人白细胞介素10受体α基因真核表达及与JAK1蛋白的相互作用的检测
2017-05-19郭欣欣王刚邓巧亭吴涵韬李坤吴英松刘天才
郭欣欣 王刚 邓巧亭 吴涵韬 李坤 吴英松 刘天才
(南方医科大学检验与生物技术学院抗体工程研究所,广州 510515)
人白细胞介素10受体α基因真核表达及与JAK1蛋白的相互作用的检测
郭欣欣 王刚 邓巧亭 吴涵韬 李坤 吴英松 刘天才
(南方医科大学检验与生物技术学院抗体工程研究所,广州 510515)
构建含人白介素10受体α(IL-10RA)基因的真核表达质粒pENTER-IL-10RA-His,并在HEK293中进行真核表达,并用免疫共沉淀检测JAK1与IL10RA在细胞内的相互作用。在HeLa细胞中提取人总RNA,通过RT-PCR获得人IL10RA的基因全长,并将其克隆至真核表达载体pENTER-His中。经PCR扩增、双酶切、测序鉴定后,将重组质粒pENTER-IL-10RA-His转染至HEK293细胞中。免疫印迹法检测IL-10RA蛋白在HEK293细胞中的表达。结果显示,经PCR扩增和双酶切,测序鉴定质粒克隆正确。免疫印迹可见63 kD的目的蛋白。共同转染JAK1和IL-10RA的质粒,免疫印迹可见分别为133 kD和63 kD的目的条带,免疫共沉淀鉴定了JAK1和IL-10RA的相互作用。IL-10RA基因成功构建在pENTER-His中,并在HEK293细胞中成功表达,并成功共转染JAK1和IL-10RA质粒,免疫共沉淀检测两者的相互作用。这为JAK1和IL-10RA相互作用的机制研究奠定基础。
白介素10受体α(IL-10RA);真核表达;免疫共沉淀;蛋白相互作用
人白介素10(Interleukin-10,IL-10)是一种通过跨膜受体介导的重要的免疫细胞调节因子,是重要的抗炎细胞因子之一。普遍认为,IL-10可以抑制炎症反应和各种肿瘤,增强体液免疫应答[1-5]。IL-10是由多种细胞释放的,对T细胞、B细胞和骨髓瘤细胞有不同的影响[6],它可以下调Th1细胞因子的表达,促进B细胞的增值、生存。IL-10受体(IL-10R)由两条链组成,IL-10RA和IL-10RB,IL-10RA可以调节IL-10的免疫抑制信号,从而抑制促炎因子的合成[7,8]。IL-10RA可参与PEDF通路、EGF-β通路、AKT信号通路、JAK-STAT信号通路。IL-10RA的激活可导致非受体型酪氨酸激酶JAK1的激活,引发下游信号转导通路的激活,产生一系列的反应。JAK1是非受体型酪氨酸蛋白激酶的家族成员之一,JAK1的突变或者过度表达可以引发多种疾病,如子宫内膜癌、胃癌等[9,10]。因此,研究JAK1与IL-10RA之间的蛋白相互作用极具意义。
目前对于IL-10RA蛋白的研究主要是对下游的信号通路的激活等,IL-10RA与JAK1的相互作用的研究报道并不多。结合前期的工作,本实验室已成功构建并表达JAK1蛋白,接下来构建IL-10RA的真核表达质粒,并表达IL-10RA蛋白。共转染pENTER-IL-10RA-His和pENTER-JAK1-His重组质粒,用免疫印迹检测两者的表达,免疫共沉淀检测两者的相互作用,旨在为研究两者蛋白相互作用的机制奠定基础。
1 材料与方法
1.1 材料
1.1.1 实验材料 DH5α感受态细胞购自天根生化科技(北京)有限公司;pENTER-JAK1-His质粒、Hela细胞和HEK293细胞由南方医科大学抗体工程研究所保存。
1.1.2 试剂 胎牛血清、1640培养基和DMEM培养基购自Hyclone公司;Total RNA小提试剂盒、和质粒提取试剂盒购自OMEGA公司;SuperScriptTMⅡReverse Transcriptase试剂盒和Lipofectamine® 2000转染试剂购自Invitrogen公司;PrimeStar Max DNA聚合酶和DNA 2000核酸Marker购自TaKaRa公司;限制性内切酶AsiS I和MIu I购自BioLabs公司;一步法克隆试剂盒购自biotool公司。引物合成和质粒测序由华大基因完成。蛋白Marker购自ThermoFisher;PVDF膜购自Bio-RAD公司。鼠源抗GAPDH抗体、鼠源抗His抗体、山羊抗鼠IgGHRP、山羊抗兔IgG-HRP购自Bioworld公司;兔源抗JAK1抗体购自Cell Signaling Technology公司,抗IL-10RA抗体(鼠源)由罗树红教授赠与。Protein G Agrose beads 购自Millipore公司。
1.1.3 仪器设备 PCR扩增仪购自Life Technologies公司;蛋白电泳仪及转移装置购自BioRad公司;核酸电泳仪购自JUNYI。
1.2 方法
1.2.1 引物设计与合成 在GenBank中获取IL-10RA基因序列(NM_001558),根据mRNA编码区,用Primier5.0软件进行引物设计,并在5' 端加入保护碱基和酶切位点,上游引物:5'-GCCGCGAT CGCCATGCTGCCGTGCCTCGTAGTGCTGC-3'(下划线为AsiS I酶切位点),下游引物:5'-GCATGCGCA TCACTCACTTGACTGCAGGCTAGAG-3'(下划线为MIu I酶切位点)。扩增的IL10RA基因片段大小为1 756 bp。
1.2.2 IL-10RA基因的扩增 用含10%胎牛血清的1640培养基培养HeLa细胞,按照Total RNA小提试剂盒的步骤提取人Hela细胞的总RNA,用1%的琼脂糖凝胶电泳鉴定。按照SuperScriptTMⅡ Reverse Transcriptase试剂盒说明书将提取的RNA逆转录成cDNA。PCR体 系:PrimeStar Max 25 μL,cDNA模板5 μL,上游引物2 μL,下游引物2 μL,总体积为50 μL。PCR扩增反应条件:95℃ 10 min,95℃ 10 s,56℃ 5 s,72℃ 2 min,35个循环,72℃ 10 min,4℃保存。用1%的琼脂糖凝胶电泳鉴定扩增产物。
1.2.3 真核表达质粒pENTER-IL-10RA-His的构建 将pENTER-His质粒进行AsiS I和MIu I双酶切3 h,IL-10RA的PCR克隆产物和pENTER-His质粒用胶回收试剂盒回收产物。用一步法克隆试剂盒连接IL-10RA回收产物和回收的pENTER-His产物(IL-10RA和pENTER-His的摩尔比为5:1),16℃,20 h。连接产物转化DH5α感受态细胞,次日挑取单克隆于5 mL 含Kana的LB培养基中,37℃摇床培养10 h,按照质粒提取试剂盒说明书提取质粒。质粒经PCR扩增和双酶切鉴定为阳性克隆后,送至华大基因科技有限公司测序鉴定。
1.2.4 HEK293细胞培养及IL-10RA质粒转染 用含10%胎牛血清的DMEM培养HEK293细胞。转染前1 d将细胞以4×105细胞/孔接种于6孔板培养。次日待细胞生长至60%,按照Lipofectamine® 2000转染试剂说明书转染,质粒(μg):转染试剂(μL)为1∶3的比例将空质粒、重组质粒pENTERIL-10RA-His各2 μg转染至HEK293细胞,转染6 h后HEK293细胞换液,转染48 h后收细胞进行蛋白水平鉴定。
1.2.5 JAK1与IL10RA质粒共转染 转染前1 d将细胞以6×106细胞/皿接种于10 cm培养皿中。用同样的方法将7 μg的pENTER-JAK1-His和3 μg的pENTER-IL-10RA-His重组质粒和30 μL转染试剂转染至HEK293细胞中,7 μg的pENTER-JAK1-His和3 μg的pENTER-IL-10RA-His分别转染作为对照组。48 h后收细胞进行蛋白水平鉴定。
1.2.6 免疫印迹检测蛋白的表达 将收集的蛋白样品加入等体积的SDS-loading buffer(2×),沸水煮15 min使蛋白变性,后14 000 r/min,室温离心15 min。经10% SDS-PAGE进行蛋白电泳。PVDF膜用甲醇浸泡5 min后进行电转移。PVDF膜用5%脱脂奶粉室温封闭2h,分别加入1∶10 000用封闭液稀释的抗His标签的鼠抗和抗GAPDH抗体为一抗,室温孵育1 h。TBST工作液洗涤4次,每次8 min。加入山羊抗鼠IgG-HRP为二抗(1∶10 000稀释),室温孵育1 h。TBST工作液洗涤6次,每次8 min。ECL显影。
1.2.7 免疫共沉淀鉴定IL-10RA和JAK1的相互作用 将共转染表达的JAK1和IL-10RA的细胞用细胞裂解液收集,充分裂解。制备的裂解液中加入2 μg的抗IL-10RA的抗体,以HEK293细胞为对照,4℃结合过夜。40 μL/管 Protein G Agrose beads 5 000 r/min离心1 min,弃上清,用40 μL细胞裂解液重悬,加入到过夜反应的混合液中,4℃结合1 h。5 000 r/min离心1 min弃上清,重复4次,加入40 μL SDS-loading buffer(2×)准备蛋白电泳样品,免疫印迹法检测,这里抗IL-10RA的抗体和抗JAK1的抗体为一抗分别孵育相应位置的条带,相对应的山羊抗鼠IgG-HRP和山羊抗兔IgG-HRP为二抗。
2 结果
2.1 人Hela细胞总RNA鉴定
提取的Hela的总RNA经1%琼脂糖凝胶电泳,结果(图1)显示,有明显的28S和32S两条条带,OD260/280约为1.98,说明此次提取的总RNA可用于逆转录cDNA。

图1 人Hela细胞的总RNA
2.2 IL-10RA扩增产物的鉴定
合成的引物以cDNA为模板经PCR扩增目的片段,产物进行1%琼脂糖凝胶电泳。结果(图2)显示约1 756 bp可见特异性目的条带,与IL-10RA大小一致。
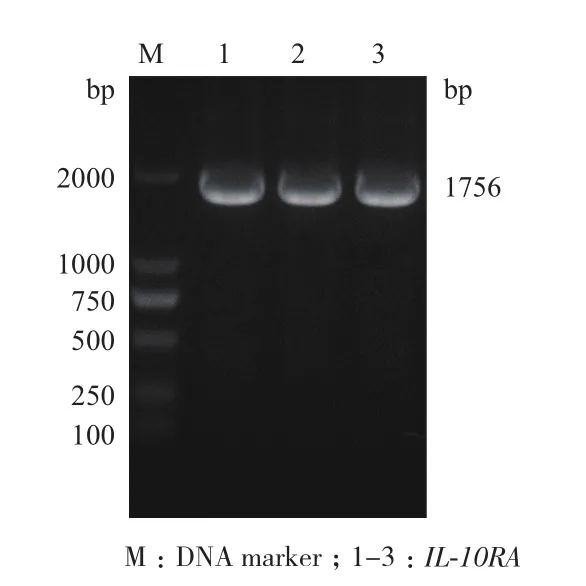
图2 IL-10RA基因PCR产物电泳图
2.3 重组质粒的PCR鉴定
提取质粒并进行重组质粒的PCR鉴定。1%琼脂糖凝胶结果(图3)在约1 756 bp可见特异性目的条带,与IL-10RA大小一致,说明重组质粒含有IL-10RA基因片段。
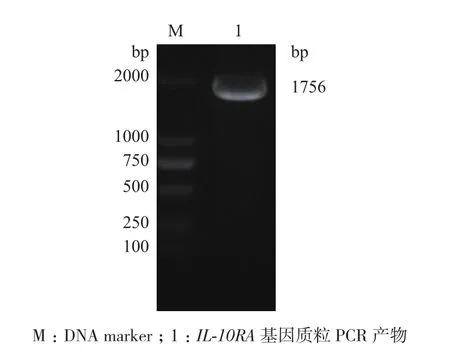
图3 重组质粒pENTER-IL-10RA-His的质粒PCR
2.4 重组质粒的双酶切鉴定
分别用限制性内切酶AsiSⅠ和MIuⅠ对重组质粒进行双酶切鉴定,产物进行1%琼脂糖凝胶电泳(图4),在约1 756 bp和7 472 bp处可见IL-10RA基因和pENTER-His质粒的条带,说明重组质粒构建成功。
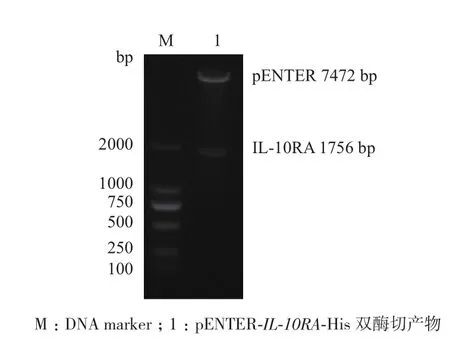
图4 重组质粒pENTER-IL-10RA-His的双酶切鉴定
2.5 免疫印迹法鉴定重组质粒pENTER-IL-10RAHis在HEK293细胞的表达
收 集 转 染48 h的HEK293细 胞,Western blotting鉴定IL-10RA蛋白表达量(图5),以未处理的HEK293细胞作为对照组。实验组HEK293细胞中转染pENTER-IL-10RA-His质粒,Western Blotting鉴定可见约63 kD的IL-10RA蛋白条带。

图5 IL-10RA蛋白表达的western blotting分析结果
2.6 免疫印迹法鉴定共转染质粒pENTER-IL-10RA-His和pENTER-JAK1-His在HEK293的表达
收集共转染重组质粒pENTER-IL-10RA-His和pENTER-JAK1-His的HEK293细胞,检测其蛋白表达量,以未处理的HEK293细胞、单转质粒pENTER-IL-10RA-His、质粒pENTER-JAK1-His为对照组,Western Blotting鉴定。由图6可知,实验组出现两条条带,位置在约63 kD和133 kD处;未处理的HEK293细胞未出现特异性条带;单转染的质粒pENTER-IL-10RA-His在约63 kD有条带出现;单转染的质粒pENTER-JAK1-His在133 kD处出现条带,这与共转染的结果相符,说明共转染质粒成功表达IL-10RA和JAK1蛋白。

图6 JAK1和IL-10RA共转染结果分析
2.7 免疫共沉淀检测IL-10RA和JAK1蛋白的相互作用
共转染JAK1和IL-10RA质粒的细胞裂解液用抗IL-10RA的抗体捕获裂解液中的IL-10RA,同时捕获的还有可以与之相互作用的JAK1蛋白。Protein G Agrose beads可特异性结合抗IL-10RA的抗体。加入Protein G Agrose beads后, 抗IL-10RA的 抗 体和IL-10RA及与之相互作用的蛋白JAK1均结合在Protein G Agrose beads上,通过低速离心,使得与裂解液中的其他蛋白分离,Western Blotting鉴定(图7)。HEK293细胞对照组只出现抗体的重链和轻链,而实验组除了抗体的重链和轻链,还有目的条带JAK1和IL-10RA的条带,这说明两个蛋白在HEK293细胞中存在相互作用,并且可以用免疫共沉淀检测。
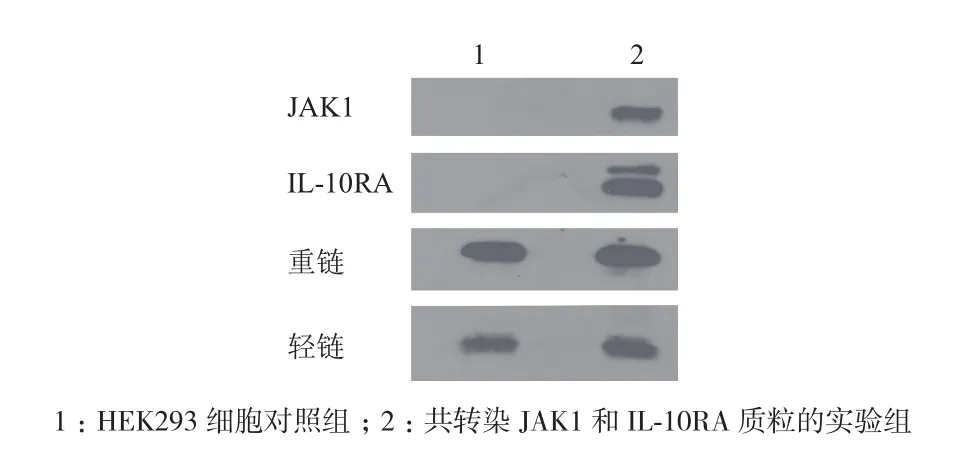
图7 检测JAK1蛋白和IL-10RA蛋白相互左右的免疫共沉淀分析
3 讨论
自90年代开始,人类开始实施基因组计划,至2003年经过世界各国科学家的努力,提前完成了人全基因组测定。但是,大多数疾病并非仅仅因为基因改变造成,随后又提出后基因组计划,蛋白质组的研究占据及其重要的部分,但是对于蛋白质功能及其相互作用方面的研究仍相对缺乏。目前研究蛋白质相互作用技术方法很多,如噬菌体展示技术、串联亲和纯化技术、pull down,免疫共沉淀等[11-17]。研究发现,IL-10可以调节相关细胞的神经炎症反应,而且也存在抗凋亡作用。IL-10通过与神经元上的IL-10RA相互作用,导致JAK1-STAT3通路的激活,抑制神经元凋亡和神经炎症[18]。Wagner等[19]发现JAK1激活程度不同可引起下游不同的信号转导通路,JAK1的抑制可某种程度上减轻自身免疫病及炎症,这提示未来JAK1的抑制剂可能可以应用于自身免疫性疾病的治疗。但是,目前对于IL-10RA蛋白的研究主要致力对下游信号通路的激活等,IL-10RA与JAK1的相互作用的研究报道并不多。因此,本研究尝试用免疫共沉淀的方法检测IL-10RA与JAK1的蛋白相互作用。而IL-10RA参与多条信号通路,因此与其存在相互作用的蛋白必然很多,现今用经典的免疫共沉淀的方法对这些相互作用蛋白尚并不能定量,而且检测过程繁琐而复杂。因此,尝试用免疫分析的方法如ELISA、时间分辨免疫分析、化学发光免疫分析等,以期对已知存在相互作用的蛋白进行简便快速的检测,同时也可为其他研究者寻找未报道的存在相互作用的蛋白提供便利,这将是未来的一个研究方向。
4 结论
本实验从Hela细胞中提取人总RNA,并通过逆转录PCR获得IL-10RA基因全长,以pENTERHis为载体构建重组真核表达质粒转染至HEK293细胞表达蛋白,Western blot可检测到与人IL-10RA大小一致的蛋白条带。共转染及免疫共沉淀均可检测到IL-10RA和JAK1。真核表达蛋白特异性较强,具有生物学活性,其结构更接近天然蛋白。
[1]Mocellin S, Marincola FM, Young HA. Interleukin-10 and the immune response against cancer:a counterpoint[J]. Journal of Leukocyte Biology, 2005, 78(5):1043-1051.
[2]von Lanzenauer SH, Wolk K, Kunz S, et al. Interleukin-10 receptor-1 expression in monocyte-derived antigen-presenting cell populations:dendritic cells partially escape from IL-10’s inhibitory mechanisms[J]. Genes & Immunity, 2015, 16(1):8-14.
[3]Trifunović J, Miller L, Debeljak Z, et al. Pathologic patterns of interleukin 10 expression--a review[J]. Biochemia Medica, 2015, 25(1):36-48.
[4]Kim R, Emi M, Tanabe K, et al. Tumor-driven evolution of immunosuppressive networks during malignant progression[J]. Cancer Research, 2006, 66(11):5527-5536.
[5]Darnell JJ, Kerr IM, Stark GR. Jak-STAT pathways and transcriptional activation in response to IFNs and other extracellular signaling proteins[J]. Science, 1994, 264(5164):1415-1421.
[6]Beser OF, Conde CD, Serwas NK, et al. Clinical features of interleukin 10 receptor gene mutations in children with very earlyonset inflammatory bowel disease[J]. Journal of Pediatric Gastroenterology & Nutrition, 2014, 60(3):332-338.
[7]Pestka S, Krause CD, Sarkar D, et al. Interleukin-10 and related cytokines and receptors[J]. Annual Review of Immunology, 2004, 22:929-979.
[8]Commins S, Steinke JW, Borish L. The extended IL-10 superfamily:IL-10, IL-19, IL-20, IL-22, IL-24, IL-26, IL-28, and IL-29[J]. Journal of Allergy & Clinical Immunology, 2008, 121(5):1108-1111.
[9]Stelloo E, Versluis MA, Nijman HW, et al. Microsatellite instability derived JAK1 frameshift mutations are associated with tumor immune evasion in endometrioid endometrial cancer[J]. Oncotarget, 2016,7(26):39885-39893.
[10]Song B, Zhan H, Bian Q, et al. Piperlongumine inhibits gastric cancer cells via suppression of the JAK1, 2/STAT3 signaling pathway[J]. Molecular Medicine Reports, 2016, 13(5):4475-4480.
[11]Li Q, Gu S, Li N, et al. Protein interaction network construction and biological pathway analysis related to atherosclerosis][J]. Sheng Wu Yi Xue Gong Cheng Xue Za Zhi, 2015, 32(6):1255-1260.
[12]Verhelst J, De Vlieger D, Saelens X. Co-immunoprecipitation of the Mouse Mx1 Protein with the Influenza A Virus Nucleoprotein[J]. J Vis Exp, 2015(98):e52871.
[13]Duan JW, Lu Z, Jiang B, et al. Discovery of pyrrolo[1, 2-b]pyridazine-3-carboxamides as Janus kinase(JAK)inhibitors[J]. Bioorganic & Medicinal Chemistry Letters, 2014, 24(24):5721-5726.
[14]Einarson MB, Pugacheva EN, Orlinick JR. GST Pull-down[J]. CSH Protoc, 2007(2007):t4757.
[15]Valero ML, Sendra R, Pamblanco M. Tandem affinity purification of histones, coupled to mass spectrometry, identifies associated proteins and new sites of post-translational modification in Saccharomyces cerevisiae[J]. J Proteomics, 2016, 136:183-192.
[16]Saito Y, Nakagawa T, Kakihana A, et al. Yeast Two-Hybrid and One-Hybrid screenings identify regulators of hsp70 gene expression[J]. J Cell Biochem, 2016, 117(9):2109-2117.
[17]Yu Y, Li Y, Zhang Y. Yeast Two-Hybrid screening for proteins that interact with the extracellular domain of amyloid precursor protein[J]. Neurosci Bull, 2016, 32(2):171-176.
[18]Zhu Y, Chen X, Liu Z, et al. Interleukin-10 protection against lipopolysaccharide-induced neuro-inflammation and neurotoxicity in ventral mesencephalic cultures[J]. International Journal of Molecular Sciences, 2016, 17(1).
[19]Wagner AH, Conzelmann M, Fitzer F, et al. JAK1/STAT3 activation directly inhibits IL-12 production in dendritic cells by preventing CDK9/P-TEFb recruitment to the p35 promoter[J]. Biochem Pharmacol, 2015, 96(1):52-64.
(责任编辑 李楠)
Eukaryotic Expression of Human Interleukin 10 Receptor α and Detection of Interactions in Protein JAK1s
GUO Xin-xin WANG Gang DENG Qiao-ting WU Han-tao LI Kun WU Ying-song LIU Tian-cai
(Institute of Antibody Engineering,School of Laboratory Medicine and Biotechnology,Southern Medical University,Guangzhou 510515)
This work is to construct a eukaryotic express vector pENTER-IL-10RA-His that expresses human interleukin 10 receptor α(IL-10RA)in HEK293 cells,and then to detect the intracellular interaction between JAK1 and IL-10RA by co-immunoprecipitation. Human total RNA was extracted from Hela cells,then IL-10RA gene was amplified by RT-PCR and inserted into eukaryotic vector pENTER-His. After validation by PCR,enzymatic digestion,and sequencing,the HEK293 cells were transfected with the recombined plasmid pENTER-IL-10RA-His. The expression level of IL-10RA at 48 h was determined by Western blotting. The results revealed that plasmid was cloned correctly,and the target protein of 63 kD was observed. When HEK293 cells were simultaneously transfected with the plasmid of both JAK1 and IL-10RA,the target protein band in 133 kD and 63 kD were identified. Co-immunoprecipitation validated the interaction between JAK1 and IL-10RA. In conclusion,IL-10RA gene was successfully constructed in recombined plasmid pENTER-His and expressed effectively in HEK293 cells. The interaction between JAK1 and IL-10RA was validated co-immunoprecipitation,which lays a foundation for further understanding the mechanism of interaction between JAK1 and IL-10RA.
interleukin 10 receptor α(IL-10RA),eukaryotic expression,co-immunoprecipitation,protein - protein interaction
10.13560/j.cnki.biotech.bull.1985.2016-0985
2016-10-28
国家高技术研究发展计划(“863”计划)项目(2014AA020904),国家自然科学基金项目(21575058、81271931)
郭欣欣,女,硕士研究生,研究方向:免疫学,蛋白质相互作用;E-mail:347315578@qq.com
刘天才,男,研究员,研究方向:免疫学;E-mail:liutc@smu.edu.com
