滇西北剑湖湿地海菜花(Otteliaacuminata)群落物种组成及种群分布格局*
2017-05-17李宁云陈玉惠胡金明刘朝蓬敖新宇
李宁云,陈玉惠,胡金明,刘朝蓬,李 靖,敖新宇,雷 然
(1:云南大学,昆明 650091) (2:云南省林业科学院,昆明 650201) (3:西南林业大学,昆明 650224) (4:国家高原湿地研究中心,昆明 650224)
滇西北剑湖湿地海菜花(Otteliaacuminata)群落物种组成及种群分布格局*
李宁云1,2,陈玉惠3,胡金明1,刘朝蓬4,李 靖3,敖新宇3,雷 然3
(1:云南大学,昆明 650091) (2:云南省林业科学院,昆明 650201) (3:西南林业大学,昆明 650224) (4:国家高原湿地研究中心,昆明 650224)
群落物种组成及种群分布格局特征是揭示种群发展趋势及与环境相互关系的基础. 以滇西北剑湖湿地海菜花群落为对象,采用样方法与扩散系数法对其物种组成与种群分布格局进行研究. 结果表明:群落共有物种18种,分属12科、14属,包括沉水、漂浮、浮叶及湿生4类生活型;海菜花(Otteliaacuminata)、豆瓣菜(Nasturtiumofficinale)、马来眼子菜(Potamogetonmalaianus)和草茨藻(Najasgraminea)呈集群分布,其余种群呈随机分布;海菜花种群个体间拥挤效应最大,种内竞争最为激烈. 保育海菜花种源、开展流域和生境治理以减缓湖泊沼泽化进程,是目前剑湖海菜花群落保护的当务之急.
物种组成;种群分布格局;海菜花群落;湖泊沼泽化;剑湖湿地
种群分布格局研究有助于揭示种群发展趋势以及与环境的相互关系[1]. 研究表明,湿地克隆植物由侵入期、定居初期到种群发育盛期的过程中,集群程度先增大后减小,呈扩散的趋势;种子植物从幼苗到繁殖期,集群程度增大,呈聚集的趋势[2]. 湿地环境变化对种群分布格局会产生显著影响. 如由于人工沙垄的阻隔,失去了潮汐作用,导致分布于潮间带的珊瑚菜(GlehnialittoralisFr. Schmidt ex Miq.)聚集范围变窄、聚集强度变弱[3].
云南高原湖泊以海菜花(Otteliaacuminata)群落为优势沉水植物群落,该群落为云南高原湖泊湿地特有,是云南高原湖泊湿地的象征[4]. 海菜花属于水鳖科 (Hydrocharitaceae) 大型沉水单子叶植物,为云南省Ⅲ级保护植物[5-7]. 由于地理隔绝,海菜花在云南高原湖泊湿地形成4个变种,即原变种(O.acuminatavar.acuminata)、波叶海菜花(O.acuminata(Gagnep.) Dandy var.crispa(Hand.-Mazz.) H. Li)、通海海菜花(O.acuminata(Gagnep.) Dandy var.tonghaiensisH. Li) 和长湖海菜花(O.acuminata(Gagnep.) Dandy var.lunanensisH. Li);各变种形成的群落特征有所差异[5]. 自1960s以来,海菜花群落分布的滇池、洱海等湖泊湿地群落相继消亡[8-9]. 我们对滇西北洱海、茈碧湖、剑湖、拉市海和泸沽湖5个高原湖泊的研究表明[10],近30年来,天然海菜花群落仅在泸沽湖和剑湖有分布,呈现出群落物种数增加、生活型构成复杂、物种更替等变化,其中以剑湖海菜花群落上述特征变化较大;但对于种群分布格局特征尚不清楚. 因此,本文以剑湖湿地海菜花群落为对象,明确其物种组成与种群分布格局特征,为进一步揭示种群发展趋势以及与环境的相互关系奠定基础,以期为珍稀濒危物种及高原湖泊湿地的保护提供科学依据.
1 研究区概况
剑湖湿地(26°28′N,99°55′E)位于云南西北部剑川县城东南3 km处,海拔2188 m,面积623 hm2,是在新构造运动作用下,于陷落地段积水形成的断陷湖. 区域气候属北亚热带湿润气候,年均温12.3℃,年降水776.7 mm,集中于6-9月(占全年降水的77%). 水系属澜沧江水系,水量补给主要由降雨以及永丰河、金龙河、格美河、狮河等河流汇入,湖水由海尾河注入漾濞河后汇入澜沧江. 剑湖生物多样性极其丰富,黑鹳(Ciconianigra)、黑颈鹤(Grusnigricollis)、金雕(Aquilachrysaetos)、中华秋沙鸭(Mergussquamatus)等是国家Ⅰ级保护鸟类;光唇裂腹鱼(Schizothoraxlissola-biatus)、云南裂腹鱼(S.yunnanensis)、后背鲈鲤(Percocyprispingi-retrodorslis)和剑湖高原鳅(Triplophysajianhuensis)等是特有物种,其中剑湖高原鳅是1980s才发现的新种,为剑湖特有[11]. 剑湖海菜花群落的优势种为海菜花原变种,当前仅有小面积残存于格美河入湖口附近和金龙河入湖口附近,其分布水深范围为59~73 cm(均值为72.33±8.54 cm).
2 研究方法
2.1 研究对象与研究区选取
云南高原湖泊分布的4个海菜花变种,原变种分布最为广泛,波叶海菜花仅分布于泸沽湖,通海海菜花仅分布于通海杞麓湖和宜良阳宗海,长湖海菜花仅分布于石林长湖[4-5]. 原变种海菜花群落在云南高原湖泊中具有普遍性与代表性,选取其为研究对象.
通过近年对滇池、洱海、洱源西湖、茈碧湖、剑湖、拉市海等原变种分布湖泊的考察,结合文献资料[8-9,11-12]分析,以及对湿地管理部门的访谈,确定上述湖泊湿地中仅有剑湖分布有天然海菜花原变种群落. 因此,选取剑湖为研究区.
从剑湖海菜花群落演变历史看,在1970s剑湖海菜花群落呈大面积分布,为优势水生植物群落[11];但近年海菜花群落在剑湖的分布范围退缩. 我们2011年对剑湖的考察,结合1977年李恒的调查[12]、2005年西南林学院科考[11],表明格美河入湖口在1977、2005和2011年均分布有海菜花群落,金龙河入湖口在2005年和2011年均分布有海菜花群落. 两处海菜花群落是在长期的环境变化和人类活动中保留下来的,存续时间长,其现时物种组成与种群分布格局包含了丰富的环境变化和人类活动信息,明确其物种组成与种群格局特征,对于进一步揭示种群发展趋势以及与环境的相互关系具有重要意义. 因此,选取格美河入湖口与金龙河入湖口为剑湖海菜花群落调查区域.
2.2 野外调查
从水热组合来看[11],6-9月为剑湖流域降水丰沛、热量高的时段;该时段的调查能够获得较为全面的湿地植物种类信息. 因此,于2011年8月对剑湖海菜花群落开展野外调查.
剑湖现时海菜花群落分布范围退缩、面积小,难以采用系统布点法[13]对群落进行调查. 因此,采用法瑞学派的主观取样法[14],基于群落外貌、结构、种类和生境基本一致的原则确定取样点,布设调查样方;调查共布设1 m×1 m 样方9个,满足法瑞学派一个群丛取样方3个以上,6~12个最佳的取样要求[14]. 样方调查记录物种组成、生活型、物候、水深等指标.
2.3 数据分析
基于调查数据,统计和计算群落数量指标(相对频度、相对盖度、重要值、生活型及数量). 以重要值(相对频度+相对盖度)为测度指标,采用扩散系数(C)及其t检验和x2检验(自由度df=N-1,95%置信度)[15-17]对种群的分布格局进行测定,考虑检验方法的差异及密度的影响,辅以负二项参数(k)[18]进行验证. 参照研究成果[17,19],选取平均拥挤度(m*)、聚块指数(PAI)、Cassie指数(CA)、格林指数(GI)等测定种群的聚集强度.
1)扩散系数(C):
(1)

t检验:
(2)
x2检验:
x2=C(N-1)
式中,N为样方数,以N-1的自由度和95%的置信度分别查表对比,检验显著性.
2)负二项参数(k):k<0时种群随机分布;k>0时,种群呈集群分布,且随集群程度的增加k值减小.
(3)
3)平均拥挤度(m*):表示平均每个个体有多少个在同单位的其他个体,数值越大,拥挤效应越大.
(4)
4)聚块指数(PAI):不涉及密度影响,值越大聚集性越强.
(5)
5)Cassie指数(CA):当CA→0为随机分布,当CA>0时为集群分布,当CA<0时为均匀分布.
CA=1/k
(6)
6)格林指数(GI):当GI=0时为随机分布,当GI=1时为最大集群分布.
GI=(C-1)/(N-1)
(7)
3 结果与分析
3.1 物种组成
3.1.1 科、属、种组成 样方调查表明,剑湖海菜花群落共有物种18种,分属12科、14属(表1). 从科的分类阶元上来看,含单属的共10科,包括十字花科(Cruciferae)、眼子菜科(Potamogetonaceae)、禾本科(Poaceae)、蓼科(Polygonaceae)、茨藻科(Najadaceae)、钱苔科(Ricciaceae)、睡菜科(Menyanthaceae)、狸藻科(Lentibulariaceae)、萍科(Marsileaceae)和小二仙草科(Haloragidaceae),占总科数的83.33%;含2个属的科共2科,包括水鳖科(Hydrocharitaceae)和浮萍科(Lemnaceae),占总科数的16.67%. 从属的分类阶元来看,含单种的属共计11属,包括水车前属(Ottelia)、黑藻属(Hydrilla)、紫萍属(Spirodela)、狐尾藻属(Myriophyllum)、假稻属(Leersia)、狸藻属(Utricularia)、茨藻属(Najas)、萍属(Marsilea)、豆瓣菜属(Nasturtium)、浮苔属(Ricciocarpus)和荇菜属(Nymphoides),占总属数的78.57%;含2个种的属共2属,包括蓼属(Polygonum)和浮萍属(Lemna),占总属数的14.29%;含3个种的属只有1属,即眼子菜属,占总属数的7.14%. 海菜花群落物种组成呈现出单科单属比例大、物种组成较多的特点.
3.1.2 生活型构成 群落由沉水、漂浮、浮叶及湿生4类植物构成,以沉水植物占优势,其种数为9个;其次为漂浮植物,种数为4个;湿生植物和浮叶植物种数分别为3个和2个(表1);反映了物种生活型齐全、群落层次丰富的特点.

表1 剑湖海菜花群落物种及生活型构成
3.1.3 数量特征 群落中以优势种海菜花的重要值最大,其次是豆瓣菜,但与海菜花重要值相差较大,表明海菜花在群落中占有绝对优势地位;再次为马来眼子菜和李氏禾;其余14种物种重要值较低,重要值范围为1.52~10.66,其中酸模叶蓼和两栖蓼的重要值分别为6.97和6.56,在14种物种中相对较高(表2);李氏禾、酸模叶蓼和两栖蓼等湿生植物具有较高的重要值,表明其在群落中具有相对较高的地位和作用.

表2 剑湖海菜花群落种群数量特征
3.2 种群分布
3.2.1 种群分布格局 对海菜花群落种群分布格局的测定可看出,群落组成物种呈现出两类种群分布格局,即集群分布与随机分布. 优势种海菜花种群和重要值较大的伴生种豆瓣菜、马来眼子菜和草茨藻种群呈集群分布,表明这些种群以集群方式占有生态空间,呈斑块状;李氏禾、眼子菜、酸模叶蓼等14个种群则呈现随机分布(表3).

表3 剑湖海菜花群落种群分布格局1)

3.2.2 聚集强度 对群落内各种群聚集强度的测定(表4)表明,集群分布种群4项指数均较随机分布种群大,表明其聚集性强,个体间拥挤效应大,种内竞争激烈. 其中,海菜花种群m*最大,表明其个体间拥挤效应最大、种内竞争最为激烈;草茨藻聚集性最强,但其个体间拥挤效应最小,种内竞争相对缓和.
随机分布种群,以李氏禾4项指数最大,m*甚至比集群分布的草茨藻大,表明其聚集性较强、种内竞争较为激烈. 但GI和GA的值与指数含义有差异,表明这两项指数用于量化种群聚集强度的效果较差.
4 讨论
水文条件是决定湿地植物群落物种组成和丰富度的主导因素[20]. 由于森林植被的破坏,当前剑湖流域水土流失面积达10545 hm2,占径流区总面积的21.7%,年均产生淤积10.67×104m3[21];周边社区种植茭草(Zizanialatifolia)以获取牲畜饲料,沿湖形成了茭草单优群落的湖滨带,拦截了泥沙[22-23];湖周垦殖以及环湖公路的建成,切割地表径流并压缩了径流空间,其呈散流状态的汇入形式发生改变,缩减了径流补给湖滨带的面积. 这一系列的变化使得剑湖湖底淤积、湖盆变浅、湖滨带萎缩,沼泽化进程加剧. 李恒1977年调查海菜花群落生境水深为1.1、1.5 m[12],2005年西南林学院科考调查的生境水深为1.1 m[11],本次调查(2011年)发现其水深范围为59~73 cm. 水位大幅下降,生境条件变得严酷,虽未改变沉水植物群落的属性,但为李氏禾、酸模叶蓼、两栖蓼等湿生物种提供了生长条件;从水环境的变化来看,湖体水质由1990s的Ⅱ类发展为当前的Ⅲ类[21],为耐污的浮萍科植物提供了生长条件.
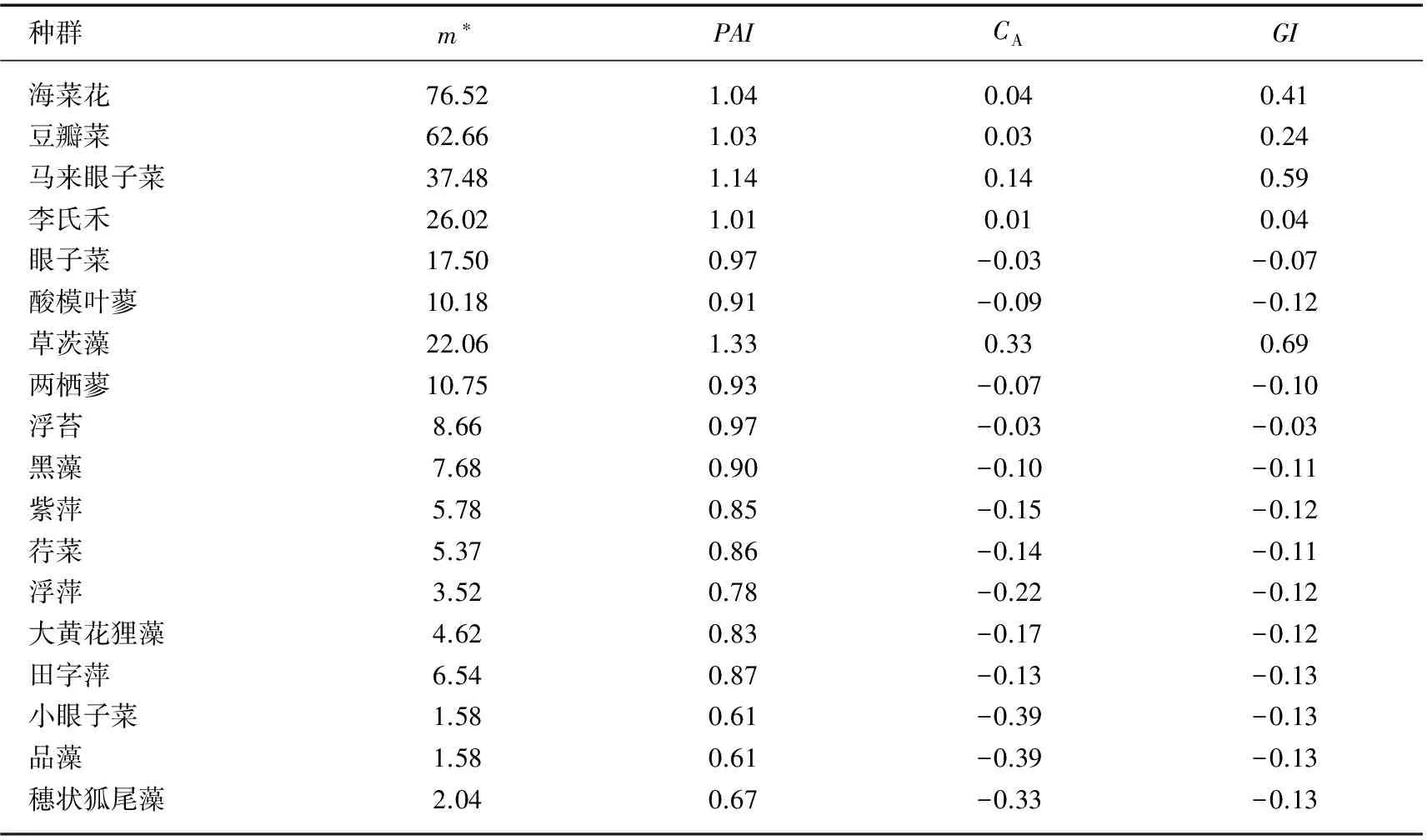
表4 剑湖海菜花群落种群聚集强度
剑湖海菜花群落生境由深水向浅水发展,沉水植物的优势被削弱;同时,周边社区对水生植物的大量打捞使群落空余较多的生态位,为适宜物种的侵入提供了有利条件;植物种类的增加,使生态位重新进行分化和组合,群落出现分层现象,生活型种类增多. 从近30年剑湖海菜花群落生活型构成的变化来看,呈现出由1977年的单一沉水植物构成向2005年的沉水、浮叶和漂浮植物构成[11-12],直至2011年的沉水、浮叶、漂浮和湿生植物构成的变化(图1). 沉水植物比例由1977年的100%降至2005年的70%,2011年降至50%,反映了优势生活型物种在群落中地位的削弱;漂浮植物与浮叶植物在2005年出现,前者比例由2005年的20%增至2011年的22.22%,后者比例由2005年的10%增至11.11%;湿生植物在2011年出现,比例为16.67%(图1);优势生活型物种比例的下降,多种生活型物种的出现及比例的变化,反映了生境由均一向复杂、多样化的变化趋势. 研究表明[24-26],在水生生态系统向陆生生态系统的演替中,物种丰富度呈增加的趋势. 剑湖现时海菜花群落物种组成较多、生活型构成多样化的特点反映了水生生态系统的逆向生态演替,与30年前群落物种较少、生活型单一有本质的区别.
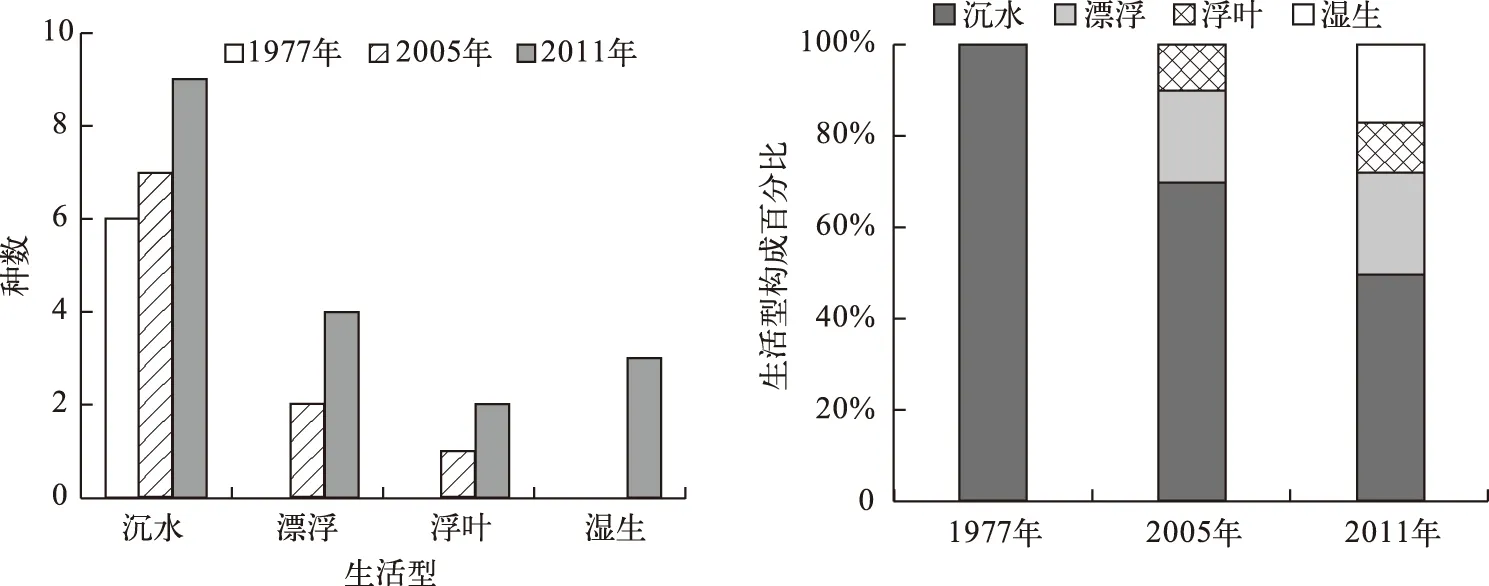
图1 剑湖海菜花群落生活型及其构成变化Fig.1 Changes of life form and its composition of Ottelia acuminata community in Lake Jianhu
对种群分布格局的测定表明,群落中优势种群海菜花和重要值较大的伴生种群豆瓣菜、马来眼子菜和草茨藻呈集群分布,其余伴生种群均呈随机分布. 有性繁殖和无性繁殖交替是沉水植物繁殖方式对环境的适应策略[27],当前剑湖残存的两处海菜花生境均处于湖滨带,相对封闭、水流平缓,限制了种子的传播范围与距离,有利于在小尺度空间形成集群分布;同时,湖滨带的过渡性使环境资源的空间分布呈斑块状,“环境筛”作用增强,为适宜物种的集群分布创造了环境资源条件;在种子完成扩散和繁殖后,沉水植物通过无性繁殖增强资源利用能力,进而提高竞争能力,维持并拓殖占据的空间[27],而无性繁殖所延伸的空间范围更为狭小,促进了种群集群分布. 海菜花为优势种群,种间竞争对其种群的影响较小;豆瓣菜、马来眼子菜和草茨藻的重要值较大,种间竞争压力的影响也相对较小,在上述生境条件下形成集群分布. 但是,海菜花种群个体间拥挤效应最大,导致种内竞争大于种间竞争. 随种群密度的增加或环境的改变如水位的持续下降,会使种群的分布格局发生演变,这需要进一步研究.
在滨湖湿地环境里,克隆繁殖产生的无性系,其形态可塑性、生理整合性等生态对策有利于提高种群个体的生存和竞争力,较种子繁殖生态适合度高,具有进化优势[2]. 李氏禾是以无性繁殖为主的湿生植物,其地下茎萌芽能力强[28]. 由于每个分株群的增大,导致聚集性增强. 种群集群分布可以促进种间共存,允许群落容纳更多的物种,增加丰富度和多样性[29]. 剑湖海菜花群落集群分布的均为优势种群和重要值较大的伴生种群,对群落环境资源的占有较大,导致其余种群种间竞争压力增大,种群世代更替减缓,停滞于对生境适应的早期阶段,呈随机分布格局;随机分布是种群适应严酷环境的初级阶段,种群内部迟早会形成不同程度的集群[19].
从湖泊演替规律来看[30],剑湖由湖泊向沼泽发展并最终演替为陆生生态系统是不可避免的. 但从当前剑湖的情况来看,人类活动干扰加速了这一过程. 因此,减缓人类活动干扰对生境的改变是海菜花群落保护的关键. 针对当前的人类活动干扰,应基于断陷湖泊,湿地环境较为封闭,且受流域影响较大的特征,从流域尺度上治理水土流失,减缓湖泊沼泽化和水环境恶化的趋势;基于剑湖湖盆-湖滨-面山的空间结构特征,恢复与重建湖滨带,构建湖泊生态安全屏障. 对现时海菜花群落的保护,应根据种群分布格局,采取不同的管理措施维持群落的稳定. 首先,应维持优势种群的繁荣与稳定,应针对海菜花种群高聚集性的特点,采取移植措施对海菜花种源进行保育;其次,应降低湿生植物在群落中的地位与作用,针对生境位于河流入湖口湖滨带的位置,通过拦砂坝等措施拦截入湖泥沙,减缓生境的沼泽化进程.
5 结论
剑湖湿地海菜花群落呈现出物种组成较多,生活型构成齐全,优势种海菜花和重要值较大的豆瓣菜、马来眼子菜和草茨藻呈集群分布,部分湿生物种聚集性强的特点. 这是对水生生境逆向生态演替的反映. 采取移植措施保育海菜花种源并开展流域与生境治理以减缓生境的沼泽化进程,是目前剑湖海菜花群落保护的关键和当务之急.
[1] Li Wei, Wang Ruixue, Zhang Guangfuetal. Point pattern analysis ofex-situpopulation ofTaxuswallichianavar.mairei.ChineseJournalofEcology, 2014, 33(1):16-22.[李伟, 王瑞雪, 张光富等. 南方红豆杉迁地保护种群的点格局分析. 生态学杂志, 2014, 33(1): 16-22.]
[2] Huang Zhiwei, Peng Min, Chen Guichenetal. The spatial distribution patterns and dynamics of some wetland dominant plants of the Qinghai Lake.ChinaJApplEnvironBiol, 2001, 7(2): 113-116.[黄志伟, 彭敏, 陈桂琛等. 青海湖几种主要湿地植物的种群分布格局及动态. 应用与环境生物学报, 2001, 7(2): 113-116.]
[3] Zhang Min, Yang Hongxiao. Spatial patterns ofGlehinalittoralispopulation on sandy seashore and their responses to artificial beach ridge.ChineseJournalofEcology, 2015, 34(1): 47-52. [张敏, 杨洪晓. 滨海沙滩珊瑚菜种群的空间格局及其对岸垄的响应. 生态学杂志, 2015, 34(1): 47-52.]
[4] Jiang Zhutan, Li Heng, Dao Zhilingetal. Ethnobotanical study onOtteliaacuminata, an aquatic edible plant occurring in Yunnan.JournalofInnerMongoliaNormalUniversity(NaturalScienceEdition), 2010, 39(2):163-168. [蒋柱檀, 李恒, 刀志灵等. 云南传统食用植物海菜花(Otteliaacuminata) 的民族植物学研究. 内蒙古师范大学学报 (自然科学版), 2010, 39 (2): 163-168.]
[5] Li Heng. Classification, distribution and phylogeny of the genusOttelia.ActaPhytotaxonomicaSinica, 1981, 19(1): 29-42.[李恒. 海菜花属的分类、地理分布和系统发育. 植物分类学报, 1981, 19(1): 29-42.]
[6] Forestry Department of Yunnan Province. The first batch of rare and endangered plants list of Yunnan province.YunnanForestry, 1985, 2: 2-3.[云南省林业厅. 云南省第一批珍稀濒危保护植物名录. 云南林业, 1985, 2: 2-3.]
[7] Forestry net of China. Rare and endangered plants list of China. http://www.forestry.gov.cn/portal/zrbh/s/3053/content-457748.html.[中国林业网.中国珍稀濒危植物名录. http://www.forestry.gov.cn/portal/zrbh/s/3053/content-457748.html.]
[8] Li Heng. The flourishing and decinling ofOtteliaacuminatein the lake of Dian Chi.JournalofYunnanUniversity, 1985, 7(supplement): 138-142.[李恒. 滇池海菜花的盛衰.云南大学学报, 1985, 7(增刊): 138-142.]
[9] Fu Hui, Yuan Guixiang, Cao Teetal. Succession of submerged macrophyte communities in relation to environmental change in Lake Erhai over the past 50 years.JLakeSci, 2013, 25(6): 854-861. DOI: 10.18307/2013.0609.[符辉, 袁贵香, 曹特等. 洱海近50a来沉水植被演替及其主要驱动要素. 湖泊科学, 2013, 25(6): 854-861.]
[10] Li Ningyun, Tian Kun, Chen Yuhuietal. Changes ofOtteliaacuminatacommunities in the plateau lakes of northwestern Yunnan over the past three decades.JLakeSci, 2015, 27(3):401-406. DOI: 10.18307/2015.0305.[李宁云, 田昆, 陈玉惠等. 近30年滇西北高原湖泊海菜花群落变化. 湖泊科学, 2015, 27(3): 401-406.]
[11] Southwest Forestry College ed. Comprehensive scientific survey report of Yunnan Jianhu Nature Reserve. Kunming: Southwest Forestry College, 2005: 53-54, 122.[西南林学院. 云南省剑湖湿地自然保护区综合科学考察报告. 昆明: 西南林学院, 2005: 53-54, 122.]
[12] Li Heng. A study on the lake vegetation of Yunnan Plateau.ActaBotanicaYunnanica, 1980, 2(2):113-118.[李恒. 云南高原湖泊水生植被的研究. 云南植物研究, 1980, 2(2): 113-118.]
[13] Fang Jingyun, Wang Xiangping, Shen Zehaoetal. Methods and protocols for plant community inventor.BiodiversityScience, 2009, 17(6): 533-548. [方精云, 王襄平, 沈泽昊等. 植物群落清查的主要内容、方法和技术规范. 生物多样性, 2009, 17(6): 533-548.]
[14] Jin Zhenzhou ed. Theories and methods of phytosociology. Beijing: Science Press, 2009: 69-72.[金振洲. 植物社会学理论与方法. 北京: 科学出版社, 2009: 69-72.]
[15] He Zongming, Yang Yusheng, Chen Guangshuietal. Spatial patterns of main species in a 76-year-old Chinese fir community in Ancaoxia, Fujian.JournalofFujianCollegeofForestry, 2003, 21(3): 212-2l5.[何宗明, 杨玉盛, 陈光水等. 安曹下76年生杉木群落主要种群分布格局. 福建韩学院学报, 2003, 21(3): 212-2l5.]
[16] Cao Honglin, Cai Xi’an, Peng Shaolinetal. An analysis of the community of secondary evergreen broad-leaf forestin Longkou, Heshan.TropicalGeography, 1999, 19(4): 312-317.[曹洪麟, 蔡锡安, 彭少麟等. 鹤山龙口村边次生常绿阔叶林群落分析. 热带地理, 1999, 19(4): 312-317.]
[17] Zhang Yashuang, Su Zhixian, Hu Jinyaoetal. Spatial distribution pattern ofDavidiainvolucratepolulation in Wolong of Sihcuan Province.ActaBotanicaYunnanica, 2005, 27(4): 395-402.[张亚爽, 苏智先, 胡进耀等. 四川卧龙自然保护区珙桐种群的空间分布格局. 云南植物研究, 2005, 27(4): 395-402.]
[18] Peng Shaolin, Fang Wei. Studies on dynamics ofCastanposischinensisandSchimasuperbapopulation in forest succession of Dinghushan mountain.ActaPhytoecologicaSinica, 1995, 19(4): 311-318.[彭少麟, 方炜. 鼎湖山植被演替中椎栗和荷木种群的动态.植物生态学报, 1995, 19(4): 311-318.]
[19] Zhang Chao, Huang Qinglin, Pubu Dunzhuetal. Population distribution patterns of shrub in Tibet.JournalofMountainScience, 2011, 29(1): 123-128.[张超, 黄清麟, 普布顿珠等. 西藏灌木林种群分布格局. 山地学报, 2011, 29(1): 123-128.]
[20] Cui Baoshan, Yang Zhifeng eds. Wetlands. Beijing: China Normal University Press, 2006: 54-55.[崔保山, 杨志峰. 湿地学. 北京: 北京师范大学出版社, 2006: 54-55.]
[21] Zhang Baoyuan. Consideration on Jianhu lake conservation in plateau area.EnvironmentalScienceSurvey, 2011, 30(3): 49-52. [张宝元. 对保护高原湖泊剑湖的思考. 环境科学导刊, 2011, 30(3): 49-52.]
[22] Li Weidong, Liu Yungen, Tian Kunetal. Research on reduction efficiency for in let lake sandy sludge by plateau JianhuZizanialatifoliawetland lakeside zone of Yunnan north-west region.JournalofAnhuiAgriculturalSciences, 2010, 38(31): 17643-17645,17685. [李卫东, 刘云根, 田昆等. 滇西北高原剑湖茭草湿地湖滨带对入湖泥沙截留效果研究. 安徽农业科学, 2010, 38(31): 17643-17645, 17685.]
[23] Li WD, Liu YG, Tian Ketal. Research on purification effect of Jian LakeZizanialatifoliawetland lakefront zone in northwest of Yunnan Plateau on nitrogen and phosphorus of agricultural non-point source.AgriculturalScienceandTechnology, 2010, 11(7): 109-112, 150.
[24] Wu Tonggui, Wu Ming, Xiao Jianghua. Dynamics of community succession and species of vegetations in beach wetland of Hangzhou Bay.ChineseJournalofEcology, 2008, 27(8): 1284-1289.[吴统贵, 吴明, 萧江华. 杭州湾滩涂湿地植被群落演替与物种多样性动态. 生态学杂志, 2008, 27(8): 1284-1289.]
[25] Hou Yuan, Guo Zhenggang, Long Ruijun. Changes of plant community structure and species diversity in degradation process of Shouqu wetland of Yellow River.ChineseJournalofAppliedEcology, 2009, 20(1): 27-32.[后源, 郭正刚, 龙瑞军. 黄河首曲湿地退化过程中植物群落组分及物种多样性的变化. 应用生态学报, 2009, 20(1): 27-32.]
[26] Lou Yanjing, Zhao Kuiyi, Ma Keping. Change in floristic composition and species diversity of plant community along environment gradient in Honghe National Nature Reserve, China.ActaEcologicaSinica, 2007, 27(9): 3883-3891. [娄彦景, 赵魁义, 马克平. 洪河自然保护区典型湿地植物群落组成及物种多样性梯度变化. 生态学报, 2007, 27(9): 3883-3891.]
[27] Xiao Keyan, Yu Dan. Concepts and studies on clonal population of aquatic plant.ActaHydrobiologicaSinica, 2008, 32(6): 920-925. [肖克炎, 于丹. 克隆种群的有关概念在水生植物中应用和研究进展. 水生生物学报, 2008, 32(6): 920-925.]
[28] Zhu Guicai, Zhang Xiangyin, Yang Zhongyi. Study on reproductive characteristics of co-tolerance plantLeersiahexandra.ShandonAgriculturalScience, 2009, 9: 26-29.[朱桂才, 张饷银, 杨中艺. 共耐性植物李氏禾繁殖特性的研究. 山东农业科学, 2009, 9: 26-29.]
[29] Zhang Jiyi, Zhao Halin. Spatial patterns of main species of the grassland community in the recovering succession in Horqin sandy land.ChineseJournalofEcology, 2004, 23(2): 1-6. [张继义, 赵哈林. 科尔沁沙地草地植被恢复演替进程中群落优势种群空间分布格局研究. 生态学杂志, 2004, 23(2): 1-6.]
[30] Zhao Kuiyi, Sun Guangyou, Yang Yongxing eds. Mires of China. Beijing: Science Press, 1999: 31-32.[赵魁义, 孙广友, 杨永兴. 中国沼泽志. 北京: 科学出版社, 1999: 31-32.]
Species composition and distribution patterns ofOtteliaacuminatacommunity in Lake Jianhu, northwestern Yunnan
LI Ningyun1,2, CHEN Yuhui3, HU Jinming1, LIU Zhaopeng4, LI Jing3, AO Xinyu3& LEI Ran3
(1:YunnanUniversity,Kunming650091,P.R.China) (2:YunnanAcademyofForestry,Kunming650201,P.R.China) (3:SouthwestForestryUniversity,Kunming650224,P.R.China) (4:NationalPlateauWetlandResearchCenter,Kunming650224,P.R.China)
Otteliaacuminatais listed in Yunnan Province Protected Plants (Class III) as a rare and endangered plant species.Otteliaacuminatacommunity used to be a dominant community type in Yunnan’s plateau lakes, but it is only found in a limited number of lakes nowadays with its observable changing characteristics. In the present study, species composition and distribution patterns of the community in Lake Jianhu were studied, with plot method and dispersal coefficient method in order to understand the development trend of the population, the relationship between its distribution pattern and environmental factors, and to provide some scientific basis for the protection ofOtteliaacuminataand plateau lakes. The results showed that the community was composed of 18 species belonging to 12 families and 14 genera, with submerged type, free-floating type, floating-leaved type and hygrophyte as four major life forms. The distribution pattern ofOtteliaacuminata,Nasturtiumofficinale,PotamogetonmalaianusandNajasgramineawas clumpy, while other species were randomly distributed. “Crowding effect”inOtteliaacuminatapopulation was the highest among all species,which indicated the intense intraspecific competition.Ex-situconservation, basin maintenance and habitat management are some promising approaches to effectively slow down lake swamping process and conserveOtteliaacuminatacommunity.
Species composition; population distribution pattern;Otteliaacuminatacommunity; lake swamping; Lake Jianhu
*云南省应用基础研究计划面上项目(2011FB130)和国家重点基础研究发展计划“973”项目(2010CB434807)联合资助. 2016-03-07 收稿;2016-07-22收修改稿. 李宁云(1977~),男,博士研究生;E-mail: ningyun.li@163.com.
