相位同步讨论哺乳动物的昼夜节律
2017-05-24樊庆端刘曾荣
樊庆端,刘曾荣
(1.上海工程技术大学基础教学学院,上海 201620; 2.上海大学系统生物技术研究所,上海 200444; 3.上海大学通信与信息工程学院,上海 200444)
相位同步讨论哺乳动物的昼夜节律
樊庆端1,2,刘曾荣1,3
(1.上海工程技术大学基础教学学院,上海 201620; 2.上海大学系统生物技术研究所,上海 200444; 3.上海大学通信与信息工程学院,上海 200444)
实验表明哺乳动物视交叉上核(suprachiasmatic nucleus,SCN)神经元集体涌现出的节律控制着中央神经中枢和外围组织,从而形成哺乳动物的昼夜节律.为了探索昼夜节律的涌现机制,根据已有实验结果构建了SCN神经元耦合网络,并建立了描述昼夜节律的修正的Kuramoto模型.研究发现,昼夜节律的涌现是通过相位同步来表达的.另外,讨论了模型参数对描述相位同步的序参数的影响,其结果与生物学讨论吻合.该模型为解释昼夜节律的产生机制提供了一个新思路.
昼夜节律;视交叉上核(suprachiasmatic nucleus,SCN);Kuramoto模型;相位同步
昼夜节律是哺乳动物的一种十分常见现象.从生物学的角度来看,该现象与位于下丘脑的视交叉上核(suprachiasmatic nucleus,SCN)神经元中的蛋白表达及其活性是否与外界环境(主要是光照)的周期变化一致有关.神经节律的失调会严重影响患者的生活质量,导致疾病产生,如睡眠障碍[1]、精神分裂[2-3]等.该现象与SCN细胞的周期行为是否与地球自转的周期行为实现周期和相位同步有关.按此思路,研究者们开始从分子层次上进行昼夜节律的研究工作.已有学者通过实验观察到SCN细胞中PERIOD(PER1,2,3),FRQENCY (FRQ1,2)和CRYPTOCHROME(CRY1,2)等时钟蛋白的表达具有振荡行为,这吸引了很多学者运用转录-翻译反馈环探讨昼夜节律行为.1965年,Goodwin[4]运用3个变量分别表示时钟基因的mRNA、蛋白和相关转录抑制子,构建了一个转录-翻译反馈环模型.2003年, Leloup等[5]构建的19个方程组成的模型中,变量为主要时钟基因PER,CRY,CRY/PER二聚体,BMAL1,CLOCK/BMAL1二聚体的mRNA,蛋白的磷酸化与非磷酸化状态以及REVERBα的mRNA和蛋白浓度.同年,Forger等[6]进一步建立了含有73个变量的微分方程模型,该模型再现了SCN神经元的昼夜节律行为.Gonze等[7]在Goodwin模型的基础上建立了网络上的转录-翻译的反馈环动力学模型,并分析了耦合的影响.然而,上述模型很难用来直接讨论SCN中所有神经元与地球自转产生的周期与相同步.近年来,Li等[8]指出了SCN神经元之间耦合的重要性,并运用Goodwin模型分析了耦合诱导的同步机制.Gu等[9]通过Poincare模型分析了耦合强度的分布对神经元的内在周期及其同步的影响.Ullner等[10]运用全连接网络上的Goodwin模型分析了光照强度与耦合强度对昼夜节律的影响.这些模型通常应用于周期同步的分析,而直接用来分析相同步的模型相对较少.
目前,采用Kuramoto模型讨论相同步的工作进展较大.Blanter等[11]讨论了两个振子的同步行为.Gu等[12]讨论了SCN中神经元振子的相位分离现象.近年来,已有学者采用Kuramoto模型研究了复杂网络上中振子的周期和相位同步[13-16].这表明Kuramoto模型可用于讨论大量基本单位组成系统的周期和相位同步.
基于上述研究,本工作考虑SCN的结构和功能特点,构造了描述SCN所有神经元整体行为的修正Kuramoto模型,并用该模型讨论哺乳动物相同步产生昼夜节律的合理性;针对模型中结构参数与动力学参数的影响作进一步讨论,所得结果合理地解释了昼夜节律的一些现象.
1 SCN网络上修正的Kuramoto模型
根据SCN的结构和功能特点,将SCN神经元分为腹外侧(ventral-lateral,VL)和背中侧(dorsal-medial,DM)两部分,并分别构造子网络.然后,通过有向耦合形成一个网络,并在此网络上构建Kuramoto模型.
1.1 SCN网络
首先讨论SCN的结构和功能.SCN包含了大约20 000个自激振荡神经元,在解剖学上SCN的两个子区域VL与DM产生不同的多肽神经元,其中VL中的多肽神经元主要有血管活性肠肽(vasoactive intes polypeptide,VIP),DM中的多肽神经元主要有精氨酸血管加压素(arginine vasopressin,AVP).VL神经元约占SCN的20%~33%[17],具有感光功能且易被外部的周期光卷带,而DM神经元不感光且其在未经VL神经元收到光信号时大多会产生自激振荡行为[18].果蝇的生物钟神经元实验表明:VL通过光信号对DM起调控作用[19];VL神经元耦合紧密呈现周期性振荡,而DM神经元之间的弱耦合几乎可以忽略[20].这表明两个子区域具有不同的结构和功能,因此探讨昼夜节律时应先将SCN神经元划分为VL与DM两部分,分别构建相关子网络,再将二者耦合起来形成一个网络.
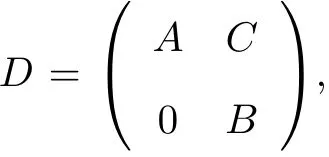
1.2 修正的Kuramoto模型
按照Kuramoto模型的基本方法,可以在SCN网络上构建描述SCN昼夜节律行为的模型,为简单起见,其中的光信号用正弦函数表示,
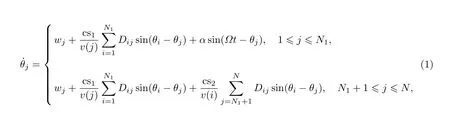
式中:θj与wj分别表示第j个神经元振子的相位与频率;N表示SCN中神经元的数目,N1表示VL中神经元的数目,N2表示DM中神经元的数目;Ω与α分别表示昼夜光信号的频率与强度;v(j)表示影响神经元j的神经元的数目,即矩阵D中第j列不等于0的元素的个数;由于DM内部的耦合较弱,假定VL内部以及VL到DM的耦合强度相等,且与DM内部的耦合不同,参数cs1表示VL内部神经元之间以及VL到DM神经元的耦合强度,cs2表示DM内部神经元之间的耦合强度.
2 相位同步
2.1 神经元相位在光照下的演化
在昼夜节律系统中,神经元的相位可以被外界信号卷吸到一起,神经元的耦合结构与耦合强度以及光照强度对系统的同步影响较大.为了说明这些现象,以400个SCN神经元为例,运用修正Kuramoto模型讨论SCN中细胞的相同步.
生物实验表明,SCN内神经元的周期分布为20~28 h[23],因此这里假定神经元服从正态分布N(24,1).另外,注意到DM内部的耦合强度较小,故取VL内部耦合强度的1/10.模型中的参数值如表1所示.

表1 SCN网络上修正Kuramoto模型中的参数Table 1 Parameters in the modified Kuramote model on SCN networks
取VL与DM中神经元的相位初值在[0,2π]上均服从均匀分布,它们在周期为24 h、初相位为0的外界光信号作用下演化.对系统在240 h内的相位变化进行数值模拟,选取其中9个具有代表性的相位演化过程,具体如图1所示,其中θ20,θ60与θ100表示VL的3个神经元, θ120,θ160,θ200,θ240,θ280,θ320,θ360与θ400表示DM的8个神经元的相位.从图1中可以看出,SCN神经元逐渐地被周期为24 h的光照所卷吸.大多数神经元都可达到卷吸效果,即基本上与光照达到了周期同步和相位同步.同时也有少数神经元由于与环境耦合较弱,加之固有周期的差别,仍表现为自激振荡.这表明上述模型可以较好地说明哺乳动物可通过SCN神经元与光照的相同步而实现其昼夜节律.
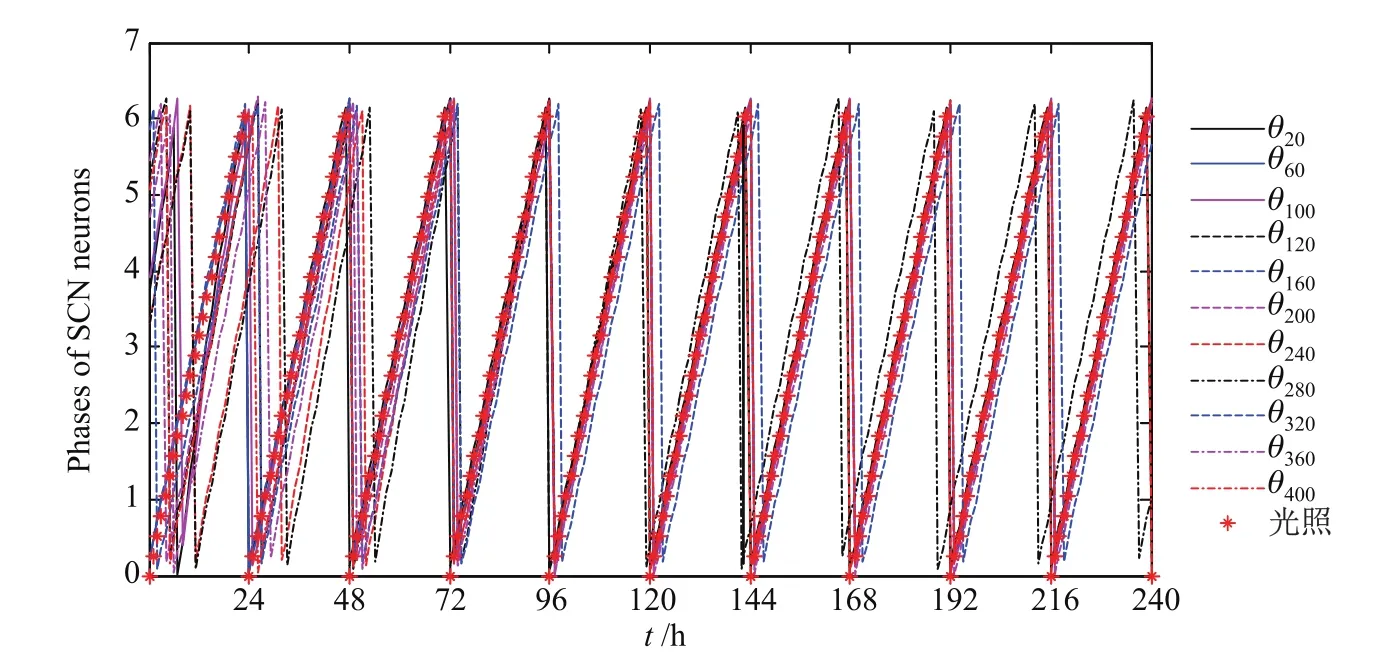
图1 SCN神经元相位与外界周期为24 h光信号同步的过程Fig.1 Processes of phases of SCN neurons’synchronizing to the 24 h cycle light signal
2.2 参数对相位序参数的影响
为进一步讨论模型的合理性,需要研究模型中参数对相同步的影响.由Kuramoto模型讨论可知,可以用序参数来刻画相位的同步程度.若记神经元j的相位为θj,则VL,DM与SCN神经元的相位序参数可分别定义为
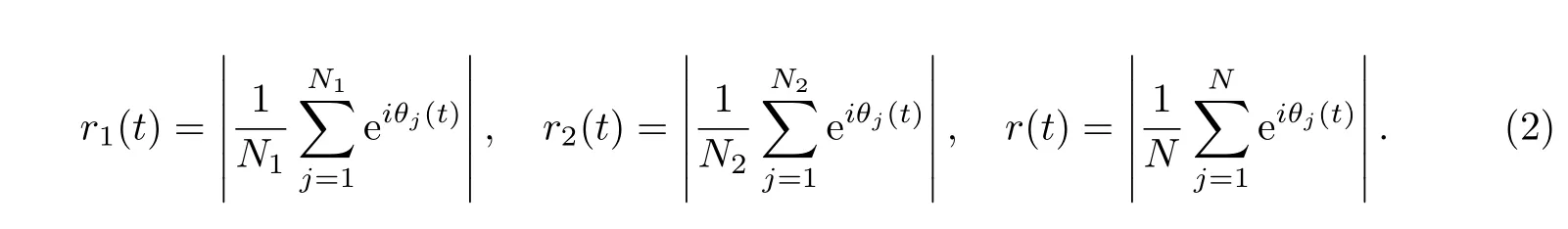
由序参数定义可知,值越接近1表示越多神经元实现了与光照相位同步.表1中参数K 与q分别决定了子区域内部及子区域之间的耦合结构,可见这两个结构参数对SCN神经元的同步能力极为重要.首先,模型中的K依次取为0,2,4,6,8和10(单侧邻居数目为K/2),其余参数与初值同图1,可得240 h内序参数随时间的变化情况(见图2).当K=0时,VL神经元只受光信号驱动,DM神经元仅受与其相连的VL神经元的影响,其相位可以很快达到同步.由图2(a)可以看到,VL神经元的相位序参数短期内都能达到较高的值,而神经元间的连接数目对其影响甚微.图2(b)和(c)表明序参数在初期的增速及其后期的稳定值都随着K的增加而减小,这是一个比较有趣且有意义的现象.当400个相位在[0,π/3](相应时间差为4 h)上均匀取值时,序参数r为0.954 7;当其(400个相位)服从均匀分布时,序参数的多次模拟值均大于0.95.然而,当K=10时,SCN的序参数小于0.9,此时相位处于较差的卷吸状态,其原因很可能是K的增大使SCN网络出现了一些结构紧凑的“小社团”.因此在数值模拟中,取K=6.随着q的增大,DM与VL之间的连接增多,即更多VL的相位信息直接传输到DM,从而使得序参数上升加快.因此q的增大可提高SCN内神经元的同步速度.
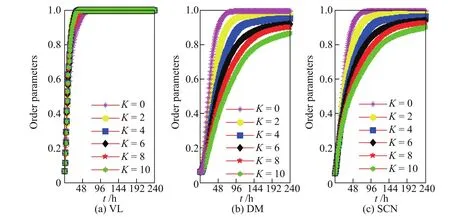
图2 连接度对VL,DM和SCN中神经元相位序参数的影响Fig.2 Effects of mean degree on order parameters of neurons in VL,DM and SCN
光照强度与耦合强度也是影响相位动力学行为的重要因素.光照强度α与耦合强度cs1对序参数的影响分别如图3和4所示.图3(a)表明VL神经元相位很快达到同步,这与神经元受光线直接驱动有关;图3(b)和(c)表明DM以及整个SCN的神经元的序参数随着光照强度的增加而缓慢增大,且都有上限.这说明光照强度有利于节律涌现,但促进水平却有限,这一点与生活实际比较吻合.图4(a)表明耦合强度的增大提高了VL神经元的同步状态,却使得VL神经元间的相位差缩小,从而使SCN神经元的相位更快地实现同步.这充分表明序参数随着耦合的增强而增大.图3与4的比较也说明光照对SCN节律的影响不如神经元间耦合强度的影响大.因此,昼夜节律涌现的主要因素为SCN内部的结构与耦合,外界环境的作用仅仅是利于节律的形成.
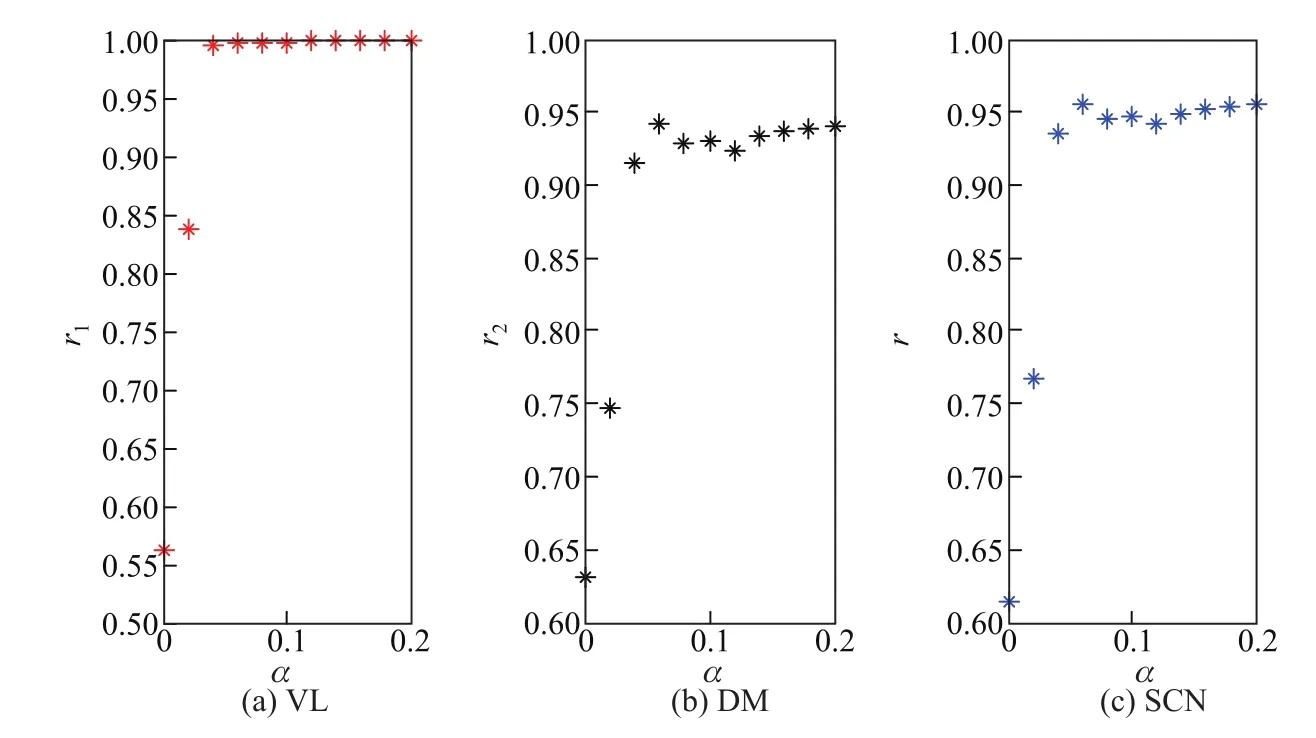
图3 光照强度对VL,DM和SCN中神经元相位序参数的影响Fig.3 Effects of light density on order parameters of neurons in VL,DM and SCN
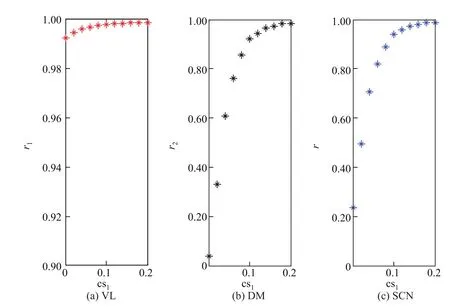
图4 耦合强度对VL,DM和SCN中神经元相位序参数的影响Fig.4 Effects of coupling strength on order parameters of neurons in VL,DM and SCN
3 结束语
哺乳动物的生理、行为等节律主要由视交叉上核中的神经元控制.已有实验和理论研究表明,这些神经元的振荡行为与外界环境(主要是光照)同时具有周期同步和相位同步才能使哺乳动物表现出昼夜节律行为.
一般将SCN的神经元分为VL与DM来研究昼夜节律,因为SCN的子区域结构与功能明显不同.另外,已有的研究中缺乏既考虑SCN的结构特点又能全面分析周期和相位行为的模型.为此,本工作结合VL与DM神经元的功能与结构特点,构建了描述SCN细胞涌现昼夜节律的一个修正的Kuramoto模型,从相位同步的观点解释了昼夜节律的涌现机制,并讨论各个参数对节律涌现的影响.数值结果表明,光照强度与神经元之间耦合强度的增加有利于昼夜节律的涌现,而光照的促进作用比较有限.另外,SCN网络中神经元的平均连接数目的增加可能导致局部出现连接紧密的神经元簇,不利于昼夜节律的涌现.这是一个有趣的结果,有待于进一步研究.通过调控神经元之间的耦合关系及耦合强度可以促进神经节律的涌现,这为节律失调药物的开发提供了新思路.
[1]DAGAN Y.Circadian rhythm sleep disorders(CRSD)[J].Sleep Medicine Reviews,2002,6(1): 45-54.
[2]STEPHENSON R.Circadian rhythms and sleep-related breathing disorders[J].Sleep Medicine, 2007,8(6):681-687.
[3]ROOPUN A K,KRAMER M A,CARRACEDO L M,et al.Period concatenation underlies interactions between gamma and beta rhythms in neocortex[J].Frontiers in Cellular Neuroscience, 2008,2(1):1-8.
[4]GOODWIN B C.Oscillatory behavior in enzymatic control processes[J].Advances in Enzyme Regulation,1965,3:425-438.
[5]LELOUP J C,GOLDBETER A.Toward a detailed computational model for the mammalian circadian clock[J].Proceedings of the National Academy of Sciences,2003,100(12):7051-7056.
[6]FORGERDB,PESKINCS.Adetailedpredictivemodelofthemammalian circadian clock[J].Proceedings of the National Academy of Sciences,2003,100(25):14806-14811.
[7]GONZE D,BERNARD S,WALTERMANN C,et al.Spontaneous synchronization of coupled circadian oscillators[J].Biophysical Journal,2005,89(1):120-129.
[8]LI Y,LIU Z R.Synchronization of clocks coupled by neurotransmitter in the SCN[J].Journal of Biological Systems,2013,21(1):1350006.
[9]GU C G,WANG J X,LIU Z H.Free-running period of neurons in the suprachiasmatic nucleus: its dependence on the distribution of neuronal coupling strengths[J].Physical Review E,2009, 80(3):030904.
[10]ULLNER E,BUCETA J,DIEZ-NOGUERA A,et al.Noise-induced coherence in multicellular circadian clocks[J].Biophysical Journal,2009,96(9):3573-3581.
[11]BLANTER E M,LE MOUEL J L,SHNIRMAN M G,et al.Kuramoto model of nonlinear coupled oscillators as a way for understanding phase synchronization:application to solar and geomagnetic indices[J].Solar Physics,2014,289(11):4309-4333.
[12]GU C G,WANG J X,WANG J,et al.Mechanism of phase splitting in two coupled groups of suprachiasmatic-nucleus neurons[J].Physical Review E,2011,83(4):046224.
[13]ZHANG Y F,XIAO R B.Synchronization of Kuramoto oscillators in small-world networks[J]. Physica A,2014,416(3):33-40.
[14]ARENAS A,DIAZ-GUILERA A.Synchronization and modularity in complex networks[J].The European Physical Journal Special Topics,2007,143(1):19-25.
[15]DORFLER F,BULLO F.Synchronization in complex networks of phase oscillators:a survey[J]. Automatica,2014,50(6):1539-1564.
[16]DAIDO H.Algebraic relaxation of an order parameter in randomly coupled limit-cycle oscillators[J].Physical Review E,2000,61(2):2145-2147.
[17]MEIjER J H,SCHWARTZ W J.In search of the pathways for light-induced pacemaker resetting in the suprachiasmatic nucleus[J].Journal of Biological Rhythms,2003,18(3):235-249.
[18]LI Y,LIU Z,ZHANG J,et al.Synchronisation mechanisms of circadian rhythms in the suprachiasmatic nucleus[J].IET Systems Biology,2009,3(2):100-112.
[19]YAO Z,SHAFER O T.The drosophila circadian clock is a variably coupled network of multiple peptidergic units[J].Science,2014,343(6178):1516-1520.
[20]YAMAGUCHI S,ISEjIMA H,MATSUO T,et al.Synchronization of cellular clocks in the suprachiasmatic nucleus[J].Science,2003,302(5649):1408-1412.
[21]FREEMAN G M,KROCK R M,ATON S J,et al.GABA networks destabilize genetic oscillations in the circadian pacemaker[J].Neuron,2013,78(5):799-806.
[22]HAFNER M,KOEPPL H,GONZE D.Effect of network architecture on synchronization and entrainment properties of the circadian oscillations in the suprachiasmatic nucleus[J].PLoS Computational Biology,2012,8(3):e1002419.
[23]HONMA S,NAKAMURA W,SHIRAKAWA T,et al.Diversity in the circadian periods of single neurons of the rat suprachiasmatic nucleus depends on nuclear structure and intrinsic period[J].Neuroscience Letters,2004,358(3):173-176.
本文彩色版可登陆本刊网站查询:http://www.journal.shu.edu.cn
Circadian rhythm in mammal based on phase synchronization
FAN Qingduan1,2,LIU Zengrong1,3
(1.College of Fundamental Studies,Shanghai University of Engineering Science,Shanghai 201620,China; 2.Institute of Systems Biology,Shanghai University,Shanghai 200444,China; 3.School of Communication and Information Engineering,Shanghai University Shanghai 200444,China)
A collective rhythm emerges in suprachiasmatic nucleus(SCN)of mammals hypothalamus,controlling the central nervous system and peripheral tissues.To explore the emergence of circadian rhythms,a modified Kuramoto model of the SCN network is established,and its phase synchronization is analyzed.The results show that emergence of circadian rhythm is expressed by phase synchronization.The effect of parameters on order parameter is studied,showing agreements with biological facts.This study provides insights of the mechanism of circadian rhythm.
circadian rhythm;suprachiasmatic nucleus(SCN);Kuramoto model;phase synchronization
Q 612;O 29
A
1007-2861(2017)02-0290-08
10.3969/j.issn.1007-2861.2015.02.005
2015-11-07
国家自然科学基金资助项目(10832006,11172158)
刘曾荣(1943—),男,教授,博士生导师,研究方向为系统生物学.E-mail:zrongliu@126.com
