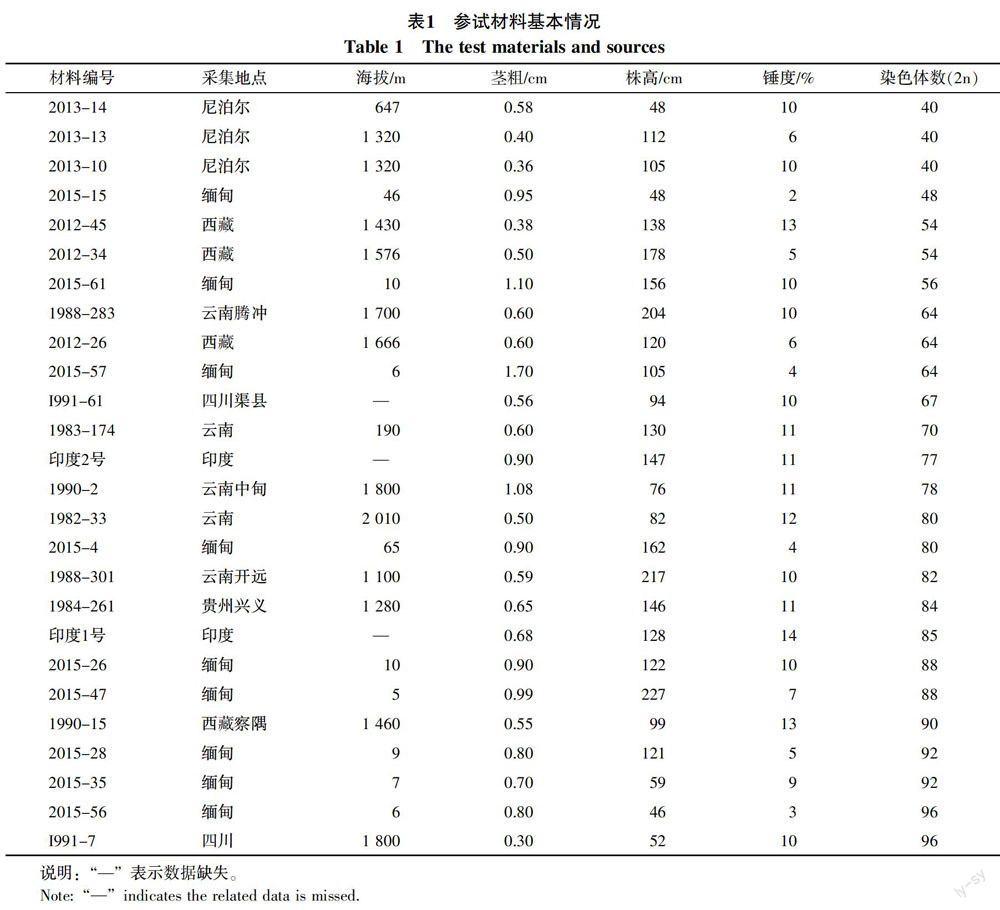
从云南农业大学农学院甘蔗研究所资源圃现有的种质资源中,选择不同基因型或基因型相同但采集地点不同的26份割手密材料(表1)。
1.2 方法
1.2.1 试验设计 盆栽试验在避雨、自然通风的塑料大棚内进行,采用土培的方法,塑料盆底留孔,带底盘,每盆装土8 kg。将供试材料均匀分兜后种植于塑料盆中,每份材料种植6盆,共156盆。材料放置于塑料大棚中,胁迫处理前施肥浇水量一致,其它管理同一般的盆栽管理。
供试材料处于伸长初期时(2016年5月24日),首先把每份材料随机分成2组,每组3盆,充分淋水直至土壤水分饱和,盆底有水流出,然后进行水分处理,一组正常供水,作为对照(每3 d灌溉1次),另一组停止供水,进行干旱胁迫处理。干旱胁迫期间每日9时观察材料生长情况,并对盆内土壤含水量用烘干法进行跟踪测定,根据测定结果决定干旱胁迫处理的时间。
当干旱胁迫处理到土壤含水量达到重度缺水时(2016年5月31日),材料表现出严重干旱状态。严重缺水按Hsiao的标准进行划分,相对含水量降低大于20%属严重缺水,此时植株从上到下叶片的叶尖枯萎,甚至整叶枯死[21]。
1.2.2 测定项目与方法 干旱胁迫处理结束后,对所有供试材料同时取样,每份材料取足量+1叶(最高可见肥厚带所在的叶片)混合后,置于封口袋中迅速带回实验室,放-20 ℃保存待用。参考前人研究方法,测定能反映割手密抗旱能力的13项形态指标、生理指标和光合参数[7-11]。株高采用带刻度的直尺测量。绿叶数、黄叶数采用目测法测定,绿叶率/%=绿叶数/(绿叶数+黄叶数+枯叶数)×100%。叶片相对含水量(RWC)采用饱和称量法[22]测定,RWC/%=(叶片鲜重-干重)/(水饱和重-干重)×100%。光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Sc)等光合参数采用LI-6400XT光合速率测定仪在资源圃实地测定[23]。水势(WP)采用WP4T型水势测定仪测量。丙二醛含量(MDA)测定采用Health等的方法[24];叶绿素含量(ChL)测定采用80%丙酮浸提法[25];超氧化物歧化酶活性(SOD)测定参照Giannoplitis等的方法[26];过氧化物酶活性(POD)测定采用李杨瑞[27]的方法;脯氨酸含量(Pro)采用茚三酮法测定[28];质膜透性(PMP)根据谭常的方法测定[29]。伸长期叶面积参照蔡泽霖[30]的方法测定。
1.3 统计分析
数据整理后用SPSS17.0软件进行统计和聚类分析。绿叶率/%=绿叶数/(绿叶数+黄叶数+枯叶数)×100%;株高长速/cm=胁迫后株高-胁迫前株高,抗旱性综合评价采用模糊隶属函数评价法[31],计算公式如下:
式中,∧Xij为第i个材料第j个性状的隶属函数值,Xij为第i个材料第j个性状值,Xjmin为各材料j性状的最小值,Xjmax为各材料j性状的最大值。Xi为i基因型的模糊隶属函数均值,n为指标数,模糊隶属函数值的区间为[0,1]。若指标与抗旱性呈正相关,用公式(1)计算模糊隶属函数值,若指标与抗旱性呈负相关,选用公式(2)计算模糊隶属函数值,最后用公式(3)计算模糊隶属函数均值,Xi值越大,表明该材料抗旱性越强。
2 结果与分析
2.1 干旱胁迫下割手密材料各指标的变化
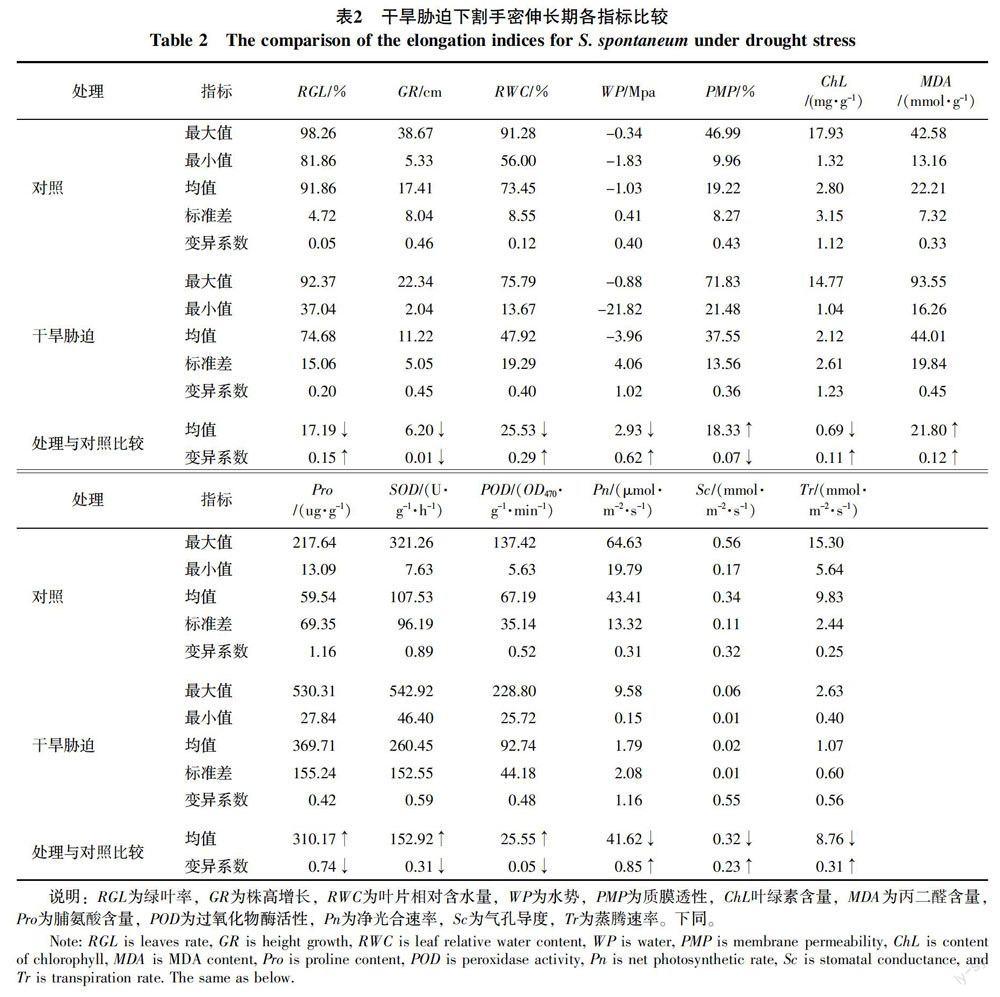
26份不同基因型割手密13项指标的测定结果表明(表2),在干旱胁迫下,割手密的绿叶率为37.04%~92.37%,平均为74.68%,较对照下降17.19%;株高增长为2.04~22.34 cm,平均为11.22 cm,较对照下降了6.20 cm,其它指标也发生了不同程度的变化,其中叶片相对含水量均值较对照减少25.53%、水势均值较对照减少2.93 Mpa、叶绿素均值较对照减少0.69 mg/g,净光合速率、气孔导度、蒸腾速率分别减少41.62 μmol/(m2·s)、0.32 mmol/(m2·s)、8.76 mmol/(m2·s)。可见,不同基因型割手密受干旱胁迫影响的形態指标、生理指标和光合指标不同,且影响程度存在差异,表明抗旱性不同。
此外,干旱胁迫处理与对照相比,除了株高增长、质膜透性、脯氨酸、超氧化物酶、过氧化物酶的变异系数有稍微的下降外,其它指标的变异系数有不同程度的增长,其中增长最大的是净光合速率,其变异系数比对照增加了0.85,其次是水势,其变异系数比对照增加了0.62。可见,净光合速率和水势对干旱胁迫最敏感。综上所述,不同基因型割手密抗旱性与多种指标有关,运用任何单一指标进行评价均存在一定的片面性,需对其进行综合分析和评价。
2.2 割手密材料的抗旱性综合评定
2.2.1 模糊隶属函数 利用模糊隶属函数法计算隶属函数加权平均值,对26份不同基因型割手密的抗旱性进行综合评价(表3)。结果显示,干旱胁迫下26份不同基因型割手密材料的抗旱性由强到弱排序依次为:2013-13>2013-10>2013-14>1990-2>2015-57>2015-15>I991-61>2015-47>印度2号>1983-174>2015-61>2012-45>2012-26>1988-301>2015-56>I991-7>1988-283>2012-34>2015-4>1990-15>2015-26>印度1号>2015-35>2015-28>1982-33>1984-261。可以看出材料2013-13叶片的丙二醛含量、叶绿素含量,叶片相对含水量和质膜透性等指标的模糊隶属函数值都达到了最大值1,表明在干旱胁迫下2013-13通过合成大量的保护性物质或改变生理特性来提高自身的抗旱性,因此其表现较强的抗旱能力。相反,材料1984-261叶片的丙二醛含量、脯氨酸含量,水势和质膜透性等指标的模糊隶属函数值相对比较小,抗旱性表现最弱。
2.2.2 聚類分析 采用系统聚类法对材料的模糊隶属函数值进行聚类分析(图1),结果表明26份不同基因型割手密材料划分为3类,2013-13自成一类,可列为高度抗旱类型,占供试材料的3.8%; 2015-56、2012-26、1988-301、2012-45、2013-10、2015-61、1983-174、2015-47、印度2号、2015-57、1990-2、2013-14、2015-15和I991-61共14个材料聚为一类,列为中度抗旱类型,占供试材料的53.85%;1984-261、1982-33、2015-26、1990-15、2015-4、2012-34、1988-283、I991-7、2015-35、2015-28和印度1号共11个材料聚为一类,列为不抗旱类型,占供试材料的42.31%。
2.3 26份割手密材料抗旱性与叶面积、基因型的相关性
正常供水条件下,26份割手密材料伸长期的叶片形态特征差异明显,叶面积大小不同(表4)。叶面积最小的是2013-13,最大的是2015-57,将模糊隶属函数评价出的抗旱性不同的割手密与植株的染色体数和伸长期的叶面积进行相关性分析表明,染色体数与叶面积呈极显著正相关,相关系数为0.558;染色体数与抗旱隶属函数呈极显著负相关,相关系数为0.653;叶片面积与抗旱隶属函数呈显著负相关,相关系数是0.398(表5)。可见,叶面积大小和染色体数多少对不同基因型割手密的抗旱强弱有显著影响,可作为割手密抗旱种质材料筛选的参考依据。
3 讨论
抗旱性为多基因控制的数量性状,涉及的基因和代谢众多[32],另外,群体、个体及环境的作用与割手密的抗旱性密切相关,例如植株的高矮,茎秆的粗细,叶片的气孔大小和角质化程度,根系的粗细和长短,生长环境等因素,因此准确评价割手密的抗旱性是相当困难的[8]。
作物在干旱条件下主要通过质膜透性、渗透调节、抗氧化保护酶系统清除自由基的能力等方式产生保护性物质降低对干旱的敏感性,或产生一系列生理生化变化使干旱解除后各种生理功能够迅速恢复正常,从而减轻干旱的伤害表现出抗逆性[33]。本研究结果表明,干旱胁迫后不同基因型割手密在形态结构上表现绿叶率、株高增长、叶片相对含水量、水势、叶绿素、净光合速率、蒸腾速率、气孔导度均呈下降趋势;在生理生化上表现质膜透性、丙二醛、脯氨酸、超氧化物歧化酶、过氧化物酶均呈上升趋势,这与前人[7-10,34-37]的研究结果一致。
割手密对干旱的适应性和抗御能力不同,表现在生理生化反应上存在相应的差异[3,38],利用抗旱性综合评价能消除单一指标直接判断带来的误差,再结合各材料在形态指标上的表现,使试验结果更准确,更可靠[39]。本试验利用模糊隶属函数法对抗旱性进行综合评价,结果表明,供试材料抗旱性可聚为3种类型,即1份高度抗旱材料,14份中度抗旱材料,11份不抗旱材料,最终筛选出不同抗旱级别的不同基因型割手密种质,为今后甘蔗抗旱生理机制研究和抗旱育种奠定了基础。
在植物的进化过程中,叶片对环境的变化反应较敏感且可塑性较大,因而植物对环境的反应也较多的表现在叶的形态和构造上[40],叶片发育状况和叶面积大小对植物抗逆性形成有很大影响[41]。本研究结果表明,不同基因型割手密的抗旱性与叶面积呈显著负相关(r=0.398,p<0.05),这与许文花等[10]研究结果一致。从割手密叶片形态观察来看,抗旱性强的基因型叶片细小,叶片的叶脉相对较粗,叶脉占叶面积的比值相对较大,叶片的角质化程度也比较高,这样的叶片结构能够降低植株的蒸腾作用,表现出更强的抗旱能力,因此叶片面积可作为割手密抗旱材料筛选的参考依据。
本研究结果表明总体上割手密的染色体数少,叶面积小,其抗旱性表现较强的趋势,例如2013-14、2013-13、2013-10的染色体数最少,均为40条,同时叶面积也最小,2013-13和2013-10的采集地点和海拔又完全一样,因此这类割手密的叶面积与抗旱性呈显著负相关。但并不是所有参试材料都符合这个规律,即割手密材料的抗旱能力和染色体数、叶片面积不是严格意义上的一一对应,例如材料I991-7和2015-56的染色体数最多,均为96条,然而I991-7的叶面积却较小,2015-56的叶片面积也不是最大的,经鉴定I991-7属于不抗旱类型,2015-56属于中度抗旱类型,具体原因有待进一步研究。本研究还发现对材料2012-34等具有匍匐茎[20]的种质,评价结果也可能会有所偏差。
本研究鉴定出的割手密抗旱种质,下一步可与热带种、地方种进行种属间远缘杂交获得创新种质材料,用于扩宽栽培品种的遗传基础,进而选育甘蔗抗旱优良亲本或抗旱新品种。另一方面,随着生物技术和基因工程的广泛应用,从割手密中克隆抗旱基因进行转基因育种创造甘蔗抗旱新品种,是当今甘蔗抗旱育种的努力研究方向。
参考文献
[1] Lü J M, Ju J H, Ren J Z, et al. The influence of the Madden-Julian Oscillation activity anomalies on Yunnan, sextreme drought of 2009-2010[J]. Science China Earth Science, 2012, 55(1): 98-112.
[2] Li Y R. Research and development strategies to improve sugar productivity in China[C]. Proceedings of International Symposium on Technologies to Improve Sugar Productivity in Development Countries, 2006: 7-14.
[3] 陈如凯, 张木清, 陆裔波. 干旱胁迫对甘蔗生理影响的研究[J]. 甘蔗, 1995, 2(1): 1-6.
[4] Hemaprabha G, Nagarajan S R, Adarmelu. Response of sugarcane genotypes to water deficit stress[J]. Sugar Teeh, 2004, 6(3): 165-168.
[5] 何顺长, 杨清辉, 肖凤迥, 等. 全国甘蔗野生种质资源的采集和考察[J]. 甘蔗, 1994, 1(1): 15-22.
[6] Nishiyama M Y J, Ferreira S S, Tang P Z, et al. Full-length enriched cDNA libraries and ORFeome analysis of sugarcane hybrid and ancestor genotypes[J]. PLoS One, 2014, 9(9): e107351.
[7] 经艳芬, 桃联安, 杨李和, 等. 云南野生甘蔗种质血缘F3代亲本材料抗旱性研究初报[J]. 甘蔗, 2002, 9(1): 19-23.
[8] 陈义强, 邓祖湖, 郭春芳, 等. 甘蔗常用亲本及其衍生品种的抗旱性评价[J]. 中国农业科学, 2007, 40(6): 1 108-1 117.
[9] 桃联安, 杨李和, 经艳芬, 等. 云南割手密血缘F2代抗旱性隶属函数法综合评价[J]. 西南农业学报, 2011, 24(5): 1 676-1 680.
[10] 许文花, 杨清辉. 甘蔗割手密无性系抗旱性鉴定[J]. 亚热带农业研究, 2005, 1(1): 22-26.
[11] 杨李和, 桃联安, 经艳芬, 等. 云南野生甘蔗抗旱性遗传表现分析[J]. 中国糖料, 2008(4): 10-13.
[12] 陈能武, 杨荣仲, 吴才文, 等. 四川割手密(S. spontaneum)资源的杂交育种利用潜力研究[J]. 甘蔗, 1996, 3(4): 1-7.
[13] 边 芯, 董立华, 孙有芳, 等. 云南割手密及其血缘F1代材料抗旱相关性状的主成分分析[J]. 干旱地区农业研究, 2014, 32(3): 56-61.
[14] 张木清, 王华忠, 白 晨. 糖料作物遗传改良与高效育种[M]. 北京: 中国农业出版社, 2006.
[15] 刘新龙, 苏火生, 应雄美, 等. 中国十倍体割手密资源的表型相关性及遗传多样性[J]. 湖南农业大学学报(自然科学版), 2012, 38(6): 574-579.
[16] 杨清辉, 何顺长. 云南割手密染色体数目及其地理分布研究[J]. 甘蔗, 1996, 3(1): 10-13.
[17] 王先宏, 郭 超, 娄红波, 等. 不同基因型割手密无性系的核型分析[J]. 植物遗传资源学报, 2014, 15(4): 912-17.
[18] 杨清辉, 李富生, 肖风回. 割手密RAPD指纹图谱分析[J]. 云南农业大学学报, 1998, 13(4): 348-350.
[19] 杨清辉, 萧凤迥, 张发春, 等. 西藏甘蔗亚族植物种质资源考察采集报告[J]. 云南农业大学学报, 1991, 6(3): 183-186.
[20] 胡存脉, 王先宏, 余兴华, 等. 藏南甘蔗野生种质资源的考察与植物学性状分析[J]. 云南农业大学学报, 2015, 30(3): 351-356.
[21] Hsiao T C. Plant responses to water stress[J]. Ann Rev Plant Physiol, 1973, 24: 519-570.
[22] Zhang L D, Zhang L X, Sun J L, et al. Rubisco gene expression and photosynthetic characteristics of cucumber seedlings in Response to water deficit[J]. Scientia Horticulturae, 2013, 161: 81-87.
[23] 程智慧, 孟焕文, Stephen A Rolfe, 等. 水分胁迫对番茄叶片气孔传导及光合色素的影响[J]. 西北农林科技大学学报(自然科学版), 2002, 30(6): 93-96.
[24] Health R L, Packer L. Photoperoxidation in isolated chloroplasts. I. kinetics and stoichiometry of fatty acid peroxidation[J]. Archives of Biochemistry and Biophysics, 1968, 12(5): 189-198.
[25] 陳福明, 陈顺伟. 混合液法测定叶绿素含量的研究[J]. 浙江林业科技, 1984, 4(1): 19-23.
[26] Giannoplitis C N, Ries S K. Superoxide dismutase I. purification and quantitative relationship with water-soluble protein in seedling[J]. Plant Physiology, 1987, 59: 315-318.
[27] 李杨瑞. 甘蔗组织中过氧化物酶活性及其工艺成熟的关系初探[J]. 广西农学院学报, 1990, 9(1): 13-18.
[28] Latef A A H A, He C X. Arbuscular mycorrhizal influence on growth, photosynthetic pigments, osmotic adjustment and oxidative stress in tomato plants subjected to low temperature stress[J]. Acta Physiologiae Plantarum, 2011, 33: 1 217-1 225.
[29] 上海植物生理协会. 植物生理学实验手册[M]. 上海: 上海科技文化出版社, 1985.
[30] 蔡泽霖. 割手密抗旱生理研究[J]. 甘蔗糖业, 1992(5): 16-20.
[31] 揭雨成, 黄呸生, 李宗道. 苎麻品种抗旱性早期鉴定研究[J]. 中国麻作, 1999, 21(3): 21-25.
[32] Rana V, Singh D, Dhiman R, et al. Evaluation of drought tolerance among elite Indian bread wheat cultivars[J]. Cereal Research Communications, 2014, 42: 91-101.
[33] 毕黎明, 刘伟丽, 李杨瑞. 甘蔗抗旱性研究进展与展望[J]. 广西农业科学, 2006, 37(5): 522-526.
[34] 张木清, 陈如凯, 高三基, 等. 甘蔗基因型对水分胁迫的形态生理响应[J]. 中国农业科学, 1997, 30(6): 72-77.
[35] 高三基, 罗 俊, 张 华, 等. 甘蔗抗旱性生理生化鉴定指标[J]. 应用生态学报, 2006(6): 1 051-1 054.
[36] 姚艷丽, 邢淑莲, 徐 磊, 等. 水分胁迫对甘蔗、 割手密和斑茅抗氧化酶活性的影响[J]. 甘蔗糖业, 2013(6): 1-4.
[37] 姚艳丽, 刘 洋, 苏俊波, 等. 干旱胁迫对甘蔗品种及野生近缘材料光合特性及水分利用率的影响[J]. 甘蔗糖业, 2013(1): 14-18.
[38] 夏红明, 陈学宽, 蔡 青, 等. 甘蔗优异材料的抗旱性研究[J]. 亚热带农业研究, 2005, 1(1): 17-21.
[39] 高三基, 罗 俊, 陈如凯, 等. 甘蔗品种抗旱性光合生理指标及其综合评价[J]. 作物学报, 2002, 28(1): 94-98.
[40] Carroll A B, Pallardy S G, Galen C. Drought stress plant water status and floral trait expression in fireweed, Epilobium Angustifoliun(Onagraceae)[J]. American Journal of Botany, 2001, 88(3): 438-446.
[41] Carmo-Silva A E, Francisco A, Powers S J, et al. Grasses of different C4 subtypes reveal leaf traits related to drought tolerance in their natural habitats: Changes in structure, water potential, and amino acid content[J]. American Journal of Botany, 2009, 96(7): 1 222-1 235.
