鳙幼鱼在下行过程中应对加速流的行为响应研究
2017-05-16林晨宇刘德富许家炜石小涛
林晨宇 罗 佳 刘德富 吴 欢 许家炜 石小涛
(三峡大学, 三峡库区生态环境教育部工程研究中心, 宜昌 443002)
鳙幼鱼在下行过程中应对加速流的行为响应研究
林晨宇 罗 佳 刘德富 吴 欢 许家炜 石小涛
(三峡大学, 三峡库区生态环境教育部工程研究中心, 宜昌 443002)
以具有下行洄游需求的鳙(Aristichthys noblis)幼鱼为研究对象, 在自行设计制造加速流的装置中进行下行实验, 结果发现: 在实验设置的3个入口流速下(入口流速分别为: 0.018、0.034和0.053 m/s), 鳙幼鱼的通过率均为100%。在下行通过孔口的方式上, 以尾部先通过为主(即顶流通过), 尤其是在流速较高的时候, 头部先通过比例仅为6.25%。在通过总时间(从实验开始至通过孔口的时间)上, 鳙幼鱼在低流速条件下显著小于在中流速条件下(P<0.05), 而在高流速条件下与中低两个流速条件下均无显著性差异(P>0.1)。在通过时间(从实验鱼最后一次进入加速区域, 至通过孔口的时间)上, 鳙幼鱼在三种流速条件下没有显著差异性(P>0.1)。研究表明, 加速流会对鳙幼鱼下行的时间和方式产生影响。研究旨在为鱼类资源保护, 帮助鱼类安全高效过坝提供基础数据, 同时也为研究鱼类在下行过程中应对加速流的行为研究提供一种研究方法。
鳙幼鱼; 下行; 加速流; 洄游
鱼类在不同水域进行周期性迁徙的行为, 称之为洄游[1], 常见的洄游类型根据其洄游目的不同可以分为: 生殖洄游、索饵洄游和越冬洄游等。大坝的修建在阻断河流连续性的同时也阻断了鱼类的洄游通道[2—4]。无论是溯河洄游还是降河洄游, 鱼类在从大坝的一端向另一端迁徙时, 会因大坝的阻隔而难以顺利完成迁徙过程[5—7]。其中, 降河洄游的鱼类在下行通过大坝及其他水工建筑物时, 除了要面对不同程度的水流落差外, 还需要克服各下行通道产生的各典型特征流场[8,9]。而水流速度逐渐增加的加速流作为一种鱼类下行过程中必须面临的特征水流, 在各通道中普遍存在, 包括水轮机、溢洪道、船闸及过鱼旁路等[10]。加速流可能引起鱼类下行过坝过程中的逃逸行为或造成鱼类因被动卷吸而受到伤害[11]。因此研究鱼类下行过程中应对加速流的响应行为, 可以为设计帮助鱼类安全高效过坝的辅助设施等方面提供理论依据。虽然过鱼设施的修建, 能够在一定程度上缓解大坝修建给鱼类等水生生物带来的不利影响[5—7], 但是我国的鱼类保护研究仍然相对落后, 存在诸多的科学问题和实践困难, 如鱼类分布对环境改变的适应策略、鱼类行为对水力特征的响应关系和辅助鱼类通过闸坝的救鱼技术等[12]。
目前有关鱼类行为与水力因子响应关系的研究主要集中在辅助鱼类上溯的过鱼设施领域[13—15]。如有学者通过模型结合数值模拟的方法(Fluent, Flow3D以及Mike等软件)获取鱼道内的流速分布、自由水面特征、紊动能、雷诺应力以及射流分布形态等, 以优化鱼道设计或发现鱼类的上溯行为规律[16]。而在鱼类下行方面的研究相对较少, 仅局限于少量行为学指标的分析[2]。鳙(Aristichthysnobilis)与青鱼(Mylopharyngodon piceus)、草鱼(Ctenopharyngodon idellus)、鲢(Hypophthalmichthys molitrix)合称为“四大家鱼”在我国各大水域广泛分布,是重要的经济鱼类之一, 其仔鱼具有下行洄游的需求, 本文基于鳙幼鱼下行通过加速流的行为研究,了解鳙幼鱼下行过程中应对加速流的行为响应特征, 有助于鱼类顺利下行设施的优化设计。同时,本研究还提供了一种研究鱼类下行过坝的思路, 为鱼类安全下行过坝研究提供参考。
1 材料与方法
1.1 实验材料
实验用鱼由湖北宜都渔场提供, 体重(以湿重表示)(13.44±2.55) g, 体长(10.14±0.75) cm。实验用鱼于2015年4月通过活鱼充氧袋运送至三峡大学生态水工实验室, 暂养7d后进行实验。暂养水槽为直径2 m、深0.5 m的圆形玻璃纤维水缸。暂养期间水温为(20.00±1.00)℃, 24h连续持续充氧。
根据行为学实验要求, 同一实验鱼不再重复实验, 避免对实验结果造成影响。
实验设备主要包括鳙幼鱼的下行实验装置(图 1)和视频采集装置(索尼DSC-W360)。下行装置为长方形水槽(300 cm×100 cm×50 cm), 材质为钢化玻璃。实验区域上游部分为长方形, 下游部分为收缩状, 用于制造水流速度梯度。水槽通过调节电机频率来控制水流速度, 通过整流栅(C)整流后得到均匀流场。整个实验装置为开放式水槽, 溶解氧(6.5±0.2) mg/L。装置上方距离地面6 m处架设摄像头, 用于记录实验过程。
1.2 实验方法
实验采用曝气48h的自来水作为实验用水, 水槽中水温在实验前调节至与暂养水槽中水温一致。实验选择健康活泼的鳙幼鱼, 在三组不同的入口水流速度(指在D区域上游的水流速度)下进行, 水流速度分别为0.018、0.034和0.053 m/s, 流速选择为鳙在自由游泳状态下的游泳速度, 0.018 m/s为鳙在流水条件下的不活跃游泳速度, 逐渐增加流速, 但是流速不超过鳙在流水条件下的活跃游泳速度[17]。每一次实验随机选取1尾鱼, 置于D区域上游部分,适应1.5h后, 打开拦网(D区, 距离C部分5 cm处)同时开启摄像头。若2h内实验鱼通过水槽下游的加速流区域(E区), 则实验结束, 否则实验在2h(长时间不通过加速流区域不利于鱼类的下行, 因此设置2h不通过则结束实验, 并视为不能通过加速流区域)后结束, 且视为实验鱼不通过加速流区域。整个实验过程采用摄像头录像。同一实验鱼不重复使用, 每一个流速下的实验重复18—22次, 3组实验共使用60尾鱼。
1.3 数据处理
本实验根据摄像头录像, 通过视频处理获得相关数据包括头部通过比例、通过总时间、通过时间等。其中, 通过总时间的定义为: 从实验开始至通过孔口的时间; 通过时间的定义为: 从实验鱼最后一次进入加速区域, 至通过孔口的时间。
采用视频分析软件Logger Pro对视频进行分析获取相关数据。所得数据首先采用Excel计算处理以及绘图, 之后采用SPSS19.0综合数据分析。统计值(主要包括通过时间和总时间在各个不同流速下的差异性)使用平均值±标准差(Mean±SE)表示, 用单因素方差分析法分析差异性, P<0.05表示差异显著。
2 结果
在实验设置的3种水流速度下, 鳙的通过率均为100%。鳙在通过孔口的方式上基本相同, 主要以尾部通过为主, 尤其是在水流速度较高的时候,头部通过的比例最低, 仅6.25%。在低流速和中流速条件下, 头部通过比例相对高流速较大, 分别为31.82%和31.25%(图 2)。
鳙在三种不同的水流速度下, 从实验开始至通过孔口的总时间存在一定的差异性(图 3)。其中鳙在低流速下的总时间与鳙在中流速下的总时间存在显著性差异(P<0.05), 低流速下的总时间明显小于中流速下的时间, 但是在高流速下的总时间与前两种水流条件下的总时间不存在差异性(P>0.1)。

图 1 鳙幼鱼下行实验装置示意图Fig. 1 Equipment for the simulation of fish going downstreamA. 储水区; B. 整流区; C. 整流栅; D均匀流区域; E. 加速流区域; F. 尾水区域

图 2 鳙幼鱼在3种水流条件下头部通过孔口的比例Fig. 2 The proportion of head-first in three different flow condition
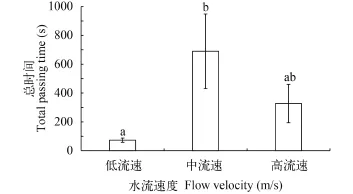
图 3 鳙在3种不同流速下下行的总时间Fig. 3 The total passing time of fish in three different flow velocity
本研究还对鳙在下行过程中最后一次进入加速区域至通过孔口的时间进行了统计(图 4), 结果显示鳙在3种不同的水流条件下, 通过加速区域的时间没有差异性(P>0.1)。
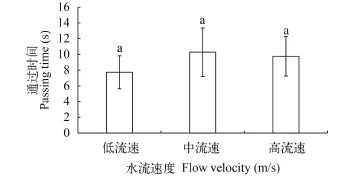
图 4 鳙在3种不同水流速度下下行通过加速流的通过时间Fig. 4 Time for fish passing the accelerating flow in three different flow velocity
3 讨论
不同鱼类在下行通过加速区域时, 其通过方式存在一定的差异性。例如鲑(Oncorhynchus)幼鱼在主动洄游时, 通常会以头部先通过加速区域, 而太平洋鲑(Oncorhynchus spp .)幼鱼通常会先面向下游靠近加速流, 然后在特定的点改变方向[18]。在本研究中, 鳙在3种设定流速下, 以尾部通过为主, 尤其是在水流速度较大的时候, 头部通过比例最低, 同时本研究也发现了转弯现象, 但是发生转弯现象的区域并没有显示明显的规律性。这也说明了不同鱼类在下行过程中会采取不同的行为策略。
水流速度会影响鱼类的游泳行为[19,20]。鱼类对水流具有一定的选择性, 可以主动对抗水流而逆流游泳, 同时可以通过调节自身游泳速度来适应水流速度[20]。钟金鑫等[19]对白鱼(Anabarilius grahami)游泳行为的研究发现, 在有流速的时候, 趋流率高于静水组, 但是不同的流速组之间没有差别。本研究发现鳙下行的总时间在低流速和中流速之间存在显著的差异性, 高流速与另外两种流速之间不存在显著差异性; 而通过时间在3种水流条件下均不存在差异性。这说明随着水流速度的增加, 鳙顶流时间会随之增长, 而流速继续增加超过特定水平时, 顶流时间不会增加并且会呈现降低的趋势。这与何大仁等[21,22]的研究结果类似, 鱼类会根据水流速度和方向调节自身的游泳速度和方向, 例如保持逆流游泳的状态, 长时间的停留在某一地方, 因此才会引起时间上的差异性。
本研究还发现, 在设置的3种水流条件下, 鳙的通过率均达到了100%。其中一种可能性是由于在较高的水流速度下, 鳙通无法抵抗水流速度而顺水下行, 而在较低的水流速度下加速流对鳙有一定的吸引能力, 使鳙自行下行; 另外一种可能是本装置中设计的3种水流梯度不够大, 还未能达到使鳙逃离加速区域并拒绝下行的程度。因此后续应该改变加速区域的变化梯度, 进一步探讨鳙下行过程中应对加速流的行为。研究鳙幼鱼在不同水流速度下应对加速流的行为特征, 可以帮助优化设计鱼类下行过鱼设施, 同时也可以作为研究其他重要鱼类的参考。
[1]Cooke J S, Hinch G S, Donaldson R M, et al. Conservation physiology in practice: how physiological knowledge has improved our ability to sustainably manage Pacific salmon during up river migration [J]. Philosophical Transactions of the Royal Society of London Series B, 2012, 367(1607): 1757—1769
[2]Zhu H F, Liu L, Lu B, et al. A review of fish passage facilities for downstream migrants [J]. Journal of Yangtze River Scientific Research Institute, 2015, 32(10): 33—37 [朱海峰, 刘流, 路波, 等. 鱼类下行过坝相关工程措施综述. 长江科学院院报, 2015, 32(10): 33—37]
[3]Gao Y, Chang J B. Population viability analysis on Chinese Sturgeon [C]. Ichthyology Branch of China Marine Lakes and Marshes Institute & Ichthyology Branch of China Institute of Zoology. 2004, 43—44 [高勇, 常剑波. 过鱼设施对鱼类资源保护的意义. 中国海洋湖沼学会鱼类学分会、中国动物学会鱼类学分会2004年学术研讨会摘要汇编. 2004, 43—44]
[4]Bednarek A T, Hart D D. Modifying dam operations to Restore River: Ecological Responses to Tennessee River Dam mitigation [J]. Ecological Applications, 2005, 15(3): 997—1008
[5]Liang Y Y, Liu D F, Shi X T, et al. Overview of fish barging [J]. Journal of Yangtze River Scientific Research Institute, 2014, 43(4): 25—29, 34 [梁园园, 刘德富, 石小涛, 等. 集运鱼船研究综述. 长江科学院院报, 2014, 31(2): 25—29, 34]
[6]Luo J, Jiang W, Chen Q W, et al. Estimation on abundance of egg-predatory fishes in the spawning ground of Chinese sturgeon below the Gezhouba Dam [J]. Freshwater Fisheries, 2013, 43(4): 27—30 [罗佳, 姜伟, 陈求稳,等. 葛洲坝下中华鲟产卵场食卵鱼资源量的调查和分析. 淡水渔业, 2013, 43(4): 27—30]
[7]Xiao H. Gezhouba Water condervancy hub and the protection practice of rere fish [A]. In: Huang Z L, Fu B J, Yang Z F (Eds.), The Yangtze River Ecological and Environmental Protection in Large Water Conservancy Project in 21st Century [M]. Beijing: China Environmental Science Press. 1998, 199—209 [肖慧. 葛洲坝水利枢纽与珍稀鱼类保护实践. 黄真理, 付博杰, 杨志峰. 21世纪长江大型水利工程中的生态与环境保护. 北京: 中国环境科学出版社. 1998, 199—209]
[8]Piper A T, Wright R M, Walker A M, et al. Escapement, route choice, barrier passage and entrainment of seaward migrating European eel, Anguilla anguilla, within a highly regulated lowland river [J]. Ecological Engineering, 2014, 57: 88—96
[9]Haro A, Odeh M, Noreika J, et al. Effect of water acceleration on downstream migratory behavior and passage of Atlantic salmon smolts and juvenile American shad at surface bypasses [J]. Transactions of the American Fisheries Society, 1998, 127(1): 118—127
[10]Venditti D A, Rondorf D W, Kraut J M. Migratory behavior and forebay delay of radio-tagged juvenile fall Chinook Salmon in a lower snake river impoundment [J]. North American Journal of Fisheries Management, 2000, 20(1): 41—52
[11]Boysen K A, Hoover J J. Swimming performance of juvenile white sturgeon (Acipenser transmontanus): training and the probability of entrainment due to dredging [J]. Journal of Applied Ichthyology, 2009, 25(2): 54—59
[12]Heisey P G, Mathur D, Fulmer J L, et al. Turbine passage survival of late-running adult American shad and its potential effect on population restoration [C]. Amaral S V, Mathur D, Taft III E P. American Fisheries Society Symposium. 2008, 61: 141—152
[13]Sun X L, Zhao Y, Tian Z L. Latest international development of anadromous fish passage facilities at hydropower stations [J]. Water Resources and Hydropower Engineering, 2009, 40(12): 133—136 [孙小利, 赵云, 田忠禄. 国外水电站的洄游鱼类过坝设施最新发展. 水利水电技术, 2009, 40(12): 133—136]
[14]Zhou P, Zhou Y T, Yao B S. Water resources and hydropower development in the research status and development trend of the fishway [J]. Water Conservancy Construction & Management, 2011, 31(7): 40—43 [周鹏, 周殷婷, 姚帮松. 水利水电开发中鱼道的研究现状与发展趋势. 水利建设与管理, 2011, 31(7): 40—43]
[15]Bell M C. Fisheries handbook of engineering requirements and biological Criteria [R]. Fisheries Engineering Research Program, North Pacific Division, Corps of Engineers, Portland, Dregon. 1973
[16]Qi L, Yang Y, Wang Y, et al. Fish behavior characteristics in response to change of hydrodynamic environment [J]. Journal of Hohai University (Natural Sciences), 2012, 40(4): 438—445 [齐亮, 杨宇, 王悦, 等. 鱼类对水动力环境变化的行为响应特征. 河海大学学报(自然科学版), 2012, 40(4): 438—445]
[17]Luo J, He L, Lin C Y, et al. Comparative study on circadian rhythm of swimming of Aristichthys nobilis juveniles in still and flowing water [J]. Acta Hydrobiologica Sinica, 2016, 40(1): 194—197 [罗佳, 何立, 林晨宇,等.鳙幼鱼在静水和流水条件下昼夜游泳节律初步研究. 水生生物学报, 2016, 40(1): 194—197]
[18]Kroese A B A, Schellart N A M. Velocity and acceleration-sensitive units in the trunk lateral line of the trout [J]. Journal of Neurophysiology, 1992, 68(6): 2212—2221
[19]Zhong J X, Zhang Q, Li X R, et al. Effects of water velocity on the swimming behavior of Anabarilius graham [J]. Chinese Journal of Ecology, 2013, 32(3): 655—660 [钟金鑫, 张倩, 李小荣, 等. 不同流速对白鱼游泳行为的影响. 生态学杂志, 2013, 32(3): 655—660]
[20]Li D, Lin X T, Zhu Z M, et al. Effects of flow rate on swimming states and activity metabolism in juvenile Hybrid Sturegeon [J]. Acta Hydrobiologica Sinica, 2011, 35(4): 578—585 [李丹, 林小涛, 朱志明, 等. 不同流速下杂交鲟幼鱼游泳状态与活动代谢研究. 水生生物学报, 2011, 35(4): 578—585]
[21]He D R. The research of Russian fish behavior and feeling (I) [J]. Taiwan Strait, 1996, 15(2): 191—199 [何大仁.俄国鱼类行为与感觉研究(I). 台湾海峡, 1996, 15(2): 191—199]
[22]He D R. Fish Behavior [M]. Xiamen: Xiamen University Press. 1998, 141—145 [何大仁. 鱼类行为学. 厦门: 厦门大学出版社. 1998, 141—145]
THE BEHAVIOR RESPONSE OF JUVENILE ARISTICHTHYS NOBILIS TO THE ACCELERATING FLOW DURING THEIR DOWNSTREAM MIGRATION
LIN Chen-Yu, LUO Jia, LIU De-Fu, WU Huan, XU Jia-Wei and SHI Xiao-Tao
(Engineering Research Center of Eco-environment in Three Gorges Reservoir Region, China Three Gorges University, Yichang 443002, China)
To investigate the ability of downstream migration of Juvenile bighead carp (Aristichthys noblis), a self-designed equipment with three inlet velocity (0.018m/s, 0.034m/s, 0.053m/s) was utilized. Results indicated that the passing rate under three inlet velocity were all 100%. Most fish used tail through first (tail-first) to pass the hole, especially under the high velocity, and the percentage of head-first was merely 6.25%. The total passing time in the low velocity was significantly less than the middle velocity (P<0.05), and no difference existed between the high velocity and the other two conditions (P>0.10). In regard to the passing time, from the time subject enter the accelerating flow field to the moment it left the experimental area, the carp appeared no significant difference (P>0.10) in all three conditions. These results indicated the impact the accelerating flow on the duration and methods of Juvenile bighead carp’s downstream action, which provide knowledge for fish passage techniques and preserve fish resources. In addition, this work established an available method to study the behavioral reaction fish fight accelerating flow during its downstream migration.
Juvenile Aristichthys nobilis; Downstream; Accelerating flow; Migration
Q178.1
A
1000-3207(2017)03-0560-05
10.7541/2017.72
2016-07-19;
2016-12-24
国家自然科学基金(51579136); 技术服务项目(SDHZ2015158)资助 [Supported by the National Natural Science Foundation of China (51579136); Technical Service Project (SDHZ2015158)]
林晨宇(1991—), 男, 福建福安人; 硕士; 主要从事生态水利学研究。E-mail: 529663732@qq.com
石小涛(1981—), 男, 湖北红安人; 博士; 主要从事水生动物生态学研究。E-mail: sxtshanghai@163.com
