外源NO对低温胁迫下龙眼幼苗生理特性的影响
2017-05-30高俊杰
摘 要 研究外源一氧化氮(NO)对5 ℃低温胁迫下龙眼幼苗生理特性的影响。结果表明:低温胁迫下,龙眼幼苗叶片中的SOD、POD和CAT活性降低,叶绿素、可溶性糖和蛋白质含量下降,丙二醛含量升高。喷施硝普钠(SNP、NO供体)显著提高龙眼幼苗的叶绿素、可溶性糖和蛋白质含量,提高净光合速率,增强SOD、POD和CAT活性,降低丙二醛含量和电解质渗透率;其中150 μmol/L SNP处理效果最好。可见,外源NO通过刺激抗氧化酶活性来清除活性氧,减轻膜脂过氧化作用,缓解低温胁迫对龙眼幼苗的伤害作用,提高龙眼的抗低温胁迫能力。
关键词 一氧化氮;硝普钠;低温胁迫;龙眼;生理特性
中图分类号 S667.2 文献标识码 A
Effects of Exogenous Nitric Oxide on Physiological Characteristics in Longan Seedling under Low Temperature Stress
GAO Junjie
Planting Station, Quanzhou Municipal Bureau of Agriculture, Quanzhou, Fujian 362000, China
Abstract The effects of sodium nitropprusside(SNP)as an exogenous nitric oxide donor at different concentrations on the physiological reaction of longan seedlings under low temperature(5 ℃)stress were studied. Under the stress of low temperature, superoxide dismutase(SOD), peroxidase(POD), and catalase(CAT)activities as well as the total chlorophyll, soluble protein, and soluble sugar contents of the leaf decreased obviously, and the leaf malondialdedyde content had a remarkable increase. The SOD, POD, and CAT activities and the total chlorophyll, soluble protein, and soluble sugar contents were improved significantly after applying SNP, the malondialdedyde content was reduced. The best abating effect occurred with the SNP concentration 150 μmol/L. It was suggested that exogenous nitric oxides could alleviate the toxic effect of low temperature stress on longan seedlings via activating the seedlings leaf antioxidant enzyme activities and reducing oxidative stress,
Key words Nitric oxide(NO); sodium nitroprusside(SNP); low temperature stress; Dimocarpus longana cv. Fuyan; physiological effects
doi 10.3969/j.issn.1000-2561.2017.09.009
龙眼(Dimocarpus longan Lour.)是典型的亚热带常绿果树,主要分布在广东、广西、福建、海南等省(区);福建省为“中国龙眼之乡”,种植面积已超过6万hm2,占全国龙眼种植总面积的26%[1]。龙眼喜温忌冻,耐寒性差;幼果发育期和反季节栽培的开花期,遇到低温天气,果实生长发育会出现生理障碍,严重时引发大量落果[2]。福建地处东南沿海,寒潮天气和周期性冻害偶有发生,这将给龙眼生产造成严重经济损失。因此,研究龙眼低温胁迫的缓解措施为促进龙眼产业可持续发展有重要意义。
一氧化氮(Nitric oxide,NO)作为一种新的植物生长调节信号分子,参与植物逆境响应和抗病防御反应等过程。NO作为防御响应中的关键信使,在缓解高温胁迫[3]、低温胁迫[4]、酸雨胁迫[5-6]、干旱胁迫[7]和重金属胁迫[8]等非生物脅迫起到重要作用。NO作为信号分子在植物抗逆性中的作用越来越受到重视。目前,龙眼耐寒性的研究主要集中在叶片组织细胞结构特性、成花诱导、生理生化特性和基因表达[9-11]影响等方面,有关龙眼低温胁迫下的缓解作用尚未见报道。本文开展低温胁迫下外源NO缓解龙眼叶片生理特性的变化研究,为龙眼抗寒性和耐寒机理研究提供理论依据。
1 材料与方法
1.1 材料
供试的龙眼品种为主栽中熟品种‘福眼(Dimocarpus longana‘Fuyan),果实由泉州市帝福生态农业有限公司提供。2015年8月进行种子湿沙催芽,待种子萌芽长出1片真叶后种植于塑料盆中,种植盆规格为直径26 cm×高28 cm的圆形塑料盆,基质为蛭石与营养土质量比为2︰1,每盆种植5株龙眼幼苗。龙眼在自然条件下培养至长出5片真叶(株高35~40 cm×茎粗3.5~4.0 cm)进行试验,试验期间按常规进行水肥管理。
1.2 方法
1.2.1 試验设计 2016年5月选择长势基本一致的健康龙眼幼苗进行盆栽试验。低温处理前叶片喷施硝普钠(Sodium nitropprusside , SNP)溶液,浓度分别为0、50、100、150、200 μmol/L,共设5个处理,每个处理3个重复,每盆为1个重复,随机区组设计;每隔 24 h 喷施1次,共喷施3次,每次喷至所有叶片滴液为止。SNP处理后置于人工气候箱进行昼夜低温(5 ℃/5 ℃,12 h)培养,以喷施蒸馏水且正常温度(25 ℃/18 ℃,12 h)培养为对照组(CK)。第3次SNP处理后培养3 d,采集植株上部倒数第2片功能叶放入冰盒,带回实验室,保存于-80 ℃冰箱中用于各项生理指标的测定。
1.2.2 测定项目与方法 丙二醛含量测定采用硫代巴比妥酸反应法[12];电导测定仪测定相对电导率[12]。超氧化物歧化酶活性测定采用氧化硝基四氮唑蓝(NBT)法[13],以单位时间内抑制NBT光还原50%为一个酶活性单位(U);愈创木酚比色法测定过氧化物酶活性[13],以每分钟A470变化值表示酶活性大小;过氧化氢酶活性测定采用比色法[12],以每分钟A240的变化值表示酶活力大小。可溶性糖含量测定采用蒽酮比色法[12],蛋白质含量测定采用考马斯亮蓝法[12];叶绿素含量和类胡萝卜素含量采用乙醇浸提法测定[12]。叶片净光合速率采用CI-340便携式光合仪进行测定[14]。
1.3 数据统计分析
采用Excel 2016统计软件进行数据整理,采用SPSS 22.0统计软件进行数据统计分析,采用单因素方差分析(one-way ANOVA)和Duncan法进行多重比较及差异显著性分析(α=0.05)。
2 结果与分析
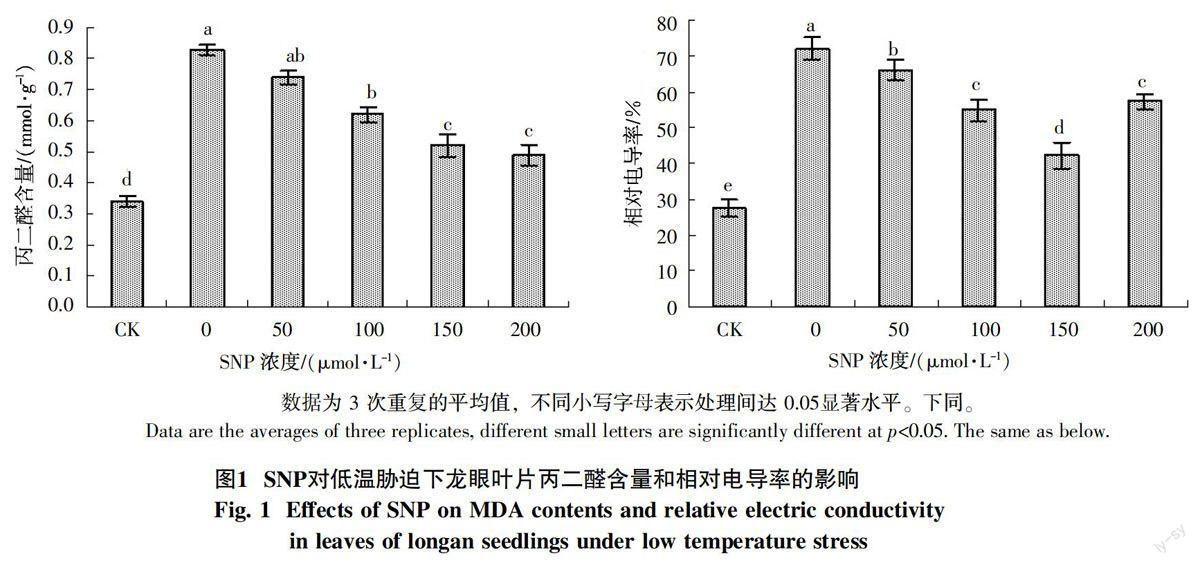
2.1 外源NO对低温胁迫下龙眼苗期丙二醛含量和相对电导率的影响
低温胁迫下龙眼叶片喷施SNP后丙二醛(MDA)含量和相对电导率发生变化(图1)。由图1可见,低温胁迫下(0 μmol/L SNP)丙二醛含量均显著高于对照,比对照增加143.5%,表明低温对龙眼产生伤害。低温胁迫下喷施SNP显著降低丙二醛含量,随着SNP浓度的增加,丙二醛含量逐渐降低;200 μmol/L SNP处理后MDA含量降至最低,比低温胁迫(0 μmol/L SNP)降低了40.9%,仍显著高于对照处理CK。因此,喷施SNP可以显著降低低温胁迫下龙眼丙二醛含量,减轻膜脂过氧化程度。
低温胁迫下细胞的质膜系统受到伤害,表现为电解质渗漏和相对电导率的增加。由图1可知,低温胁迫下龙眼的相对电导率均显著高于对照,喷施SNP降低相对电导率;相对电导率随着SNP浓度的增加表现为先下降后上升的变化。150 μmol/L SNP处理后相对电导率含量降为最低,比低温胁迫降低41.3%;高于对照处理54.0%。可见,低温胁迫条件下SNP可缓解细胞质膜的损伤,保护细胞的质膜透性和完整性,提高龙眼叶片的抗逆性。
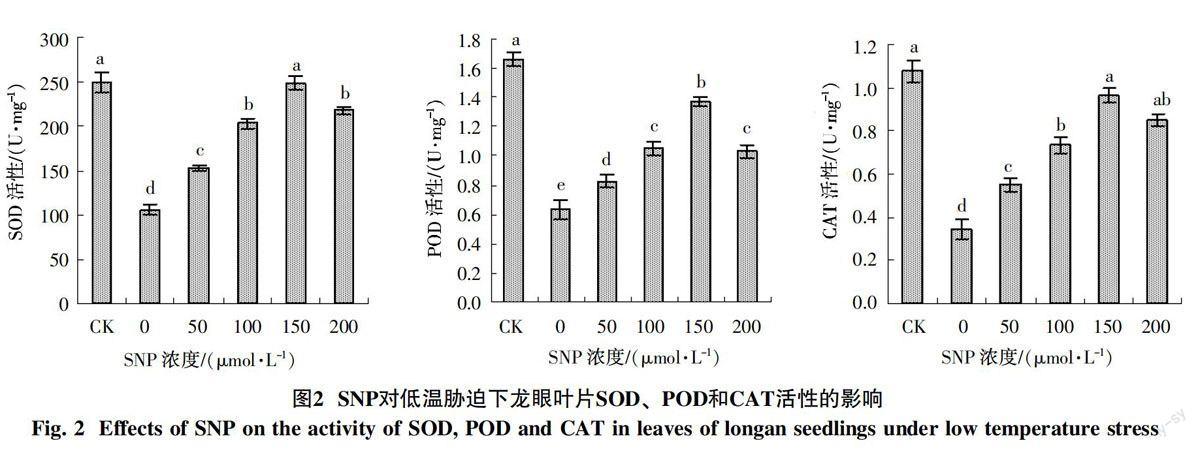
2.2 外源NO对低温胁迫下龙眼苗期抗氧化酶活性的影响
由图2可知,低温胁迫(0 μmol/L SNP)显著降低龙眼叶片的SOD、POD和CAT 3种酶活性。低温胁迫下喷施SNP显著提高龙眼叶片的SOD、POD和CAT活性,随着SNP浓度的增加酶活性出现先增加后降低的变化;150 μmol/L SNP处理后,SOD、POD和CAT活性均达到最大值,比低温胁迫(0 μmol/L SNP)分别提高136.2%、116.4%和180.8%,但SOD和CAT活性与对照处理差异不显著;200 μmol/L SNP处理后,SOD、POD和CAT活性均出现降低,仍高于对照处理。可见,喷施SNP显著提高低温胁迫下SOD、POD和CAT活性,150 μmol/L SNP有利于提高低温胁迫下龙眼叶片抗氧化酶活性,增强清除活性氧的能力。
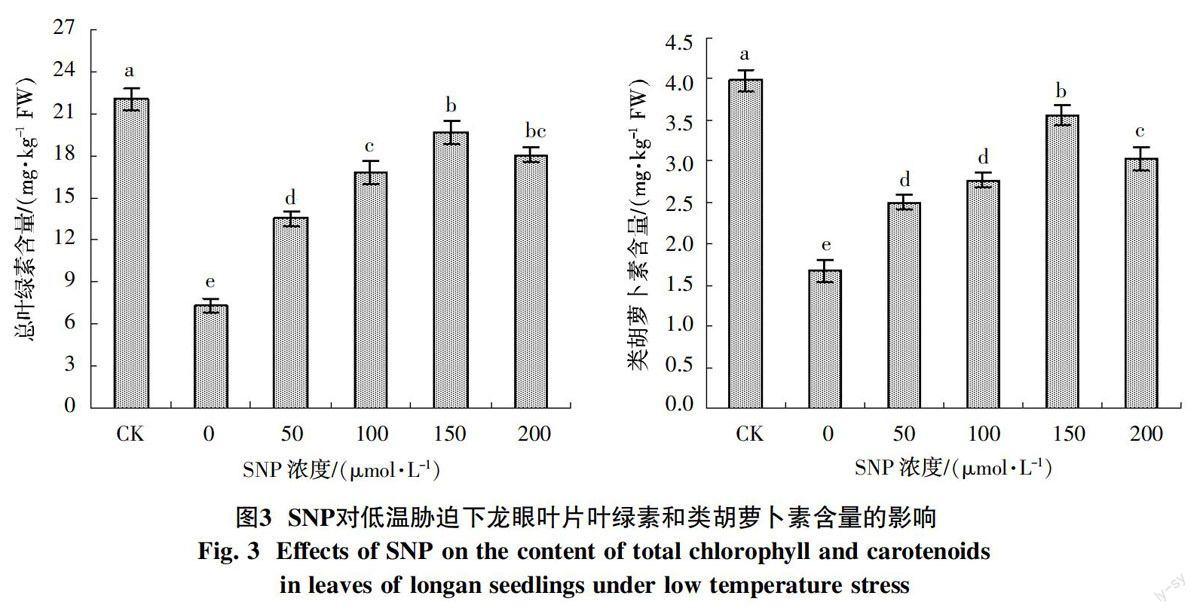
2.3 外源NO对低温胁迫下龙眼苗期光合色素含量的影响
低温胁迫显著降低龙眼叶片的总叶绿素和类胡萝卜素含量(图3),分别比对照处理降低66.9%和58.0%;喷施SNP显著提高低温胁迫下龙眼的总叶绿素和类胡萝卜素含量。由图3可知,低温胁迫下总叶绿素和类胡萝卜素含量随着喷施SNP浓度的升高而表现出先逐渐增加后降低的变化,150 μmol/L SNP处理后总叶绿素和类胡萝卜素含量均达到最大值,分别比低温胁迫(0 μmol/L SNP)处理提高170.5%和112.6%;200 μmol/L SNP处理后其含量反而出现降低,但显著高于低温处理(0 μmol/L SNP)。这表明SNP处理可以缓解低温对叶绿体结构的破坏,提高龙眼对低温胁迫的抵抗能力。
2.4 外源NO对低温胁迫下龙眼苗期叶片净光合速率的影响
低温胁迫显著降低龙眼叶片净光合速率,喷施SNP缓解低温胁迫的伤害(图4)。从图4可见,低温胁迫下净光合速率比对照处理降低53.3%,低温胁迫下喷施SNP后净光合速率随着SNP浓度的增加而呈现出先提高后降低,但显著高于低温胁迫(0 μmol/L SNP)。SNP浓度为150 μmol/L时叶片的净光合速率达到最大值,比低温胁迫(0 μmol/L SNP)提高86.7%;200 μmol/L SNP处理后,其含量反而出现降低,但显著高于低温处理(0 μmol/L SNP)。可见,喷施150 μmol/L SNP有利于提高低温胁迫下龙眼的光合能力。
2.5 外源NO对低温胁迫下龙眼苗期可溶性糖和蛋白质含量的影响
低温胁迫显著降低龙眼叶片可溶性糖和蛋白质含量,喷施SNP显著延缓龙眼叶片可溶性糖和蛋白质含量的下降(图5)。从图5可以看出,低温胁迫下可溶性糖和蛋白质含量分别比对照处理降低49.5%和48.3%;随着SNP浓度的增加,龙眼叶片的可溶性糖和蛋白质含量均表现为先升高而后降低的变化。SNP浓度为150 μmol/L时其含量均达到最大值,分别比对照提高88.0%和84.2%;200 μmol/L SNP处理后可溶性糖和蛋白质含量均下降,但显著高于低温处理(0 μmol/L SNP)。因此,喷施150 μmol/L SNP有利于提高低温胁迫下龙眼光合产物的合成,从而提高龙眼的抗寒性。
3 討论
抗氧化酶活性与植物活性氧代谢直接相关,SOD、POD和CAT是植物体内的重要活性氧清除系统,其活性水平代表植物体清除活性氧自由基的能力,反映植物抗逆境能力的高低。NO 作为信号调节分子和活性氧清除剂能调节植物对非生物胁迫的适应反应,显著提高叶片可溶性糖和可溶性蛋白含量,并可有效提高SOD、POD和CAT等保护酶的活性,增强抗逆能力[15]。樊怀福等[16]研究表明,SNP处理显著降低黄瓜幼苗叶片细胞膜的离子渗漏和MDA含量,缓和膜相变化,缓解膜透性的增大,可防止离子渗漏;可溶性蛋白是植物体内重要的渗透调节物质和保护物质。已有研究表明,外源NO可以增加胁迫下棉花[17]、香蕉[18]和苦瓜[19]幼苗叶片的可溶性蛋白含量。姜义宝等[20]发现0.1 mmol/L SNP减缓干旱胁迫下苜蓿幼苗细胞膜透性的增加,使CAT先升后降,SOD和POD活性持续升高;SNP处理黑麦草,可提高黑麦草SOD、POD和CAT等保护酶活性[21]。本研究表明,龙眼喷施低浓度SNP溶液,提高叶片中可溶性糖和蛋白质含量,SOD、POD和CAT 3种酶活性均得到不同程度的提高。可溶性糖和可溶性蛋白作为渗透调节物质,可提高低温胁迫下细胞液浓度,增加细胞保水能力,维持植株渗透势、抗脱水,从而提高植物的抗冷性[19];同时,NO能提高低温胁迫下龙眼的抗氧化酶活性,有利于清除低温胁迫诱导生产的活性氧,以及抑制脂氧合酶活力,降低超氧阴离子的产生速度和质膜相对透性从而降低丙二醛(MDA)的积累,外源NO对细胞膜具有良好的修复和保护作用,减缓膜脂过氧化程度,从而提高龙眼的抗性。外源NO对植物的作用具有双重性。高浓度对生物体有毒害作用,低浓度缓解胁迫效应[22]。这与NO缓解其他非生物胁迫的作用机制相似[23-25]。高浓度SNP处理降低龙眼低温胁迫的缓解作用,可能是高浓度NO与O2-相互作用生成大量的过氧亚硝酸根阴离子,后者经质子化后形成具有强氧化性的过氧亚硝酸,破坏生物大分子的结构与功能,表现出其生物毒性。
叶绿体是植物进行光合作用的细胞器,温度是影响叶绿体发育的重要因素;叶绿体对低温弱光反应最敏感。低温胁迫下植物叶片光合活性的显著下降与叶绿体超微结构有关。低温胁迫显著降低叶片叶绿素和类胡萝卜素含量,可能是低温降低生物合成过程酶活性,引起叶绿素合成系统受阻,以及低温胁迫下叶绿素受到破坏[1-2]。已有研究表明,外源NO能提高低温胁迫下玉米[22]、棉花[17]、黄瓜[16]、苦瓜[19]叶片的叶绿素a、叶绿素b、总叶绿素含量。本研究表明,外源NO提高低温胁迫下龙眼叶绿素和类胡萝卜素含量,这可能是NO激活龙眼叶绿素生物合成过程中的某些酶类[26];而且NO具有强氧化性的活性氮,能够促进低温胁迫下植物类囊体膜蛋白复合体的组装和稳定,增强叶绿体对光能的吸收和利用能力,缓解低温胁迫导致的叶绿体破坏,提高龙眼幼苗抗冷害能力[19]。同时,低温胁迫下龙眼幼苗的净光合速率明显下降,光合产物可溶性糖含量也显著降低[27];150 μmol/L SNP显著提高低温胁迫下龙眼幼苗的净光合速率,说明外源NO显著缓解低温胁迫造成的叶肉细胞光合活性的下降;从而缓解同化物可溶性糖和蛋白质含量的下降[17]。
综上所述,外源NO能显著缓解低温胁迫对龙眼幼苗生长造成的抑制作用,提高抗氧化酶活性,降低幼苗叶片的膜脂过氧化作用,减少低温对膜造成的伤害,提高龙眼苗期抗低温胁迫能力。NO作为一种信号分子参与植物体内非常复杂的生理过程,其缓解低温胁迫的机理尚需要进一步的研究。
参考文献
[1] 刘建福, 钟书淳, 王奇志, 等. 龙眼叶片对低温胁迫的生理响应研究[J]. 江西农业大学学报, 2014,36(4):737-742.
[2] 郑重禄. 龙眼低温胁迫及其抗寒性生理[J]. 中国南方果树, 2012, 41(3): 64-71.
[3] 李 秀, 巩 彪, 徐 坤. 外源NO对高温胁迫下姜叶片活性氧代谢的影响[J]. 园艺学报, 2014, 41(2): 277-284.
[4] 相 昆, 徐 颖, 李国田, 等. 外源NO对低温胁迫下核桃幼苗活性氧代谢的影响[J]. 林业科学, 2016, 52(1): 143-149.
[5] 刘建福, 钟书淳, 王奇志, 等. NO供体SNP对酸雨胁迫下龙眼种子萌发及生理特性的影响[J]. 江苏农业学报, 2013, 29(5): 1 143-1 148.
[6] 高俊杰. 多效唑对酸雨胁迫下龙眼幼苗生理特性的影响[J]. 安徽农业科学, 2016, 44(2):4 8-50, 53.
[7] 邵瑞鑫, 李蕾蕾, 郑会芳, 等. 外源一氧化氮对干旱胁迫下玉米幼苗光合作用的影响[J]. 中国农业科学, 2016, 49(2): 251-259.
[8] 夏海威, 施国新, 黄 敏, 等. 一氧化氮对植物重金属胁迫抗性影响的研究进展[J]. 生态学报, 2015, 35(10): 3 139-3 147.
[9] 张惠斌, 刘星辉. 龙眼叶片组织细胞结构特性与耐寒性关系[J]. 园艺学报, 1993, 20(1): 1-7.
[10] 姜建初, 裴铁雄, 王静静, 等. 低温冷害对龙眼成花的影响及抹除荔枝和芒果冷害花穗促进腋芽再生花穗的效果[J]. 广西植物, 2010, 30(2): 220-223.
[11] 陈 虎, 何新华, 罗 聪, 等. 低温胁迫下龙眼碳酸酐酶基因(CA)的克隆与表达分析[J]. 园艺学报, 2012, 39(2): 243-252.
[12] 邹 琦. 植物生理学实验指导[M]. 北京: 中国农业出版社, 2000.
[13] 王学奎. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社, 2006.
[14] 刘建福, 钟书淳, 黄安民, 等. 武夷岩茶光合特性及品质成分比较[J]. 热带作物学报, 2014, 35(6): 1 148-1 152.
[15] 郭经纬, 商 桑, 穆大伟, 等. 外源NO对低温胁迫下豇豆幼苗生长和生理特性的影响[J]. 热带作物学报, 2015, 36(12): 2 179 -2 183.
[16] 樊怀福, 杜长霞, 朱祝军. 外源NO对低温胁迫下黄瓜幼苗生长、叶片膜脂过氧化和光合作用的影响[J]. 浙江农业学报, 2011, 23(3): 538-542.
[17] 杨美森, 王雅芳, 干秀霞, 等. 外源一氧化氮对冷害胁迫下棉花幼苗生长、抗氧化系统和光合特性的影响[J]. 中国农业科学, 2012, 45(15): 3 058-3 067.
[18] 汤红玲, 李 江, 陈惠萍. 外源一氧化氮对香蕉幼苗抗冷性的影响[J]. 西北植物学报, 2010, 30(10): 2 028-2 033.
[19] 杜卓涛, 商 桑, 朱白婢, 等. 外源NO对低温胁迫下苦瓜幼苗生长和几个生理指标的影响[J]. 热带作物学报, 2016, 37(3): 482-487.
[20] 姜义宝, 杨玉荣, 郑秋红. 外源一氧化氮对干旱胁迫下苜蓿幼苗抗氧化酶活性和叶绿素荧光特性的影响[J]. 干旱地区农业研究, 2008, 26(2): 65-68.
[21] 张远兵, 刘爱荣, 方 蓉. 外源一氧化氮对镉胁迫下黑麦草生长和抗氧化酶活性的影响[J]. 草业学报, 2008, 17(4): 57-64.
[22] 陈银萍, 王晓梅, 杨宗娟, 等. NO对低温胁迫下玉米种子萌发及幼苗生理特性的影响[J]. 农业环境科学学报, 2012, 31(2):270-277.
[23] Bailly C, Benamar A, Corbineau F, et al. Changes in malondialdehyde content and in superoxide dismutase, catalase and glutathione reductase activities in sunflower seeds as related to deterioration during accelerated aging[J]. Physiologia Plantarum, 2012, 97(1): 104-110.
[24] 吳锦程, 陈伟建, 蔡丽琴, 等. 外源NO对低温胁迫下批把幼果抗氧化能力的影响[J]. 林业科学, 2010, 46(9): 73-78.
[25] Li X, Gong B, Xu K. Interaction of nitric oxide and ployamines involves antioxidants and physiological strategies against chilling induced oxidative damage in Zingiber officinale Roscoe[J]. Scientia Horticulturae, 2014, 170(7): 237-248.
[26] 刘建福, 王明元, 杨 晨, 等. 外源NO对酸雨胁迫下龙眼幼苗生理特性的影响[J]. 应用生态学报, 2013, 24(8): 2 235-2 240.
[27] Palma F, Carvajal F, Lluch C, et al. Changes in carbohydrate content in zucchini fruit(Cucurbita pepo L.) under low temperature stress[J]. Plant Science, 2014, 217-218: 78-86.
