紫萼玉簪种子和幼苗对酸雨与镉复合污染的生理生态响应
2017-05-19罗良旭高素萍王成聪雷霆闻金燕罗雁
罗良旭,高素萍,2*,王成聪,雷霆,2,闻金燕,罗雁
(1.四川农业大学风景园林学院,成都611130;2.四川农业大学园林研究所,成都611130)
紫萼玉簪种子和幼苗对酸雨与镉复合污染的生理生态响应
罗良旭1,高素萍1,2*,王成聪1,雷霆1,2,闻金燕1,罗雁1
(1.四川农业大学风景园林学院,成都611130;2.四川农业大学园林研究所,成都611130)
为了解紫萼玉簪生长初期对酸雨与镉(Cd)复合污染的生理生态响应,探究该物种在复合污染下的自我更新能力,分别以紫萼玉簪种子和幼苗为供试材料,采用2因子5水平完全随机试验,研究不同梯度(pH分别为6.5、5.5、4.5、3.5、2.5)模拟酸雨与不同质量浓度重金属Cd2+(0、10、30、50、100 mg/L)复合污染胁迫下,该植物种子和幼苗的生理生态指标变化。结果表明:酸雨和Cd复合污染比单一污染对紫萼玉簪种子和幼苗造成的伤害更大,两者之间交互作用显著,表现为协同作用,且这一作用在高强度复合污染下表现得尤为突出;紫萼玉簪种子和幼苗对低强度酸雨和低质量浓度Cd胁迫有一定的耐受性,其中,低质量浓度Cd2+(10 mg/L)处理对种子萌发表现出一定的促进作用。幼苗对Cd胁迫的耐受能力弱于成苗,在本试验条件下,耐受的临界质量浓度值为30 mg/L。过氧化氢酶活性对Cd污染胁迫的敏感性高于超氧化物歧化酶,尤其在与酸雨复合作用下敏感性增加,表现在重金属Cd2+质量浓度为30 mg/L时就到达峰值,这也进一步导致其在高质量浓度Cd2+污染时对活性氧的清除能力下降,使活性氧的产生速率超过抗氧化酶系统的清除能力,从而使幼苗完全丧失了自我保护的能力。综合各指标结果认为,紫萼玉簪种子萌发和幼苗生长分别在低强度酸雨(pH≥5.5)与低质量浓度Cd2+(≤10和30 mg/L)污染胁迫下能顺利进行自我更新,而高强度复合污染对种子萌发和幼苗生长极为不利。该研究结果对复合污染地区的土壤环境修复工作具有一定的参考价值。
镉;酸雨;复合污染;生态胁迫;抗性生理;紫萼玉簪
SummaryThe combined pollution of acid rain and cadmium(Cd)has been an important component of soil interacted with atmospheric pollution in China.The combined pollution posed harmful impact on human beings,and threatened the habitats of other creature,such as the urban garden plants.Therefore,it is important to investigate the effect of complicated pollution on plant growth,extension of plant population and ecosystem balance.The seed germination is the origin of the plant’s life cycle, which is the key link affecting extension of plant population.The stress resistance was bad in the seeding stage,especially poorer resistance to single and combined pollution compared to seedling stage.Therefore,the seeding period became the key toaccomplish extension of plant population under habitat destruction.
Previous studies onHosta ventricosafocused on single pollution of Cd or acid rain;recentlyH.ventricosawas found to have tolerance against combined pollution of Cd and acid rain.Therefore,it is potential for phytoremediation of Cd polluted soil in acid rain area.
In this study,H.ventricosawas selected as test material,to investigate the physico-ecological responses under combined pollution of simulated acid rain and Cd.The tolerance ofH.ventricosawas examined under both seed germination and seedling periods,to provide science evidence for selection of urban plants with strong stress resistance under combined pollution.A completely random scheme of two factors(simulated acid rain and Cd)and five levels was used.The simulated acid rain was set at five pH levels(6.5,5.5,4.5,3.5 and 2.5),and the Cd treatment fluids were set at five different Cd2+concentrations(0,10,30, 50 and 100 mg/L).Both normal and abnormal germination rates were measured in each group.Meanwhile,after the seedling ofH.ventricosagrew up to 10 to 15 cm height,the contents of chlorophyll,malondialdehyde(MDA)and H2O2,the activities of peroxidase(POD),superoxide dismutase(SOD)and catalase(CAT)were measured in each group to investigate the physicoecological responses.
Results showed that the combined pollution of acid rain and Cd had significant higher impact on seed germination and seedling growth ofH.ventricosathan single pollution.The interaction of acid rain and Cd posed obviously synergistic effect, especially in high strength pollution(Cd2+concentration≥30 mg/L).Seed germination and seedling growth ofH.ventricosashowed tolerance under stress of low concentration Cd2+(10 mg/L)and low intensity acid rain,and the seed germination was even promoted by low concentration Cd2+of 10 mg/L.The tolerance of seedling under Cd stress was weaker than mature seedling,and the critical tolerance value was believed to be 30 mg/L.The CAT activity was more sensitive than POD and SOD activites under the Cd stress,especially under the combined pollution of Cd and acid rain,and the peak value of the CAT activity was observed under 30 mg/L Cd2+.Therefore,the ability of CAT to eliminate reactive oxygen species(ROS)decreased under high concentration of Cd stress(≥30 mg/L).The production of H2O2was faster than scavenging by antioxidant enzyme systems,so the seedling would completely lose the self-defensive ability.
In conclusion,the seed germination and seedling growth can complete the self-update under the combined pollution of low intensity acid rain(pH≥5.5)and low concentration Cd2+(≤10 and 30 mg/L),respectively,but high intensity of the combined pollution will significantly affect seed germination and seedling growth,which can provide reference for phytoremediation of combined polluted soil environment.
酸雨和重金属等污染使城市绿化植物的生境问题正变得日趋严重[1-2]。在自然条件下,多种污染物进入土壤后相互作用并最终形成复合污染[3]。复合污染不同于单一污染,往往表现出十分复杂的复合污染特点,各成分之间可能会产生加和、协同或拮抗等作用。目前,我国酸雨和重金属镉(Cd)的复合污染已成为土壤与大气交互污染的重要组成部分,并具有普遍性,不仅给人类带来危害,也给其他生物赖以生存的环境形成很大的威胁。酸雨进入到土壤会影响土壤中Cd的化学性质及其活性,激活并增加Cd对绿色植物的毒性,二者相互作用,往往会导致城市园林植物的生存条件进一步恶化[4]。这种复杂的生存条件对植物的影响,以及它们能否顺利完成自我更新、保持种群的延续和系统的稳定是一个值得重视的课题。种子萌发是植物生长周期的起点,是否能顺利地正常萌发直接决定植物种群能否进行自我更新;而植物幼苗处于整个生长周期的初期,对外界环境的抗逆性较差,对复合性、突发性等污染的抵御能力较成苗更弱,因此,幼苗时期也成为植物在生境遭到破坏时是否能顺利完成自我更新的关键点。紫萼玉簪(Hosta ventricosa)是园林中应用较广的植物。目前,国内外对该植物的相关研究主要集中在铅(Pb)胁迫方面[5],而对其在模拟酸雨及Cd复合或单一胁迫方面的相关研究较少。王成聪等[6]对该植物的前期研究发现,无论是酸雨和Cd的单一污染还是复合污染,紫萼玉簪成苗都表现出较强的耐受性,因此,该植物成苗可作为酸雨区土壤Cd污染背景下修复植物的选择。但该植物种群在自我更新过程中非常关键的种子萌发和幼苗阶段是否同样具有耐受性尚未见报道。为此,本试验采用人工模拟酸雨与Cd复合污染对紫萼玉簪种子萌发和幼苗生长2个阶段进行处理,研究该植物在复合污染下的生理生态响应,以期为城市绿化选择既能抗酸雨和重金属复合污染又能在污染环境下完成自我更新的植被提供科学依据。
1 材料与方法
1.1 供试材料
紫萼玉簪(Hosta ventricosa)种子采自四川农业大学成都校区苗圃基地,经除杂、去皮和筛选后,室外风干,储藏待用。幼苗是种子播种后生长的当年生苗,苗木生长健康,长势一致,高约10~15 cm。
1.2 模拟酸雨溶液配制
根据成都市温江区气象局提供的气象观测资料,参考成都市近年来降水的主要离子组成及浓度,参照王成聪等[6]的方法配制本试验研究所需溶液(表1)。

表1 模拟酸雨溶液中各主要离子组成及其浓度Table 1 Main components and ion concentrations of the simulated acid rainμmol/L
1.3 外源Cd溶液配制
根据国家土壤环境质量标准以及四川盆地Cd污染情况,设置5个Cd质量浓度梯度,分别为0、10、30、50、100 mg/L(以纯Cd2+计)。具体配制参照王成聪等[6]的方法。
1.4 试验方法
1.4.1 种子萌发试验设计
取均匀一致的紫萼玉簪种子,去黑皮,用0.5%KMnO4溶液浸泡0.5 h,消毒备用。采用2因子5水平完全随机方案(表2),设置模拟酸雨和Cd处理2个因子,分别为5个水平,共25个处理组合,每个处理重复3次。模拟酸雨处理液pH值分别为6.5、5.5、4.5、3.5、2.5,表2中分别对应代号为b1、b2、b3、b4、b5;Cd处理质量浓度梯度为0、10、30、50、100 mg/L(以纯Cd2+计),表2中分别对应代号为a1、a2、a3、a4、a5。用处理液分别浸种24 h,各取30粒种子均匀放入培养皿中,定期更换相应浓度的复合处理液(每皿等量),以保持种子的湿润状态。光照培养箱温度设置为25℃,光照条件为4 000 lx。紫萼玉簪种子10 d左右即萌发结束,可选择连续3 d发芽率不再变化时进行相关指标测定。
发芽率=(正常发芽种子数/供试种子数)× 100%;
异状发芽率=(发芽异常种子数/供试种子数)× 100%。

表2 Cd和模拟酸雨复合处理组合设计Table 2 Design of combination treatment of Cd and simulated acid rain
1.4.2 幼苗生理试验设计
同样采用2因子5水平完全随机试验设计(表2)。本阶段在种子萌发试验滤纸法的基础上加以改进,克服滤纸法的缺点与不足,以增进试验的可控性。考虑到本阶段试验周期相对较长,将种子萌发着床的基质改为保水性更好的脱脂棉,将播种器皿更换成容量为1 L的烧杯(每个烧杯内均匀放置10粒种子),更换处理液的周期延长至每周1次,从而可大大降低由于频繁更换处理液导致的Cd2+浓度叠加效应及种子污染情况的发生,以保证试验的准确性。
待紫萼玉簪幼苗长高至10~15 cm时,各处理间幼苗的形态特征差异明显,生物量也已足够进行试验指标的测定,同时,模拟酸雨与Cd复合污染的伤害在形态学上也表现出十分明显的差异,因而,选择在此时进行幼苗相关生理指标的测定:叶绿素含量采用分光光度法,用80%丙酮浸提[7];丙二醛(malondialdehyde,MDA)含量参照李合生[7]的方法测定;H2O2含量参照PATTERSON等[8]的方法测定;过氧化物酶(peroxidase,POD)活性测定采用愈创木酚法[7,9];超氧化物歧化酶(superoxide dismutase,SOD)活性测定采用氮蓝四唑光还原法[10];过氧化氢酶(catalase,CAT)活性测定采用紫外分光光度法[10]。
1.4.3 数据处理
用SPSS 20.0软件进行数据分析,用Excel 2016制作图表。
2 结果与分析
2.1 在复合污染环境中紫萼玉簪种子的萌发情况
2.1.1 对种子发芽率的影响
种子发芽率可以定量体现种子的萌发能力。由表3可以看出,模拟酸雨和Cd污染对紫萼玉簪种子萌发产生了很大的影响。
在单一的酸雨或Cd处理下,随着pH值下降或Cd质量浓度增加,紫萼玉簪种子萌发率整体呈下降趋势。当Cd2+在10 mg/L低质量浓度条件下时对种子萌发表现出一定的促进作用:这表明紫萼玉簪种子具有一定的耐Cd污染能力。低强度酸雨(pH=5.5)对种子萌发率的影响与对照相比差异不显著:说明紫萼玉簪种子对酸雨具有一定的耐受力,在低强度酸雨(pH≥5.5)下能顺利完成自我更新。
在模拟酸雨和Cd复合污染条件下,随着复合胁迫强度的提高,种子萌发率整体呈现出下降的态势,这一作用在高质量浓度重金属Cd2+与高强度模拟酸雨的复合处理下表现得尤为突出。在复合强度较低(Cd2+质量浓度≤10 mg/L,pH≥5.5)时,萌发率与对照相比差异不显著:表明种子对低复合胁迫强度具有一定的耐受性。在中等强度酸雨(pH=4.5)、低质量浓度Cd2+(10 mg/L)处理下,复合与单一处理相比,种子萌发率降低,且差异有统计学意义(P<0.05):表明两者间存在交互作用,且表现为协同作用。在Cd2+质量浓度≥30 mg/L胁迫下,与对照相比,随着酸雨强度增大,种子萌发率显著降低直至为0:表明在此质量浓度Cd2+胁迫下,两者的复合作用仍表现为协同作用,加剧了对紫萼玉簪种子的伤害。
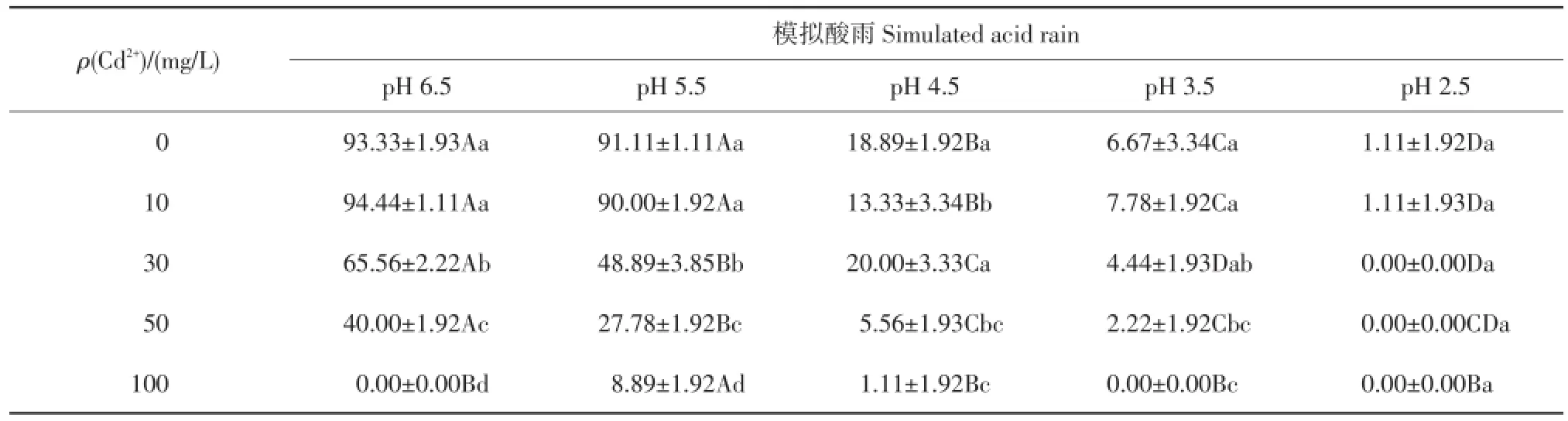
表3 复合污染对紫萼玉簪种子发芽率的影响Table 3 Effects of combined pollution on the seed germination ofHosta ventricosa%
2.1.2 对种子异状发芽率的影响
种子在受到外界环境干扰时,常常会引起发芽异常。通过测定种子的异状发芽率,可以了解在逆境下种子萌发受到的伤害强度。如表4所示,pH≥ 5.5时,种子异状发芽率为0,表明紫萼玉簪种子具有一定的耐酸雨能力;但随着pH值继续下降,种子异状发芽率明显增大,与对照(pH=6.5)相比差异显著,说明高强度酸雨对种子的伤害程度较大。在单一Cd处理下,种子异状发芽率在Cd2+质量浓度为10 mg/L时为0,表明紫萼玉簪种子对低质量浓度重金属具有一定的耐受能力;随着Cd2+胁迫质量浓度的上升,种子异状发芽率明显增大,表明高质量浓度Cd2+可以毒害紫萼玉簪种子,使其发育成为不健全的芽。在高强度的复合胁迫下,紫萼玉簪种子异状发芽率较低,其表面效应并不代表胁迫伤害程度的大小,这与复合处理下紫萼玉簪种子萌发数较低有关。
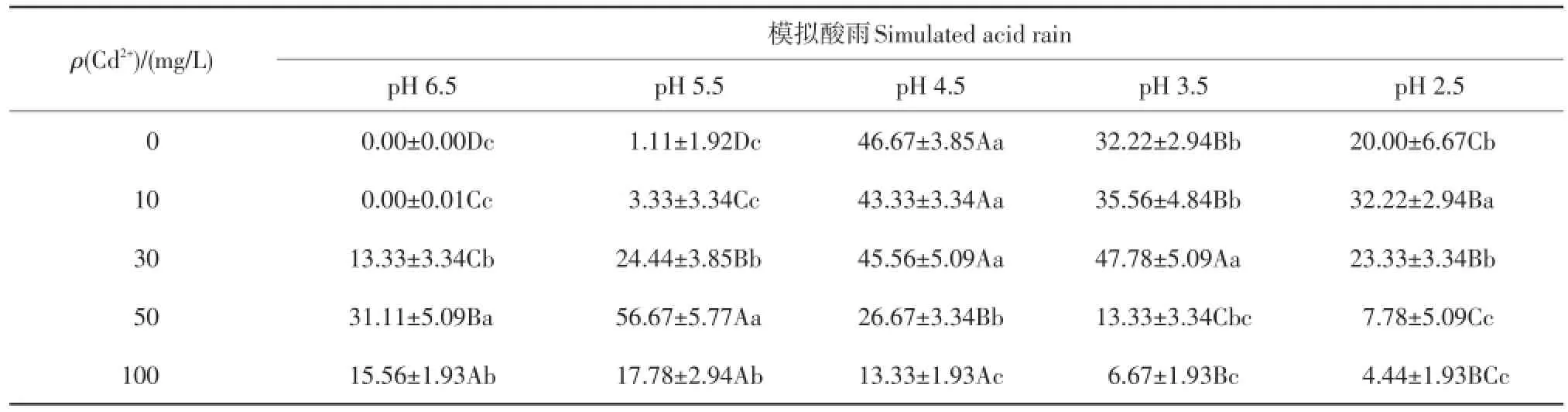
表4 复合污染对紫萼玉簪种子异状发芽率的影响Table 4 Effects of combined pollution on the abnormal seed germination rate ofHosta ventricosa%
2.2 紫萼玉簪在复合污染环境中幼苗的生理变化
2.2.1 对幼苗叶绿素含量的影响
由表5可知:在单一模拟酸雨处理下,随着pH值的下降,紫萼玉簪幼苗叶片中的叶绿素含量变化整体呈十分明显的下降趋势,与对照差异显著。在单一Cd处理下,与对照(Cd2+质量浓度为0 mg/L)相比,当Cd2+质量浓度≥30 mg/L时,紫萼玉簪叶片中的叶绿素含量显著下降。在复合处理下,Cd2+质量浓度为10 mg/L时,各酸雨胁迫与对照相比差异不显著;当Cd2+质量浓度≥30 mg/L时,与对照相比,随着酸雨强度的增大,叶绿素含量显著降低,至高酸度(pH=2.5)和高Cd2+质量浓度(50~100 mg/L)时含量为0,表明两者间存在着交互作用,且表现为协同作用。
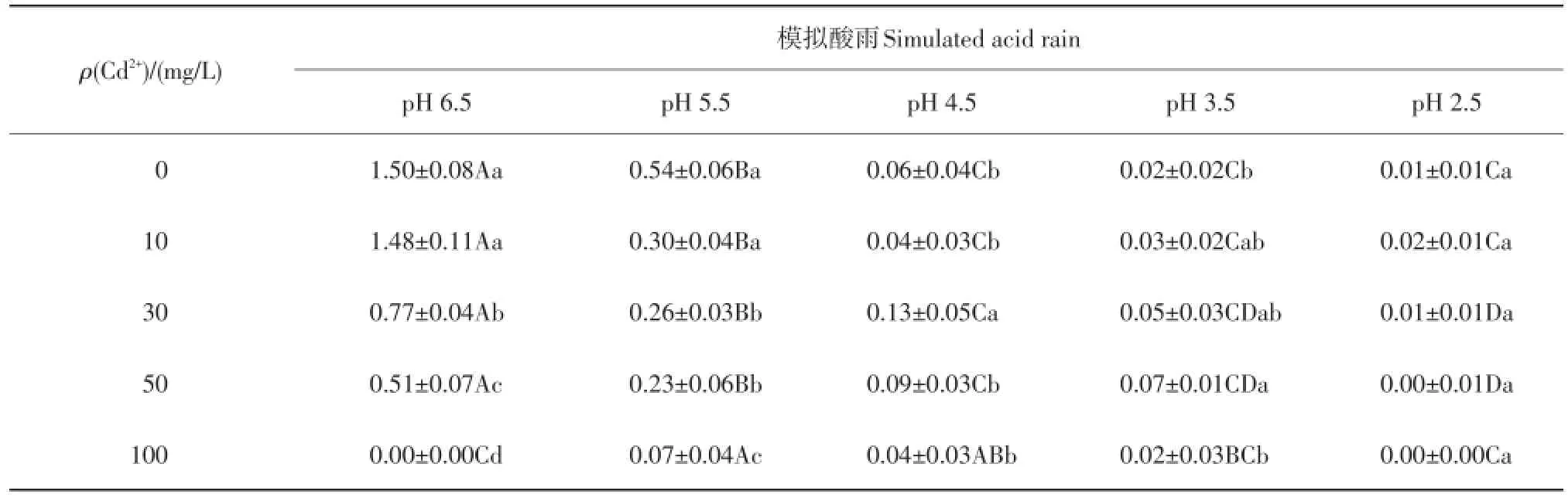
表5 复合污染对紫萼玉簪叶片叶绿素含量的影响Table 5 Effects of combined pollution on the leaf chlorophyll content ofHosta ventricosamg/g
2.2.2 对幼苗叶片质膜过氧化的影响

图1 模拟酸雨与Cd复合污染对紫萼玉簪幼苗叶片MDA含量的影响Fig.1 Effects of combined pollution of simulated acid rain and Cd on the MDA content in leaves ofHosta ventricosaseedlings
由图1、2可知,随着酸雨pH值的下降,紫萼玉簪幼苗叶片中的MDA含量和H2O2产生速率整体呈上升趋势,两者与pH值呈负相关关系。与对照相比,在单一Cd处理下,当Cd2+质量浓度≤10 mg/L时,MDA含量和H2O2产生速率的变化趋势均不明显;而当Cd2+质量浓度≥30 mg/L时,两者均随Cd胁迫强度增强而明显增大,与对照差异显著。在复合处理下,酸雨和Cd之间交互作用显著,且表现为协同作用。结合叶绿素含量变化,可初步认为幼苗顺利完成自我更新的临界值为酸雨pH=5.5和ρ(Cd2+)=30 mg/L。
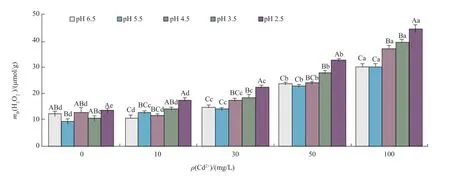
图2 模拟酸雨与Cd复合污染对紫萼玉簪幼苗叶片H2O2含量的影响Fig.2 Effects of combined pollution of simulated acid rain and Cd on the H2O2content in leaves ofHosta ventricosaseedlings
2.2.3 对幼苗抗氧化酶活性的影响
由图3A可知,随着酸雨和Cd的单一处理强度的增加,SOD活性持续增强。单一酸雨处理时,随着pH值的下降,叶片中的SOD活性不断增强,当pH≤3.5时,SOD活性才开始明显升高,与对照(pH=6.5)相比,差异达到显著水平。在单一Cd处理下,随着Cd胁迫强度的持续增大,SOD活性呈现出持续增大的趋势,在Cd2+质量浓度为100 mg/L时SOD活性达到最大值。复合污染与单一处理趋势相似,随着复合强度的增加,SOD活性持续增加,在中低强度酸雨(pH≥4.5)单一处理时,与对照(pH=6.5)相比差异不显著,而与Cd(≤30 mg/ L)复合后,SOD活性降低,与对照相比差异显著,这说明中低强度酸雨与Cd污染[pH≥4.5,ρ(Cd2+)≤30 mg/L]对SOD活性的交互作用表现为拮抗作用;而高强度酸雨与高质量浓度Cd复合[pH≤3.5,ρ(Cd2+)≥50 mg/L]的交互作用则更多地表现为协同和加和作用。
由图3B可知:单一酸雨处理时,POD活性变化趋势与SOD相似;单一Cd处理时,随着Cd胁迫强度增加,POD活性提高,在Cd2+质量浓度为50 mg/L处出现峰值,之后逐渐下降;随着复合污染强度的增加,POD活性先增加后减少,复合胁迫对POD活性的交互作用表现为协同和加和作用。
由图3C可知:单一酸雨处理时,CAT活性变化趋势与SOD、POD一致,不同的是,当pH=4.5时,CAT活性与对照(pH=6.5)相比差异显著,这说明CAT与SOD、POD相比,对酸雨胁迫的敏感性更高;单一Cd处理时,CAT活性变化趋势与POD一致,不同的是,CAT在Cd2+质量浓度为30 mg/L处出现峰值。这进一步说明,CAT无论对酸雨还是重金属Cd污染胁迫都更为敏感。在复合污染条件下,复合胁迫对CAT活性的交互作用也表现为协同作用和加和作用,与POD相似。
值得注意的是,在酸雨pH=2.5时,各处理组合的SOD活性随着Cd胁迫强度增大而显著增大,远远高于其他处理组合的SOD活性,表现出明显的爆发式增长现象,这对缓解紫萼玉簪受到的胁迫起着积极作用,但此时POD和CAT活性呈下降趋势,说明抗氧化酶系统的平衡遭到巨大破坏,细胞受伤害严重。这一结果与MDA和H2O2含量变化相印证,进一步说明高强度酸雨与重金属Cd的复合污染胁迫虽然刺激了保护酶的活性,但因此时清除活性氧的酶功能受限,不利于发挥抗氧化酶系统的功能,故导致植物幼苗受到严重伤害。
3 讨论
3.1 复合污染对紫萼玉簪种子萌发的影响
种子萌发是植物自我更新的起始,这一时期种子非常脆弱,遭受酸雨胁迫时,其细胞代谢途径被扰乱,内部结构受到破坏,种子胚和幼芽正常生长也受到干扰[11]。通过对种子在逆境胁迫下的萌发情况研究,可以较为直观地发现逆境胁迫对植物的损害情况[12]。本试验表明,低强度模拟酸雨(pH≥5.5)对紫萼玉簪种子发芽率、异状发芽率的影响不大,而中等强度的模拟酸雨(pH≤4.5)则严重抑制了紫萼玉簪种子萌发,导致种子发芽率降低和异状发芽率升高,这一结果进一步证明了酸雨对种子萌发的危害,同时也说明植物种子对酸雨胁迫具有一定的耐受性,但这种忍受有一定的限度。此外,种子萌发也容易遭受土壤重金属的伤害[13],无毒的重金属Cd变为游离态Cd2+,与其他物质化合后再被植物吸收,就会发挥出极大的毒性,并可富集于植物体内,带来持续伤害。PATRA等[14]的研究结果表明,低浓度Cd会对植物表现出正面的“刺激作用”。这一结论在本试验中也得到证实,可能是因为幼苗生长需要少量的Cd。曹德菊等[15]研究发现,低浓度Cd可能促进了植物细胞分裂,从而对其生长起到了一定的促进作用。而高浓度Cd却能严重抑制紫萼玉簪种子萌发,表明高浓度的重金属会对种子产生毒害作用,还可能会促使种子进入休眠状态,与崔大练等[16]的研究成果一致。当酸雨与Cd形成复合污染后,本试验发现,低质量浓度Cd2+(10 mg/L)与中低强度酸雨(pH≥4.5),以及高质量浓度Cd2+(≥30 mg/L)与所有模拟酸雨强度间存在明显的交互作用。通过对各复合处理分析发现:复合污染胁迫对种子萌发的影响并非是单一污染胁迫结果的简单加和,更多地表现为协同作用;随着胁迫强度的增加,这种作用尤为明显,加剧了对种子的毒害作用。这可能是因为酸雨改变了培养溶液的pH值,使得溶液中的重金属离子有效性增强,从而使植物对重金属离子的吸收作用增强[17]。

图3 模拟酸雨与Cd复合污染对紫萼玉簪幼苗叶片抗氧化酶活性的影响Fig.3 Effects of combined pollution of simulated acid rain and Cd on the activities of antioxidant enzymes in leaves ofHosta ventricosaseedlings
3.2 复合污染对紫萼玉簪幼苗生理指标的影响
幼苗期是植物对外界环境因子最敏感的生长阶段之一[18],也是植物自我更新的关键点。为此,本研究通过测量在胁迫下幼苗生长的各项生理指标,可灵敏地反映出复合胁迫对植株幼苗本身的伤害,提前预知幼苗的自我更新能力。
叶绿素是参与光合作用的重要色素[19],其含量往往决定着植物光合作用有机质的生产量和累积量[20]。当植物处于酸雨逆境胁迫下时,地上部分便会直接遭受到酸雨的毒害作用,因为酸雨中存在的大量的H+直接接触到叶片表面的角质层,与叶片内部的阳离子发生交换反应,最终严重破坏叶片组织与细胞结构[21]。而幼嫩的紫萼玉簪幼苗叶片由于代谢旺盛、角质层较薄、保卫组织发育不完全等,相较于成熟叶片会遭受到更大的伤害[22]。低强度酸雨(pH=5.5)即可对紫萼玉簪幼苗叶片叶绿素含量造成严重伤害,使其含量大大降低,影响幼苗生长,从而影响其自我更新能力。Cd能破坏植物叶绿体的结构,降低光合酶活性以及叶绿素的合成速率,导致叶绿素总量下降,光合作用强度下降[23]。低质量浓度的Cd2+(10 mg/L)对紫萼玉簪幼苗叶绿素含量未产生显著的抑制作用,当Cd2+质量浓度≥30 mg/L,叶绿素含量才开始显著降低。由此可见,紫萼玉簪幼苗对酸雨的生理响应敏感,对Cd的耐受力更强,在本试验条件下,其对Cd耐受的临界质量浓度值为30 mg/L。
SOD、POD和CAT是植物体内抗氧化酶系统的重要组成酶类,三者协同使得植物体内的活性氧(reactive oxygen species,ROS)含量维持在较低水平,从而防止逆境胁迫对植物造成的ROS伤害[24-25]。ROS的产生与抗氧化酶的清除维持着动态平衡。SOD是植物防御机制的第一道防线[26],能将转化为H2O2,防止其对细胞造成伤害;POD和CAT紧随其后,将SOD生成的H2O2转化为无害的水[27],使其维持在一个较低的、对细胞无害的浓度。酸雨和Cd胁迫可以改变三者在紫萼玉簪幼苗叶片中的活性。从3种酶活性峰值可以看出,CAT对复合污染的敏感性明显高于POD,而POD对复合污染的敏感性明显高于SOD。在酸雨pH=2.5的各处理组合中,SOD活性呈现出持续增大的趋势,并且远远高于其他处理组合:表明高强度的酸雨能够大大激发SOD的潜在活性,从而有效地清除紫萼玉簪体内过剩的ROS,减少其对植物组织的损伤。这一结果与H2O2含量的提高相互印证。本试验还发现,紫萼玉簪幼苗在酸雨与Cd复合污染强度较小时,其体内产生的ROS会诱导SOD、POD以及CAT活性上升,从而减少ROS含量并防止逆境胁迫伤害[25,28]。这是因为SOD产生的H2O2刺激了POD和CAT的合成。当胁迫强度持续增大到超过植物的耐受限度时,紫萼玉簪幼苗体内ROS的产生速率超过了抗氧化酶系统的清除能力,对植物体内多种膜系统和酶系统造成破坏,POD和CAT活性开始呈现出下降趋势。这一结果与段文芳等[29]的研究结果一致。
植物遭受伤害和胁迫时,首先作用于细胞膜,产生活性氧,损伤细胞膜,导致质膜过氧化[6,30]。其中,H2O2不仅会对植物组织造成伤害,还会产生氧化能力极强的·OH和,进一步对细胞造成伤害。本试验结果表明,随着复合污染强度的增加,紫萼玉簪幼苗的H2O2含量提高,说明高强度复合污染加剧了植物体内的活性氧含量,植物细胞伤害严重。H2O2含量增加这一结果与SOD活性爆发性增长有着密切的关系,由于POD和CAT活性的丧失,无法将H2O2转化为无害物质,而是转化为·OH和,最终形成质膜过氧化的最终产物MDA。MDA是损伤细胞膜的活性物质,具有很强的细胞毒性,因此,MDA含量可作为植物抗逆性的一个生理指标[30-31]。高强度酸雨(pH≤3.5)促进紫萼玉簪细胞质膜过氧化反应,使MDA含量增加。这与本研究中H2O2含量变化相印证。在模拟酸雨和Cd胁迫下,虽然抗氧化酶系统被激活,在Cd达到一定强度后,紫萼玉簪幼苗体内抗氧化酶达到峰值,SOD出现爆发性增长,在一定程度上可以缓解胁迫伤害,但此时细胞膜仍然遭受到了巨大伤害,POD和CAT活性降低,无法清除体内大量的H2O2,因而紫萼玉簪自我保护能力被大大削弱。随胁迫强度的增大表现为电导率的持续增大[32],这也进一步说明ROS的产生速率超过了抗氧化酶系统的清除能力,幼苗完全丧失了自我保护的能力。
4 结论
4.1 酸雨和Cd复合污染比单一污染对紫萼玉簪种子和幼苗造成的伤害更大,二者间存在明显的交互作用,更多地表现为协同作用,且高强度复合污染对紫萼玉簪的影响更大。
4.2 紫萼玉簪种子和幼苗对低强度酸雨胁迫有一定的耐受性,临界值为pH=5.5,弱于成苗(pH=3.5)[6]。
4.3 紫萼玉簪种子和幼苗对低质量浓度Cd2+胁迫也有一定的耐受性,种子耐受的临界值为10 mg/L,弱于幼苗的耐受临界值(30 mg/L)。
总之,无论是复合污染还是单一污染,紫萼玉簪种子和幼苗都表现出一定的耐受性,但这种耐受性具有一定的阈值。紫萼玉簪种子和幼苗分别在酸雨(pH≥5.5)和Cd2+(≤10和30 mg/L)污染胁迫下能顺利进行自我更新,保证了该植物完成其生活史,为该植物种群在低强度复合污染下的繁衍提供了可能。
[1]齐泽民,王玄德,宋光煜.酸雨对植物影响的研究进展.世界科技研究与发展,2004,26(2):36-41. QI Z M,WANG X D,SONG G Y.Research progress of the effect of acid rain on plant.Journal of World Science and Technology Research and Development,2004,26(2):36-41.(in Chinese with English abstract)
[2]徐旭,孙振元,潘远智,等.园林植物对重金属胁迫的响应研究现状.世界林业研究,2007,12(6):36-38. XU X,SUN Z Y,PAN Y Z,et al.Review on responses of garden plants to heavy metal stress.Journal of World Forestry Research, 2007,12(6):36-38.(in Chinese with English abstract)
[3]周东美,王慎强,陈怀满,等.土壤中有机污染物-重金属复合污染的交互作用.土壤与环境,2000,9(2):143-145. ZHOU D M,WANG S Q,CHEN H M,et al.Interaction of organic pollutantsand heavy metalin soil.JournalofSoiland Environmental Sciences,2000,9(2):143-145.(in Chinese with English abstract)
[4]何翠屏.环境中重金属污染及其对植物生长发育的影响.青海草业,2004,13(2):26-29. HE C P.Influenced on plant growth and development by heavy metals in environment.Qinghai Prataculture,2004,13(2):26-29. (in Chinese with English abstract)
[5]刘兴乐,罗敬东,唐雪辉.紫萼玉簪的繁殖及应用.南方农业(园林花卉版),2008,2(8):76-77. LIU X L,LUO J D,TANG X H.Breeding and application ofHosta ventricosa.Journal of South China Agriculture,2008,2(8): 76-77.(in Chinese)
[6]王成聪,高素萍,黄丽,等.模拟酸雨与Cd对紫萼膜脂过氧化及形态特征的影响.草业学报,2014,23(6):336-341. WANG C C,GAO S P,HUANG L,et al.Effects of simulated acid rain and Cd on the membrane lipid peroxidation and morphological characteristics ofHosta ventricosa.Acta Prataculturae Sinica,2014,23(6):336-341.(in Chinese with English abstract)
[7]李合生.植物生理生化实验原理和技术.北京:高等教育出版社,2000:134-261. LI H S.Principles and Techniques of Plant Physiology and Biochemistry Experiment.Beijing:Higher Education Press,2000: 134-261.(in Chinese)
[8]PATTERSON B D,MACRAE E A,FERGUSON I B.Estimation of hydrogen peroxide in plant extracts using titanium(Ⅳ).Analytical Biochemistry,1984,139:487-492.
[9]熊庆娥.植物生理学实验教程.成都:四川科学技术出版社, 2003:72-78. XIONG Q E.Plant Physiology Experiment Course.Chengdu: Sichuan Science and Technology Press,2003:72-78.(in Chinese)
[10]郝再彬,苍晶,徐仲.植物生理实验.哈尔滨:哈尔滨工业大学出版社,2004:111-114. HAO Z B,CANG J,XU Z.Plant Physiology Experiment.Harbin: Harbin Institute of Technology Press,2004:111-114.(in Chinese)
[11]陶丽华,杨国栋,周青.不同水稻品种种子萌发对酸雨胁迫的响应.农业环境科学学报,2006,25(3):566-569. TAO L H,YANG G D,ZHOU Q.Responses of seed germination of different varieties of rice to the acid rain stress.Journal of Agricultural Environment Science,2006,25(3):566-569.(in Chinese with English abstract)
[12]张霞,李妍.铅胁迫对补血草种子萌发和幼苗初期生长及膜透性的影响.德州学院学报,2007,23(2):23-25. ZHANG X,LI Y.Effects of lead on the germination and growth ofLimonium sinense.Journal of Dezhou University,2007,23(2):23-25.(in Chinese with English abstract)
[13]杨颖丽,王文瑞,尤佳,等.Cd2+胁迫对小麦种子萌发、幼苗生长及生理生化特性的影响.西北师范大学学报(自然科学版), 2012,48(3):88-94. YANG Y L,WANG W R,YOU J,et al.Effects of cadmium stress on the germination,growth of seedling and physiological and biochemical characters of wheat.Journal of Northwest Normal University(Natural Science),2012,48(3):88-94.(in Chinese with English abstract)
[14]PATRA J,LENKA M,PANDA B B.Tolerance and CO-tolerance of the grassChloris barbataSw.to mercury,cadmium and zinc.New Phytologist,1994,128:165-171.
[15]曹德菊,汤斌.铅,镉及其复合污染对蚕豆根尖细胞的诱变效应.激光生物学报,2004,13(4):302-305. CAO D J,TANG B.Effect of plumbum,cadmium and the combined pollution actate on root tip cell ofVicia faba.Acta Laser Biology Sinica,2004,13(4):302-305.(in Chinese with English abstract)
[16]崔大练,马玉心,俞兴伟.Cd2+对海滨木槿种子萌发及根伸长抑制效应的研究.安徽农业科学,2011,39(8):4591-4593. CUI D L,MA Y X,YU X W.Inhibition effect of Cd2+on the seed germination and root elongation ofHibiscus hamabosiebet Zucc.Journal of Anhui Agricultural Sciences,2011,39(8):4591-4593. (in Chinese with English abstract)
[17]汪雅各,盛沛麟,袁大伟.模拟酸雨对土壤金属离子的淋溶和植物有效性的影响.环境科学,1988,9(2):22-26. WANG Y G,SHENG P L,YUAN D W.Effects of simulated acid rain on the leaching and plant availability of metal ions in soil.Environmental Science,1988,9(2):22-26.(in Chinese)
[18]马文丽,金小弟,王转花.镉处理对乌麦种子萌发幼苗生长及抗氧化酶的影响.农业环境科学学报,2004,23(1):55-59. MA W L,JIN X D,WANG Z H.Effects of cadmium on seed germination,growth of seedling and antioxidant enzymes of rye and wheat.Journal of Agro-Environment Science,2004,23(1):55-59.(in Chinese with English abstract)
[19]储玲,刘登义,王友保,等.铜污染对三叶草幼苗生长及活性氧代谢影响的研究.应用生态学报,2004,15(1):119-122. CHU L,LIU D Y,WANG Y B,et al.Effect of copper pollution on seedling growth and activate oxygen metabolism ofTrifolium pretense.Chinese Journal of Applied Ecology,2004,15(1):119-122.(in Chinese with English abstract)
[20]秦天才,吴玉树,黄巧云,等.镉铅单一和复合污染对小白菜抗坏血酸含量的影响.生态学杂志,1997,16(3):31-34. QIN T C,WU Y S,HUANG Q Y,et al.Effect of cadmium,lead single and combination pollution on the contents of ascorbic acid inBrassica chinensisL.Chinese Journal of Ecology,1997,16(3): 31-34.(in Chinese with English abstract)
[21]单运锋,冯宗炜,陈楚莹.模拟酸雨对七种森林树种生物量的影响.生态学报,1989,9(3):274-276. SHAN Y F,FENG Z W,CHEN C Y.The effects of simulated acid rain on biomass of seven species of forest plants.Acta Ecologica Sinica,1989,9(3):274-276.(in Chinese with English abstract)
[22]赵栋,潘远智,邓仕槐,等.模拟酸雨对茶梅生理生态特性的影响.中国农业科学,2010,43(15):3191-3198. ZHAO D,PAN Y Z,DENG S H,et al.Effects of simulated acid rain on physiological and ecological characteristics ofCamellia sasanqua.Scientia Agricultura Sinica,2010,43(15):3191-3198. (in Chinese with English abstract)
[23]李俊梅,王焕校.镉胁迫下玉米生理生态反应与抗性差异研究.云南大学学报(自然科学版),2000,22(4):311-317. LI J M,WANG H X.Eco-physiological responses and resistance to cadmium stress in three varieties of maize.Journal of Yunnan University(Natural Sciences),2000,22(4):311-317.(in Chinese with English abstract)
[24]陈良,隆小华,郑晓涛,等.镉胁迫下两种菊芋幼苗的光合作用特征及镉吸收转运差异的研究.草业学报,2011,20(6):60-67. CHEN L,LONG X H,ZHENG X T,et al.Effect on the photosynthetic characteristics of Cd uptake and translocation in seedlings of twoHelianthus tuberosusvarieties.Acta Prataculturae Sinica,2011,20(6):60-67.(in Chinese with English abstract)
[25]汤叶涛,关丽捷,仇荣亮,等.镉对超富集植物滇苦菜抗氧化系统的影响.生态学报,2010,30(2):324-332. TANG Y T,GUAN L J,QIU R L,et al.Antioxidative defense to cadmium in hyperaccumulatorPicris divaricata V.Acta Ecologica Sinica,2010,30(2):324-332.(in Chinese with English abstract)
[26]杜秀敏,殷文璇,赵彦修,等.植物中活性氧的产生及清除机制.生物工程学报,2001,17(2):121-125. DU X M,YIN W X,ZHAO Y X,et al.The production and scavenging of reactive oxygen species in plants.Chinese Journal of Biotechnology,2001,17(2):121-125.(in Chinese with English abstract)
[27]杨淑慎,高俊凤.活性氧、自由基与植物的衰老.西北植物学报, 2001,21(2):215-220. YANG S S,GAO J F.Influence of active oxygen and free radicals on plant senescence.Northwest Journal of Botany,2001,21(2): 215-220.(in Chinese with English abstract)
[28]龙云,刘芸,钟章成.酸雨和UV-B对玉米幼苗光合速率和抗氧化酶活性的影响.生态学报,2009,29(9):4956-4966. LONG Y,LIU Y,ZHONG Z C.Effects of UV-B irradiation and acid rain on photosynthetic rate and antioxidant enzyme activities of maize seedlings.Acta Ecologica Sinica,2009,29(9):4956-4966.(in Chinese with English abstract)
[29]段文芳,石贵玉,秦丽凤,等.镉胁迫对桐花树光合、蒸腾作用及保护酶活性的影响.安徽农业科学,2008,36(4):1355-1356. DUAN W F,SHI G Y,QIN L F,et al.Effects of Cd stress on the physiological indexes inAegiceras corniculatum.Journal of Anhui Agricultural Sciences,2008,36(4):1355-1356.(in Chinese with English abstract)
[30]孙胜,张智,卢敏敏,等.Cd2+胁迫对西瓜幼苗光合生理及活性氧代谢的影响.核农学报,2010,24(2):389-393. SUN S,ZHANG Z,LU M M,et al.Effects of cadmium stress on photosynthesis and active oxygen metabolism in the leaves of small watermelon seedlings.Journal of Nuclear Agricultural Sciences,2010,24(2):389-393.(in Chinese with English abstract)
[31]刘芸,钟章成,WERGER M J A,等.α-NAA和UV-B辐射对栝楼幼苗光合色素及保护酶活性的影响.生态学报,2003,23(1): 8-13. LIU Y,ZHONG Z C,WERGER M J A,et al.Effects of α-NAA and UV-B radiation on photosynthetic pigments and activities of protective enzymes inTrichosanthes kirilowiiMaxim leaves.Acta Ecologica Sinica,2003,23(1):8-13.(in Chinese with English abstract)
[32]张治安,王振民,徐克章.Cd胁迫对萌发大豆种子中活性氧代谢的影响.农业环境科学学报,2005,24(4):670-673. ZHANG Z A,WANG Z M,XU K Z.Effect of cadmium stress on active oxygen metabolism in germinated soybean seeds.Journal of Agro⁃Environment Science,2005,24(4):670-673.(in Chinese with English abstract)
Physio-ecological responses of seeds and seedlings of Hosta ventricosa to combined pollution of simulated acid rain and cadmium.
LUO Liangxu1,GAO Suping1,2*,WANG Chengcong1,LEI Ting1,2,WEN Jinyan1,LUO Yan1(1.College of Landscape Architecture, Sichuan Agricultural University,Chengdu 611130,China;2.Landscape Research Institute,Sichuan Agricultural University, Chengdu 611130,China)
cadmium;acid rain;combined pollution;ecological stress;resistant physiology;Hosta ventricosa
S 682.19
A
10.3785/j.issn.1008-9209.2016.05.192
Journal of Zhejiang University(Agric.&Life Sci.),2017,43(2):192-202
四川省教育厅项目“乡村景观植物蓝雪花的高效快繁体系建立研究”(JGYQ2016010);四川省科技计划项目“突破性竹类(花卉)育种材料与方法创新”(2016NYZ0038)。
高素萍(http://orcid.org/0000-0002-4609-9988),E-mail:gao_suping@sicau.edu.cn
(First author):罗良旭(http://orcid.org/0000-0001-8200-8281),E-mail:liangxuluo@163.com
2016-05-19;接受日期(Accepted):2016-08-23
猜你喜欢
杂志排行

浙江大学学报(农业与生命科学版)的其它文章
- Evaluation on formation rate of Pleurotus eryngii primordium under different humidity conditions by computer vision
- 流域非点源污染的最佳管理措施成本效益分析研究进展
- Association analysis revealed importance of dominance effects on days to silk of maize nested association mapping(NAM)population
- 检测乙酰微小杆菌的双重实时荧光定量聚合酶链式反应方法的建立
- 基于竞争性杂交方法的猪-肠道微生物特异性互作靶点发掘
- 应用分子标记辅助选育甘蓝型油菜杂交种的可行性