安徽查湾甜槠林不同大小林隙光因子对比分析
2017-05-17江国华汪秀琴宣城职业技术学院建筑艺术系安徽宣城242000
江国华, 汪秀琴 (宣城职业技术学院建筑艺术系,安徽 宣城 242000)
吴泽民 (安徽农业大学林学与园林学院,安徽 合肥 230036)
安徽查湾甜槠林不同大小林隙光因子对比分析
江国华, 汪秀琴
(宣城职业技术学院建筑艺术系,安徽 宣城 242000)
吴泽民
(安徽农业大学林学与园林学院,安徽 合肥 230036)
以安徽查湾自然保护区甜槠(Castanopsiseyrei)林为研究对象,分析不同大小林隙光因子变化情况。结果表明:不同大小林隙及非林隙林分内光照强度日变化呈单峰型,光照的峰值出现在11:00~13:00。在整个生长季中,光照和气温均表现为:大林隙>中林隙>小林隙>非林隙。不同大小林隙内光照由林隙中心区向林下逐渐递减,于近心区处减幅最大。
甜槠(Castanopsiseyrei);林隙;光照;安徽查湾
林隙是广泛存在于森林中的具有特殊性质的微结构,林隙的产生增强了干扰生境的异质性。Orians[1]、宋新章等[2]认为林隙环境本身是异质的,其物理环境的异质性促进生物环境的异质化,同时林隙内生物个体种群、群落对林隙干扰也会做出不同的响应,这些响应反过来又对林隙环境产生影响。
林隙干扰最直接、最重要的作用是提高生境中的光照条件。光照是植物进行光合作用并赖以生长的前提条件,也是影响森林植物生长生存的众多环境因子中的限制因子。林隙不但增加了光到达森林下层的持续时间,而且增加了生境内的光照强度。林隙的产生导致更多的光到达林地表面,这对热带森林的演替和更新有重大影响,也使林隙内地表面温度、近地层温度等热力特征发生相应的变化。而光照环境和热力特征的改变又影响土壤理化性质、营养元素分解、土壤微生物活性等的变化,最终影响到林隙及周边各生物学过程的变化[3~6]。因此,研究林隙光照特征有助于理解其小气候形成原因和群落变化规律,对深入研究林隙演替更新机制是必不可少的。
本研究以安徽査湾自然保护区甜槠林为研究对象,对不同大小林隙内光照特征进行初步分析,旨在为探讨林隙小气候形成机制及其对林隙生物多样性的影响等提供科学依据。
1 研究区概况与研究方法
1.1 研究区概况
研究区设在安徽省祁门县查湾自然保护区,位于皖南山区最南端,与江西接壤;地理位置为东经117°50′,北纬29°35′,面积1600hm2,海拔120~670m。全年温暖湿润,年平均气温15.6℃,年平均降水量1701.6mm,年无霜期达235d,相对湿度达85%。地带性土壤为黄壤和山地黄壤,成土母岩主要是千枚岩; 土层厚度在30~150cm之间;土壤腐殖质厚,pH在4.0~7.0之间。
植被属中亚热带常绿阔叶林带,主要森林群落类型为以甜槠(Castanopsiseyrei)、青冈(Cyclobalanopsisglaica)、苦槠(Castanopsissclerophylla)等为主要建群树种的常绿阔叶林,至今还保留较大面积的天然林分,是华东地区低海拔常绿阔叶林保存完好的地区之一。有维管束植物146科505属983种[7,8],主要乔木树种有甜槠、苦槠、青冈、马尾松(Pinusmassoniana)、枫香(Liquidambarformosana)、杉木(Cunninghamialanceolata)、江南桤木(Alnustrabeculosa)、红枝柴(Meliosmaoldhamii)、香桂(Cinamomumsubavenium)、木荷(Schimasuperba)、漆树(Toxicodendronvernicifluum)、米心水青冈(Fagusengleriana);灌木有乌药(Linderaaggregata)、马银花(Rhododendronovatum)、树参(Dendropanaxdentiger)、微毛柃(Euryahebeclados)、檵木(Loropetalumchinense)、格药柃(Euryamuricata)、山胡椒(Linderaglauca)、老鼠矢(Symplocosstettaris)、杨梅(Myricarubra)等。
1.2 研究方法
1.2.1 样方设置
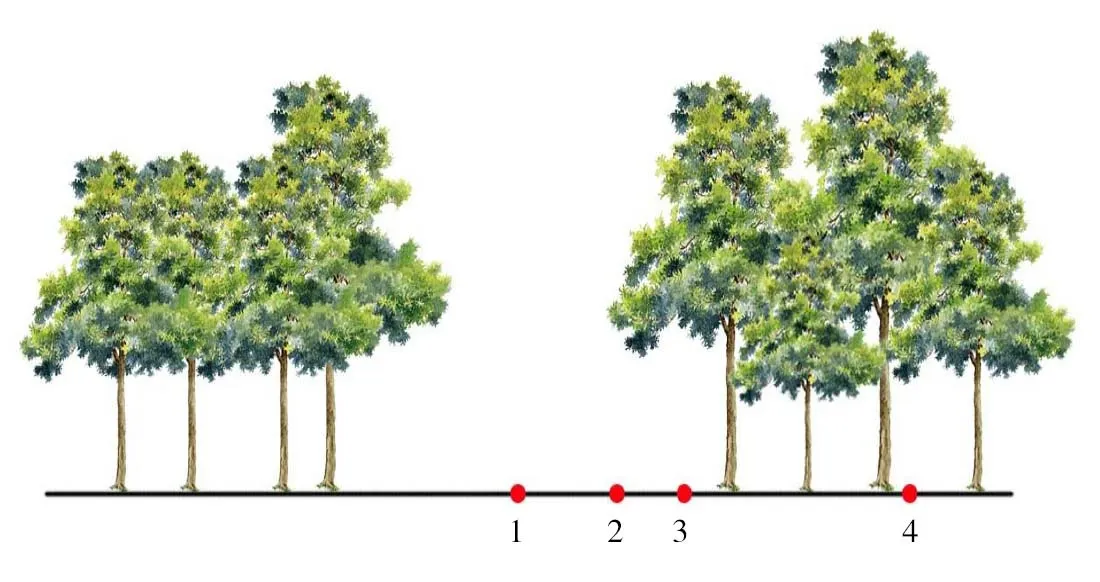
1.林隙中心区;2.林隙近心区;3.林隙边缘区;4.非林隙林分下图1 林隙观测点分布
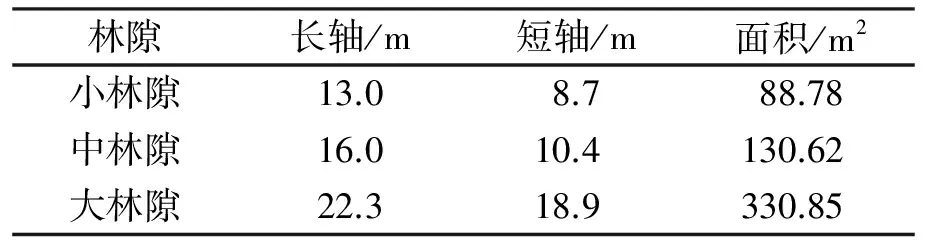
表1 观测点3个林隙的基本情况
所有研究样地设在自然保护区的缓冲区内,采用美国林务局制定的样线调查法[9]在安徽査湾自然保护区共调查36个林隙[10],根据林隙面积划分为3等级:小林隙<100m2、中林隙101~200m2、大林隙≥201m2,在每个等级中选择一个具代表性的林隙测量并设样地。每个林隙分别划分为林隙中心区(主要为林冠空隙)、林隙近心区(林冠空隙与扩展林隙的过渡区)、林隙边缘区(扩展林隙与非林隙的过渡区),同时在林冠层下设置非林隙对照区1个(图1)。所有研究林隙均位于阳坡,海拔为229m。观测点3个林隙的基本情况如表1所示。
1.2.2 光因子测定方法
于2014年4月~9月(生长季)每月选择晴好天气1~2d,从8:30至12:30每隔1h,从12:30至16:30每隔2h用ZDS-10自动换档数字式照度计沿水平梯度:林隙中心区、林隙近心区、林隙边缘区及林下分别测量光照强度(光照日变化取各林隙中心区光照强度,光照季节变化取各林隙中心区各观测时间光照强度的平均值)。
2 结果与分析
2.1 光照日变化
林隙内光照在一天内的变化是有规律的,不同大小的林隙及非林隙林分下不同,各有差异。安徽查湾甜槠林不同大小林隙及非林隙林分内光照强度的测定结果如图2所示。从光照日变化曲线可见,在一天内光照从早到晚呈现单峰型,即早晚光照较弱,而在中午11:00~13:00左右光照达到最高峰,这是各个时期光照变化都存在的规律。
由图2可知,4月18日光照在大林隙内较强,变化显著,其最大值达到3000 lx,变化幅度为924 lx,而在中、小林隙及林下都较接近,变化幅度也较小。夏季光照日变化显著,如7月14日光照在大、中、小及林下均存在较大差异,它们的变化幅度分别为2555、1473、959、459 lx。大林隙和小林隙光照变化幅度最大值在8月12日,分别为2913、1736 lx;中林隙和非林隙林分下隙光照变化幅度最大值在6月17日,分别为2775、929 lx。总的来说,大林隙、中林隙光照较接近,而小林隙和林下光照较接近。光照强度随林隙面积的增大而增大,林下最弱,变化幅度也随林隙面积的增大而增大。
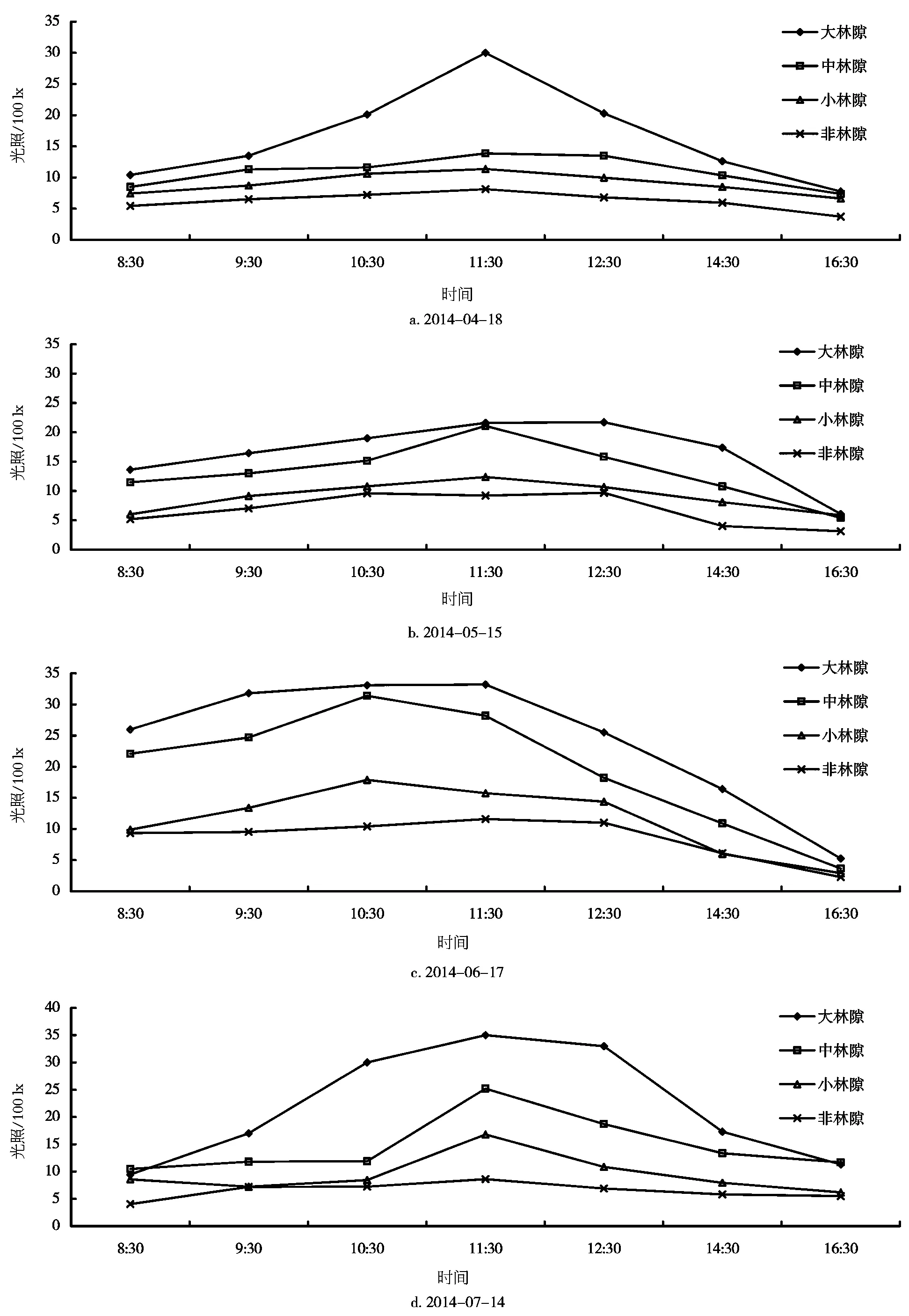
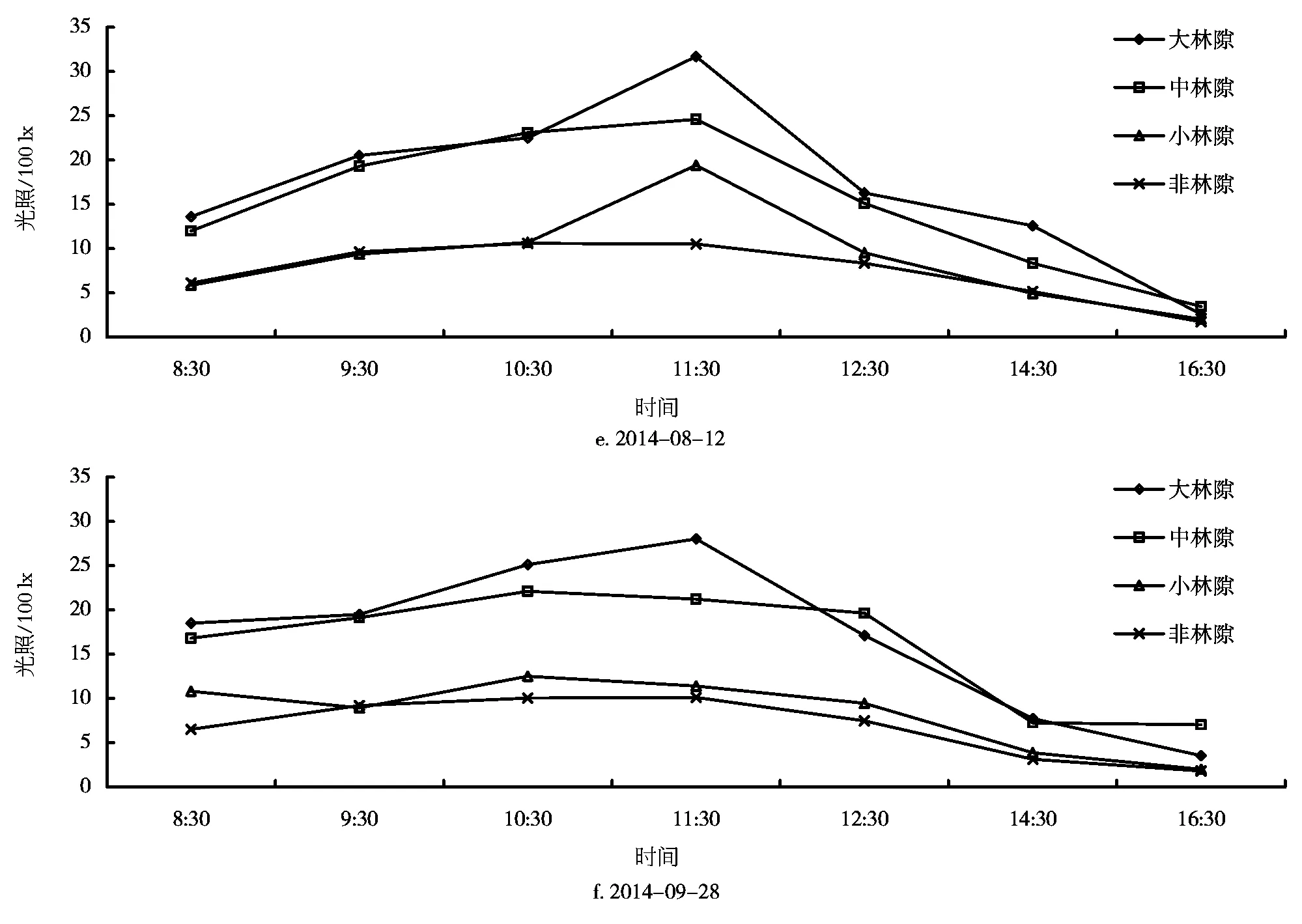
图2 不同大小林隙及林冠下光照日变化曲线
2.2 生长季内光照变化
如图3所示,日平均光照强度在生长季内的变化规律如下:生长初期到6月中下旬,林隙内与非林隙林分下光照都呈增强趋势,到6月中下旬达到高峰,以后又转而下降,即林隙内外日平均光照强度在生长季内的变化也呈单峰型。但到5月中旬,由于树叶的展开和枝条的繁茂,它们开始吸收,且阻碍了部分光线的直接射入,使得在这个时期光照增加的幅度有所下降。在这之前,绝大多数树叶尚未展开,随太阳高度角的增大,光照增强是必然的。树叶全部展开以后,随太阳高度与辐射的增强而增强,以后又逐渐下降。到9月下旬光照强度又略有增加趋势,这与皖南秋高气爽气候以及树势生长逐渐缓慢、枝叶吸收功能减弱有关。
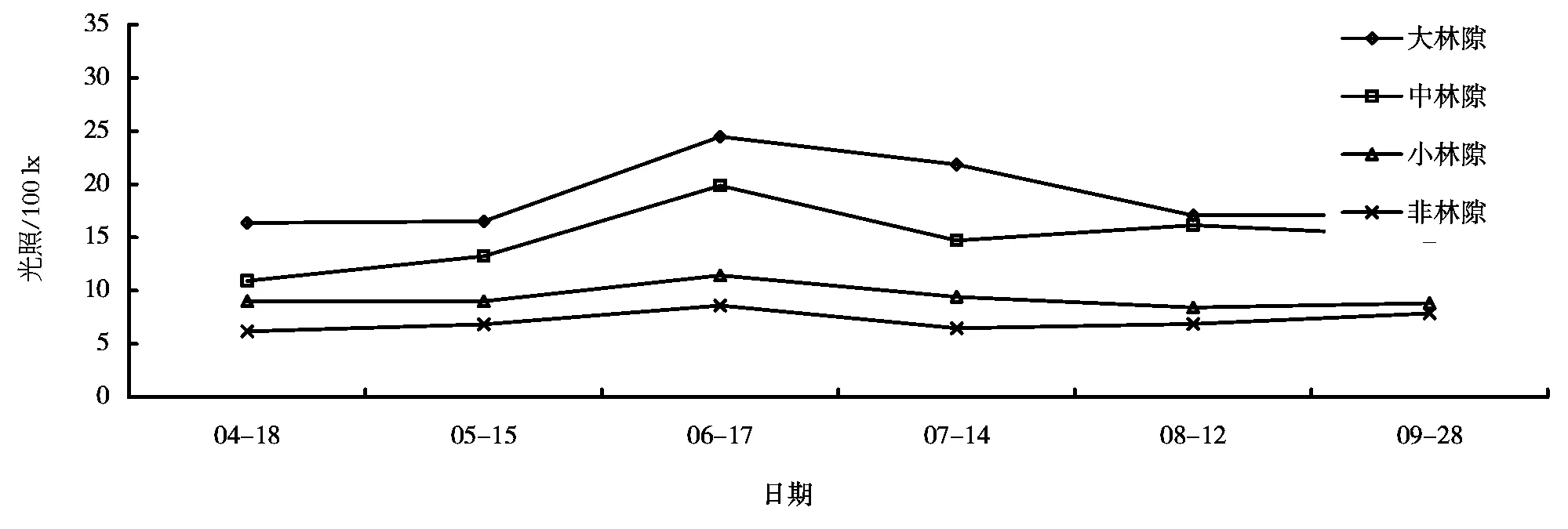
图 3 不同大小林隙及林冠下光照强度的季节变化曲线(2014年)
在观测日期内,不同大小林隙及非林隙林分下光照强度的最大值均在6月17日,分别为2447、1988、1146、862 lx;最小值除小林隙出现在5月15日(与4月20日仅相差3 lx),其它也都在4月18日。在整个生长季内,大、中、小林隙及林下光照平均值为1891、1501、936、714 lx,大、中林隙光照平均值显著高于小林隙和非林隙林分下。
2.3 光照随林隙梯度变化
林隙梯度是指从林隙中心区沿水平方向逐渐向林缘乃至非林隙林分下的位置变化梯度。不同时期、不同大小林隙的不同区域,其光照强度不同。图4为不同时期各观测点日平均光照随林隙梯度变化曲线。大、中、小林隙内光照强度沿水平梯度总的变化规律如下:由林隙中心向林下光照呈递减趋势,从林隙中心区到林隙近心区递减幅度明显大于其它各区域。
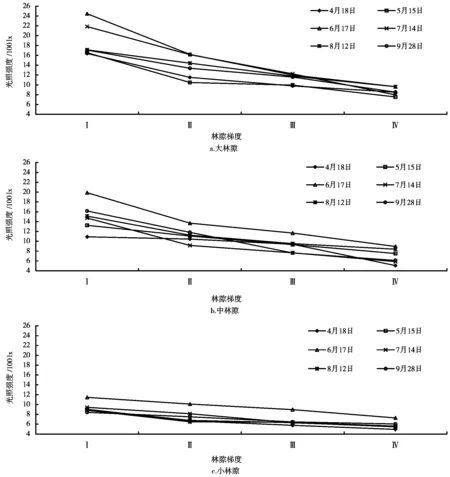
Ⅰ.林隙中心区;Ⅱ.林隙近心区;Ⅲ.林隙边缘区;Ⅳ.非林隙林分下图4 不同大小林隙光照强度的林隙梯度变化
但此递减的程度因林隙面积的大小而有区别,大、中、小林隙,其附近林下与林隙中心区光照强度平均值的比值分别为45.6%、48.6%、62.2%,林隙中心区与附近林下光照强度平均值的差值分别为1029、748 lx和354 lx,这说明随着林隙面积的增大,林隙中心区的光照明显增强,与林下相比,增幅明显增大;其次,这种变化还与各林隙更新层等因素有关。大、中林隙内光照在生长初期(4月份)、生长末期(9月下旬、10月上旬)随林隙梯度变化幅度相对较小,而在生长中期(6、7月份)变化幅度较大;小林隙内光照在整个生长期随林隙梯度变化幅度均相对较小。这与林隙边缘树冠透光率、太阳高度角、方位角、林冠结构、坡度坡向、观测点位置等因子有关。其中,5月13日日平均光照相对较低,主要原因有二:一是林隙边缘有少量落叶树种如枫香,更新层中有较多的落叶树江南桤木,此时已有新叶长出,从而遮挡了部分光照:二是该日天空有少量薄云,光照也受到一定程度的影响。
光照强度随林隙梯度的变化,使林隙不同区域接受光能辐射量的大小不同,导致水热情况不均,林隙生态环境存在异质性,从而影响林隙不同区域种子萌发、幼苗生长和树种的更新,使林隙不同区域物种多样性不一样。
3 讨论与小结
光因子是林隙生态环境重要的生态因子,它会影响林隙内的温度、湿度、土壤养分状况等其他因子。林隙生态环境研究是为林隙干扰和林隙更新研究提供机理性的数据,了解林隙内生态因子的变化规律是分析林隙更新机制的基础。将林隙的干扰状况、林隙生态因子的变化规律与树种的生物学特性结合起来全面分析,就能深入了解森林群落的动态更新、物质共存与多样性维持机制。
安徽查湾甜槠林林隙内光照强度日变化呈单峰型,光照最大值出现在11:00~13:00,大、中型林隙光照较接近,而小林隙和非林隙林分下光照较接近。光照强度总体上表现为:大林隙>中林隙>小林隙>非林隙。日平均光照强度在生长季内的变化也呈单峰型,最大光照强度出现在6月中下旬。最大值的来临较长白山阔叶红松林及南亚热带常绿阔叶林林隙为早[6]。这与所选样地处在阳坡有很大关系。
光照在林隙内与非林隙林分下,不论在日变化还是在生长季内的变化均是有差异的,并且有一定的规律性,林隙内与非林隙林分下光照有明显差异,林隙内的光照强度都明显地高于非林隙林分下,而林隙之间由于面积大小的差异亦多少会造成光照的不同。不同大小林隙内光照由林隙中心区至林下逐渐递减,于近心区处减幅最大。可见,近心区是林隙内光照强度变化的一个转折点,是林隙生态环境发生异质性变化的关键区域。
[1] Orians G H.The influence of tree-falls in tropical forests in tree species richness[J].Tropical Ecology,1982,23:255~279.
[2] 宋新章,肖文发.林隙微生境及更新研究进展[J].林业科学,2006,42(5):114~119.
[3] 张一平,王进欣,刘玉洪,等.热带次生林林窗干热季光照特征初步分析[J].广西植物,2001,21(1):1~8.
[4] Canham C D.An index for understory light levels in and around canopy gaps[J].Ecology,1989,69:786~795.
[5] 王进欣,张一平.林窗微环境异质性及物种响应研究综述[J].热带植物研究,2000,46:23~31.
[6] 臧润国,刘静艳,董大芳.林隙动态与森林生物多样性[M].北京:中国林业出版社,1999.
[7] 吴征镒.中国植被[M].北京:科学出版社,1980:321~323.
[8] 安徽植被协作组.安徽植被[M].合肥:安徽林业科技出版社,1981:133~156.
[9] Runkle J R.Pattern of disturbance in some old-growth mesic forests of eastern North America[J]. Ecology,1982,63:533~546.
[10] 江国华,吴泽民,沈后海,等.安徽查湾自然保护区甜槠林林隙形成及特征研究[J].安徽农业大学学报,2008,11(3):229~235.
[编辑] 余文斌
2016-10-31
国家“十一五”科技支撑项目(2006BAD03A06);安徽省高校自然科学重点研究项目(KJ2014A244);安徽省高校自然科学重点研究项目(KJ2016A785)。
江国华(1973-),男,硕士生,副教授,研究方向为森林生态与植物生理生态。通信作者:吴泽民,jghjrz@163.com。
S718.45
A
1673-1409(2017)06-0018-06
[引著格式]江国华, 汪秀琴,吴泽民.安徽查湾甜槠林不同大小林隙光因子对比分析[J].长江大学学报(自科版),2017,14(6):18~23.
