金门岛北部海域浮游植物的季节变动及与环境的关联
2017-05-16叶又茵林更铭杨清良
王 雨 项 鹏 叶又茵 林更铭 杨清良 林 茂
(国家海洋局第三海洋研究所, 厦门 361005)
金门岛北部海域浮游植物的季节变动及与环境的关联
王 雨 项 鹏 叶又茵 林更铭 杨清良 林 茂
(国家海洋局第三海洋研究所, 厦门 361005)
采用2013—2014年四季度月在金门岛北部海域获取的浮游植物及环境因子监测数据, 分析该区浮游植物的群落结构和季节变化及其与温度、盐度、悬浮物、营养盐、叶绿素等的关系, 初步探讨涉海工程建设对浮游植物群落的潜在影响。结果显示, 鉴定出的浮游植物隶属3门43属82种(不含未定种), 群落构成以硅藻为主, 其次是甲藻, 蓝藻仅1种。物种组成的季节差异较大, 3月物种贫乏, 1月次之, 7月和11月最丰富。四季丰度平均为47.09×103cells/L, 1月丰度最高, 7月次之, 11月最低, 3月高于11月少许。四季优势种均为硅藻, 13个优势种分别为柔弱几内亚藻(Guinardia delicatula)、短角弯角藻(Ecampia zoodicaus)、骨条藻(Skeletonema spp.)、具槽帕拉藻(Paralia sulcata)、微小海链藻(Thalassiosira exigua)、标志星杆藻(Asterionella notula)、旋链角毛藻(Chaetoceros curvisetus)、新月菱形藻(Nitzchia closterium)、派格棍形藻(Bacillaria paxillifera)、异常角毛藻(Chaetoceros abmormis)、小细柱藻(Leptocylindrus minutum)、宽角曲舟藻(Pleurosigma angulatum)和美丽曲舟藻(Pleurosigma formosum)。不同季节优势种有一定程度交错, 仅在单季占优的有6种, 有2/3在3个以上季节出现, 具槽帕拉藻、骨条藻为四季优势种。浮游植物物种多样性和均匀度总体较好, 群落结构稳定。与毗邻海区相比, 本区物种丰富度偏低, 丰度高于毗邻海区, 种类组成相似, 优势种却有较大差别。Pearson相关分析表明, 溶解无机氮及活性磷酸盐仅在1月与丰度存在极显著的正相关, 是促使丰度为四季最高的原因。涉海工程施工产生的悬浮物和冲击波是影响浮游植物群落的主要因素, 大量海洋工程建设案例表明, 施工期造成的浮游植物丰度下降趋势和优势种更替混乱在工程结束后能得以恢复。
浮游植物; 群落结构; 季节变动; 环境因子; 相关分析; 金门岛北部海域
金门岛地处福建南部九龙江口的厦门岛以东,其北部海域水动力条件复杂, 除潮流、海浪外, 还有地表径流参与作用, 尤其金门北东水道流速最大值可达到2.2 m/s, 潮流能蕴藏量巨大[1]。近期, 厦门机场二期工程在金门北部的大嶝岛开始基础性建设, 工程填海面积相当于10个鼓浪屿。随着工程全面展开, 大嶝岛与小嶝岛岛区的陆域作业(吹填)、航道疏浚等施工将造成悬浮物的大量增加进而影响海洋生态[2—4], 而港口爆破(炸礁、爆破挤淤)产生的冲击波对海洋生物也易造成影响[5]。厦门机场二期工程建设有可能使金门北部海域环境发生较大的变化, 对海洋生态与环境产生不同程度的影响。
浮游植物群落结构与初级生产力沿食物链向上传递转化为渔业资源产量的效率紧密相关, 它的种群动态和丰度变动通常是栖息环境的直接综合效应, 与环境质量密切联系[6]。浮游植物群落结构特征也是评价水体水质状况及其变化趋势的重要指标[7]。有关厦门海区浮游植物, 不少学者已作过较多的分类和生态的研究, 也多次对一些赤潮事件进行了跟综调查[8—14], 调查范围主要在厦门西港[8,9,12]、九龙江河口区[13,14]和厦门东部海域[10,11]。金门岛北部海域至今少见专门的研究[13], 特别是采水样品的浮游植物定量分析结果未见报道, 并且已有的相关文献多是20世纪90年代的研究结果, 有必要为该海域浮游植物的资料进行补充完善, 进而深入了解金门北部海域的浮游植物群落的生态现状及变动情况。
为研究金门北部海域浮游植物群落季节变动及其与环境因子关系, 并确定浮游植物物种多样性水平, 本文根据2013—2014年四个季度月在金门岛北部海域获取的海洋生态调查数据与资料, 分析该海域浮游植物的群落结构、丰度分布及季节变动、优势种组成及物种多样性, 讨论浮游植物与温度、盐度、悬浮物、营养盐、叶绿素的关系, 与毗邻海区比对分析, 并初步探讨涉海工程的施工建设对浮游植物群落的潜在影响, 为厦门机场二期工程完成后的生态影响评估提供本底资料。
1 材料与方法
1.1 调查站位和环境参数测定
浮游植物样本采集参照《海洋调查规范》(GB/T-2007)[15]进行。分别于2013年11月(秋)、2014年1月(冬)、2014年3月(春)和2014年7月(夏)共四季度的月在金门北部海域(118.05°—118.55°E, 24.33°—24.65°N)布设15个浮游植物采样站位 (图1)。同期进行水文和化学调查。温度、盐度等环境参数用美国维赛YSI多参数水质分析仪(Professional Plus 605)现场测定。悬浮物、叶绿素a、营养盐取样分析, 参照《海洋调查规范》(GB/T-2007)[15]进行测定。
1.2 浮游植物取样和分析方法
在上述站位用Niskin采水器分表层(离水面0.5 m)和底层(离海底1 m)各取500 mL水样, 样品用3%中性甲醛固定。水深低于5 m的站位仅采集表层水样, 缺失的底层数据在统计分析中不计入统计变量[15]。实验室内分析样品经沉降浓缩至一定体积后, 使用Kolkwitz计数框在Zeiss Z1倒置光学显微镜下进行鉴定和计数, 并按种类计算丰度(cells/L)。浮游植物种名更改参照孙军的研究[16]。需要注意的是, 骨条藻属下涵盖多个物种, 中肋骨条藻(Skeletonema costatum)一直作为骨条藻的模式种。近年来借助分子生物学和电子显微镜技术, 发现骨条藻属共计有13个今生种[17]。由于骨条藻形态在光学显微镜下的不易判断, 在以往的研究中, 我国学者则普遍将分布于我国近海的骨条藻种类都定名为同一个种: 中肋骨条藻[17,18]。中肋骨条藻虽然分布范围广泛, 但其不同季节的丰度不同, 以往有关中肋骨条藻的各种报道其研究对象未必是中肋骨条藻。鉴于此, 本文基于光学镜检以骨条藻作为物种鉴定结果。
物种多样性指数(H′)、均匀度(J)、优势度(Y)分别采用以下计算公式[19]:
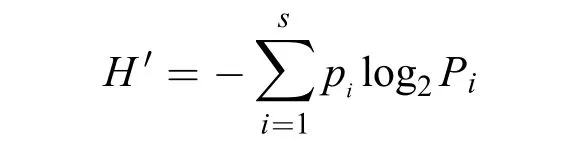
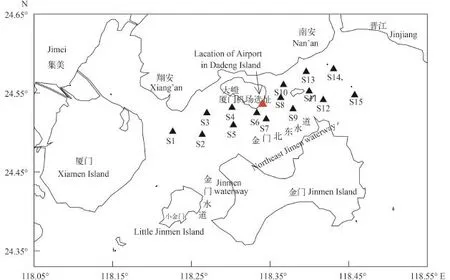
图 1 金门岛北部海域的浮游植物采样站位Fig. 1 Sampling stations of phytoplankton in sea waters of the Northern Jinmen Island
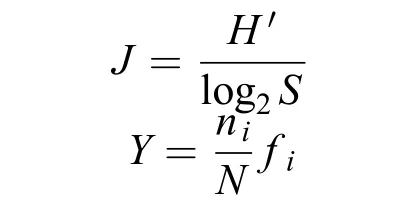
式中S为样品中的种类数, Pi为第i种的细胞丰度与样品总丰度的比值, ni为第i种的细胞丰度, N为每种出现的细胞丰度的总和, fi为第i种的测站出现率。
浮游植物丰度的平面分布采用Surfer8.0软件绘制, 多样性、均匀度指数采用Primer5.0进行数据分析, 与环境因子的关联采用SPSS11.5软件分析。
2 结果
2.1 种类组成与季节变化
金门北部海域2013—2014年四季度所获样品95份, 共鉴定浮游植物82种(不含未定种), 分属于硅藻、甲藻、蓝藻3个门类(表 1)。硅藻70种, 占所有物种的85.4%; 甲藻11种, 占所有物种的13.4%; 蓝藻1种, 占1.2%。种类组成以广温种居多, 占所有物种的77.3%, 暖水种占有率为15.8%, 此外还有少数温带种。物种的生态类型体现较明显的亚热带区系特征, 这与毗邻的厦门东海域[10,11]、厦门西港[8,9,12,13]浮游植物群落的生态特征一致。硅藻为浮游植物的主体, 对浮游植物种类组成与丰度变化均起决定作用, 成为浮游植物优势种最主要的构成者, 以圆筛藻(Coscinodiscus)、根管藻(Rhizosolenia)、菱形藻(Nitzschia)为主要类群, 分别为10种、5种和5种;几内亚藻(Guinardia)、曲舟藻(Pleurosigma)、角毛藻(Chaetoceros)也检出较多种类。甲藻以原多甲藻(Protoperidinuim)种类较丰富, 角藻(Ceratium), 鳍藻(Dinophysis)也常见。
11月检出种类最多为61种, 7月次之为55种, 3月最低为42种, 1月有46种, 这有别于1990年厦门东部及大嶝岛近海夏季(9月)物种最多, 而秋季(11月)物种较少的季节变化特点[11,13]。本区总物种数目的四季差异与硅藻种类数目的季节变化趋势一致(表 1)。四季种类组成均以硅藻种类最多, 其次为甲藻、蓝藻。蓝藻类的红海束毛藻 (Trichodesmium erythraeum)在三季均有出现。四季共有物种数目为35种, 占所有物种的42.7%。四季度仅在个别站位(1—4个站)检出的物种占总物种累计不到14.0%, 而在半数以上站位均检出的物种占73.0%以上, 说明浮游植物物种组成在空间站位分布上并无显著差异。
四季常见的种类是旋链角毛藻(Chaetoceros curvisetus)、辐射圆筛藻(Coscinodiscus radiatus)、星脐圆筛藻(Coscinodiscus asteromphalus)、琼氏圆筛藻(Coscinodiscus jonesianus)、布氏双尾藻(Ditylum brightwellii)、具槽帕拉藻(Paralia sulcata)、短角弯角藻(Ecampia zoodicaus)、微小海链藻(Thalassiosira exigua)、骨条藻(Skeletonema spp.)、菱形海线藻(Thalassionema nitzschioides)和夜光藻(Noctilluca scintillans)等。某些高温高盐种, 例如螺端根管藻(Rhizosolenia cochlea)、哈德半盘藻(Palmeria hardmaniana), 以及一些广温性高盐种,例如透明辐杆藻(Bactreristrum hylianum)、细弱海链藻(Thalassiosira subtilis)、伯氏根管藻(Rhizosolenia bergonii)等, 这些种类多出现在台湾海峡暖流较强的夏季(7—9月), 但少数也能见于秋季(10—11月), 与前期研究[8,9,11,13]一致。
2.2 丰度的分布与变化
丰度的季节变动 本区浮游植物丰度四季均值为47.09×103cells/L, 各大类群丰度及均值的季节差异较大(表 2)。1月丰度最高平均为73.28× 103cells/L, 7月次之平均为41.05×103cells/L, 11月最低平均为34.73×103cells/L, 3月略高于11月少许, 平均为39.29×103cells/L。浮游植物丰度高峰期出现在冬季1月, 这与厦门西港和九龙江河口高值出现在夏季7月[8,12,13]的特点不同, 与厦门东部及东北部同安湾高峰期出现在春季3月[10,11]也不同, 更不同于大嶝岛近海前期调查春季3月高值[13,14]的结果。
浮游植物丰度组成四季终以硅藻占绝对优势,各季度均占99.00%以上(表 2)。硅藻丰度自1月最高, 3月至7月和11月逐渐降低, 到11月最低, 不足1月丰度的1/2。1月、7月硅藻丰度比率相对减小,在7月减少尤为明显, 而适温较高的甲藻丰度在此时节则大幅上升, 最大增幅约3倍。蓝藻丰度比率在出现的3月和11月均无较大差别, 但丰度在3月上升尤为明显。

表 1 金门北部海域浮游植物类群的季节变动Tab. 1 Seasonal variation of major groups of phytoplankton in sea water of the Northern Jinmen Island
丰度的平面分布 本区浮游植物表底两层的丰度均值有较大差别, 丰度分布也全然不同。11月浮游植物总丰度四季最低, 表层丰度平均为35.91×103cells/L, 最高值出现在大嶝岛东部离岸S9站, 为49.90×103cells/L。最低值出现在东部离岸最远的S15站, 仅有13.10×103cells/L。总体大嶝岛东部近岸测站丰度都较大, 均大于33.00× 103cells/L (图 2a)。1月丰度四季最高, 高于11月2倍余, 表层丰度平均为67.73×103cells/L, 出现两个极高值, 临接厦门东海域的S1站高达276.00× 103cells/L, 其毗邻S2站出现次高值, 为121.70×103cells/L, 皆因短角弯角藻、微小海链藻高丰度聚集所致。其余测站均保持在(33.00—61.00)× 103cells/L。总体上大嶝岛东西两侧水域丰度差别不大(图 2b)。3月表层丰度平均为38.84×103cells/L, 最高值出现在大嶝岛东部的S7站, 密度为62.40×103cells/L, 西部S3站出现次高值。总体上大嶝岛东西两侧水域丰度差别不大(图 2c)。7月表层丰度平均为42.84×103cells/L, 大嶝岛东部S10、S12、S13站均出现高值。最低密度出现在大嶝岛西部S3站, 仅有15.40×103cells/L。大嶝岛东部丰度大, 西部小(图 2d)。
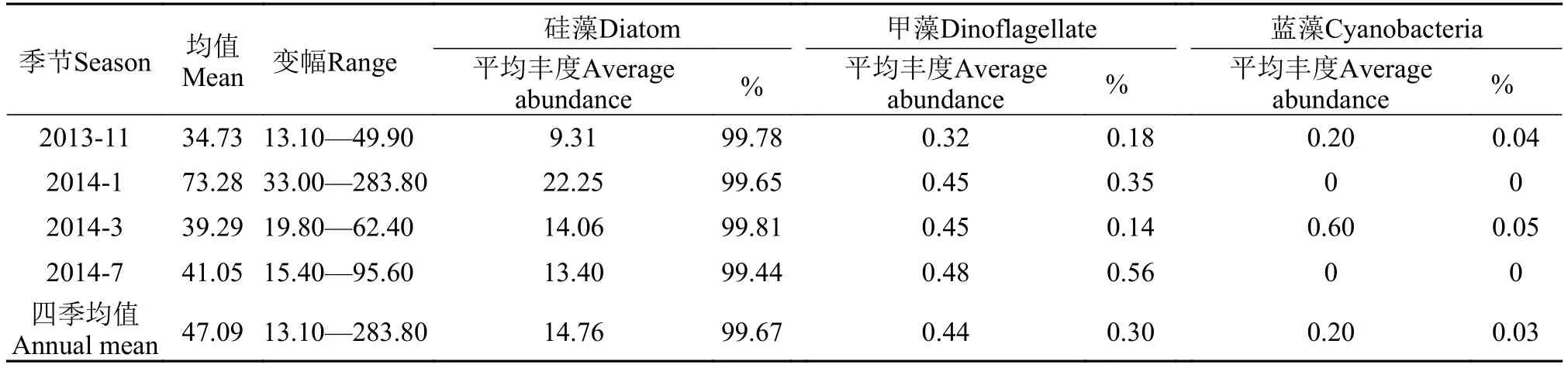
表 2 金门北部海域浮游植物丰度的季节变动(×103cells/L)Tab. 2 Seasonal variation of phytoplankton abundance in sea waters of the Northern Jinmen Island
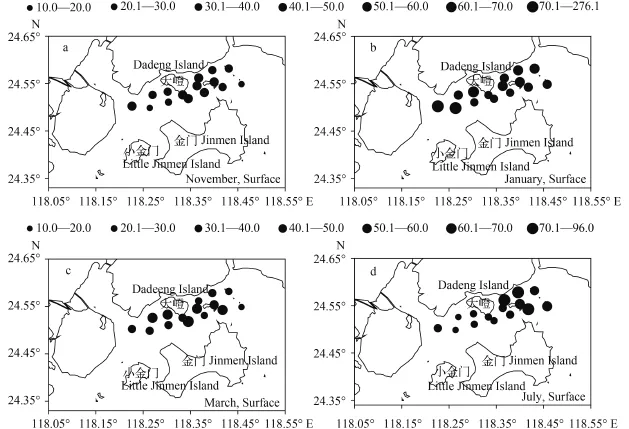
图 2 表层浮游植物丰度的平面分布(×103cells/L)Fig. 2 Horizontal distribution of phytoplankton abundance in surface waters of the Northern Jinmen Islanda. 11月表层Surface in November; b. 1月表层Surface in January; c. 3月表层Surface in March; d. 7月表层Surface in July; 下同The same applies below
因各季节采样现场的实测水深变化, 满足底层采样条件(>5 m)的站位集中在大嶝岛东部, 即是金门北东水道海域。底层丰度分布与表层丰度分布大体相同(图 3)。11月底层丰度平均为33.54× 103cells/L, 略低于表层同期(图 3a)。1月底层丰度高于表层同期, 平均为78.84×103cells/L, 1个极高值同样存在于邻接厦门东海域的S1站(图 3b)。除短角弯角藻、微小海链藻外, 具槽帕拉藻也在此高丰度聚集。3月底层丰度平均为39.74×103cells/L, 略高于表层同期(图 3c)。7月底层丰度平均为39.26×103cells/L, 远低于表层同期(图 3d)。
2.3 优势种与季节变化
以优势度指数Y>0.02为判别标准, 四季优势种均为硅藻, 主要13个优势种有2/3在3个以上季节中出现, 具槽帕拉藻、骨条藻为四季优势种, 仅在单季占优的有6种, 不同季节优势种有一定程度交错,且变化较明显(表 3)。11月优势种多达8种, 柔弱几内亚藻(Guinardia delicatula)、骨条藻和微小海链藻依次位列前三。此外, 派格棍形藻(Bacillaria paxillifera)、新月菱形藻(Nitzchia closterium)、旋链角毛藻和小细柱藻(Leptocylindrus minutum)也占有一定的优势, 派格棍形藻在表层优势度高于骨条藻。具槽帕拉藻在该季优势度不高。柔弱几内亚藻和派格棍形藻仅在11月占优。1月优势种5种, 短角弯角藻在表底的优势度即可达到11月优势种前三者之和, 其仅在该季占优。具槽帕拉藻、骨条藻和微小海链藻优势度均较11月增大, 标志星杆藻(Asterionella notula)此时占有优势种群一席, 其在11月表底的优势度均<0.01。3月优势种减为4种,标志星杆藻优势度最高, 这也是该种在四季的最大优势。具槽帕拉藻和骨条藻优势度有增无减, 标志星杆藻也在3月凸显一定优势。7月优势种最多可达9种, 具槽帕拉藻和骨条藻优势度有降幅, 旋链角毛藻出现四季的最大优势度, 小细柱藻基本与11月优势度持平, 异常角毛藻(Chaetoceros abmormis)、宽角曲舟藻(Pleurosigma angulatum)和美丽曲舟藻(Pleurosigma formosum)仅在7月占优, 而新月菱形藻和微小海链藻仅在表层占优。柳丽华等[20]认为,优势种对群落结构的稳定性有重要影响, 优势种种类数目越多且优势度越小, 则群落结构越稳定, 反之亦然。总体上, 四季优势类群丰度的分布态势决定了浮游植物丰度的分布状况, 不再赘述。
同时, 观测及分析发现, 具槽帕拉藻、骨条藻、旋链角毛藻、小细柱藻、宽角曲舟藻、美丽曲舟藻的优势度在水体表层通常都小于底层(表 3),究其原因, 藻类高丰度沉降或密集在底部, 抑或喜底栖生活, 抑或海区风浪小对底层水体的扰动以及垂直混合影响度小等均能造成该现象。
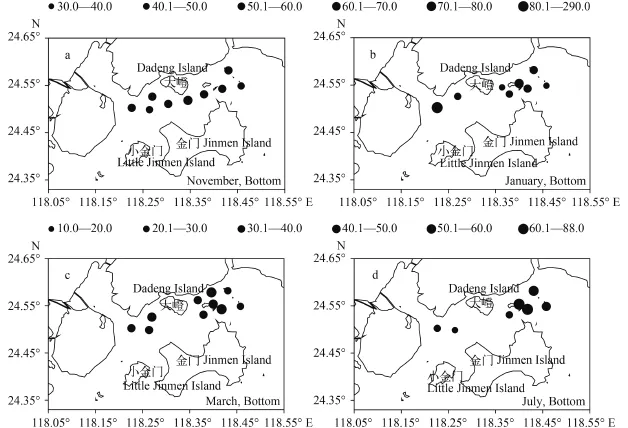
图 3 底层浮游植物丰度的平面分布(×103cells/L)Fig. 3 Horizontal distribution of phytoplankton abundance in bottom waters of the Northern Jinmen Island
2.4 浮游植物多样性评价
物种多样性可用于评价群落中物种组成的稳定程度及其数量分布的均匀程度和群落的组织结构特征。参照陈清潮等[21]提出的热带海区生物多样性阈值范围, 浮游植物多样性程度根据多样性指数的大小可分为5级: <0.6 为差; 0.6—1.5为一般; 1.6—2.5为较好; 2.6—3.5为丰富; >3.5为非常丰富。本次调查(表 4)的四季多样性均值为2.94, 介于2.6—3.5, 表征丰富的浮游植物物种多样性程度。多样性指数的四季变幅除1月表层(1.93)以及3月底层(1.84)的低值外, 多样性指数在各季节的各站位分布较均匀。四季以11月浮游植物物种多样性非常丰富, 11月表底的多样性均值均>3.5。3月表底的多样性均值最低, 介于1.6—2.5, 为较好级别。7月表底的多样性均值略高于3月表底, 二者都介于2.6—3.5, 表现丰富级别的浮游植物物种多样性程度。柳丽华等[20]研究认为, 群落中的单一或某几个物种优势度越明显, 种类的多样性就会降低。本区3月浮游植物物种多样性低的原因即是优势种集中为标志星杆藻、具槽帕拉藻以及骨条藻的缘故。
均匀度代表群落内物种分布的均匀程度, 是群落是否成熟和稳定的特征之一。孟顺龙等[22]认为,均匀度指数值越大, 表明多样性越高, 生态环境状况越好, 但孙军等[19]认为也非绝对, 多样性指数有时与均匀度指数同样趋势一致, 但代表意义截然不同, 可以确定的是, 较为稳定的群落一般具有较高的多样性和均匀度。从本研究的浮游植物均匀度分析结果(表 4)来看, 浮游植物均匀度变幅不大, 站间差异小, 在各季节的各站位分布较平缓, 四季均值为 0.80, 与多样性指数相似, 3月均匀度最低, 11月物种间丰度分配均匀, 其均匀度最高。均匀度与多样性指数的季节变化特点基本一致, 群落多样性高的季节或海域物种均匀度也好。
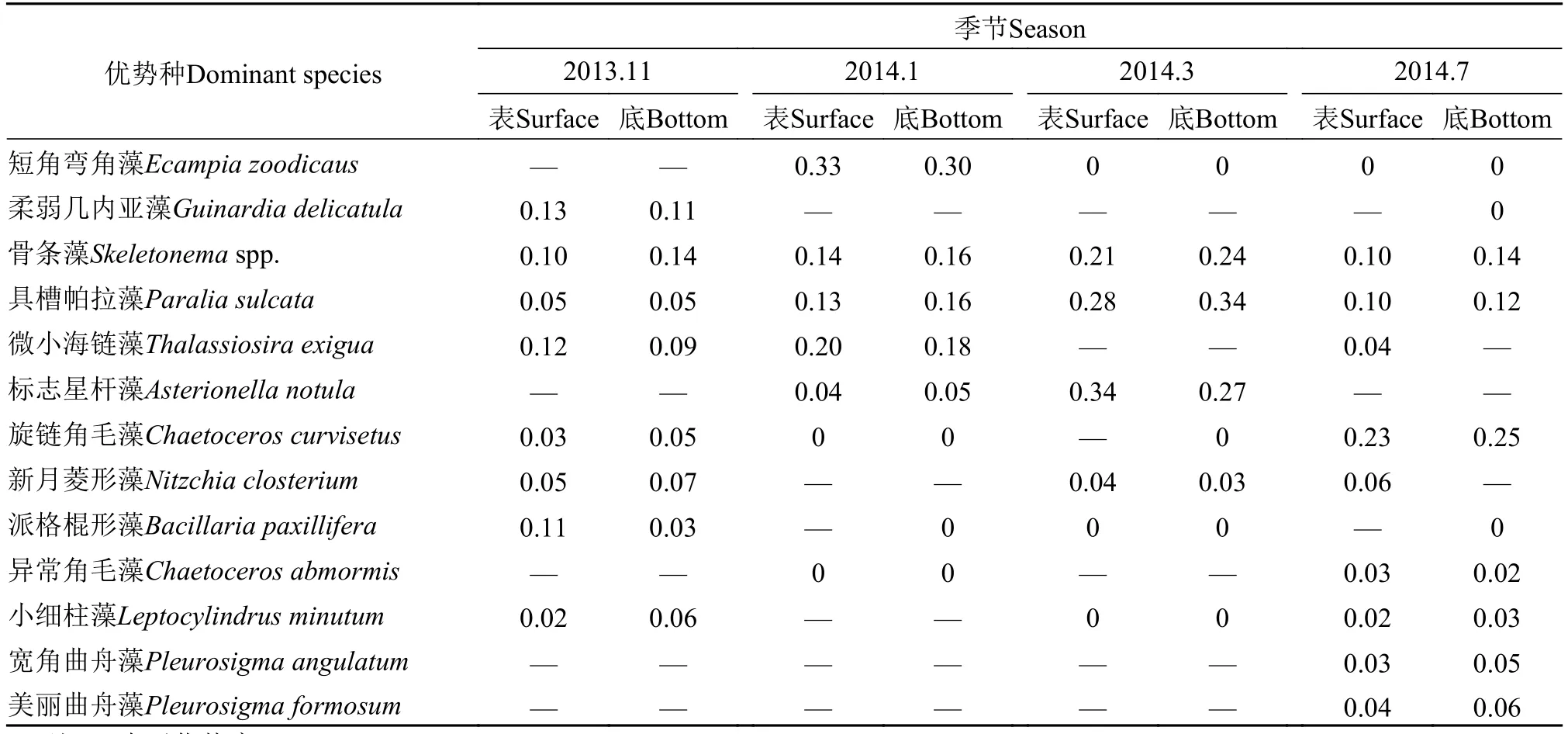
表 3 金门北部海域的浮游植物主要优势种及优势度Tab. 3 The dominant phytoplankton and their dominance in sea waters of the Northern Jinmen Island
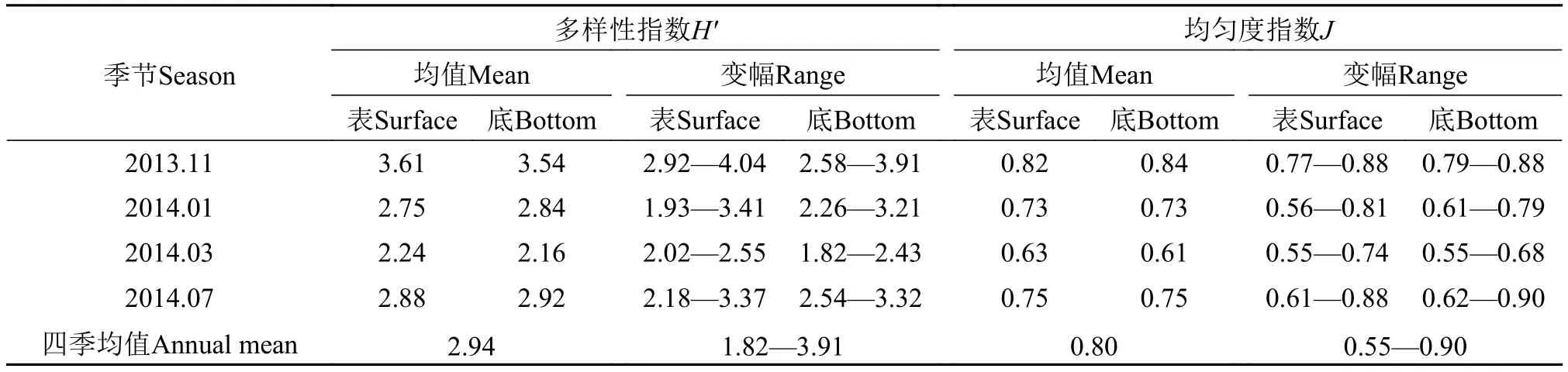
表 4 金门北部海域浮游植物多样性的季节变动Tab. 4 Seasonal variation of diversity and evenness of phytoplankton in sea waters of the Northern Jinmen Island
3 讨论
3.1 本区与毗邻海区的比对分析
本海区毗连厦门东海域, 与厦门西港、九龙江河口、厦门东北部同安湾毗邻, 与之相比(表 6), 本区种类组成与其他海区相似, 硅藻主导型群落, 甲藻次之, 蓝藻较少, 金藻出现率低至无检出。本区浮游植物物种丰富度远低于前期大嶝岛四季的物种数目[13,14], 接近厦门东部四季度月的物种数目[11]。我国早期对浮游植物丰度的调查方法主要遵从《海洋调查规范》[15]中网采的方法, 网目大小多为76 μm。Hensen和Lohmann研究[24,25]认为, 网采样不能将水体中全部粒径谱的浮游植物采获, 从网中容易遗漏一些比网目小的种类, 是一种半定量的方法, 但此方法在大洋贫营养海域及与历史资料参比方面有一定优势, 且相对于网具所能捕获的大型浮游生物(>200 μm)来说, 其损失的分量是不明显的[23]。但由于拖网浓缩的海水体积较大, 所采集的浮游植物样品中物种数目也会较高, 因此可见表5中的厦门周边海域浮游植物网样的物种数目均高于金门北部浮游植物水样的物种数目。
表 5可见, 本文分析水样所得的浮游植物丰度与网样分析的结果相比较高, 远高于前期以往大嶝岛调查结果, 但1990年在厦门东部的网采浮游植物丰度出现一个高值, 起因是厦门东北部同安湾3个站位的四季都出现高丰度, 并在春季3月出现浮游植物藻华[10,12]。表 4也表明毗邻海区之间的网样分析结果自身差别较大, 这也进一步说明拖网方法会有部分样品的丢失, 尤其是粒径小于网孔的浮游植物, 以及拖网过程中倾角的变化也会影响过滤水柱体积的计算等因素[23], 会造成拖网分析结果是非准确和非精确定量的。因而, 受拖网网目(一般为76 μm)影响及拖网时倾角的变化会低估浮游植物丰度。因此作者建议浮游植物生态研究方法尽量与国际通用方法接轨, 为资料的共享和利用提供便利, 以定量采水样和Utermöhl方法分析水样替代半定量的网采水样和常规光学显微镜分析。
此外, 受调查时间和调查站位差异的影响, 不同调查中浮游植物优势种的构成有一定的差异(表6)。优势种在群落中起着主导作用, 决定了群落内的物质循环和能量流动的方式, 支配着群落的演替方向。如果一个优势种群从群落内消亡, 不仅群落结构要发生变化, 群落内的生态环境也会发生较大的改变[20,26]。本次调查的优势种较之毗连海域季节动态, 优势种构成明显与之不同(表 6), 前期研究表明[8—14], 厦门岛周边海域的浮游植物主要以角毛藻属、圆筛藻属和骨条藻属为主体。冬季(12—2月)以圆筛藻属为主, 角毛藻属为次; 春季(3—5月)和夏季(6—9月)则分别以角毛藻属和骨条藻属占绝对优势; 秋季(10—11月), 主要属呈现多样化, 除了上述3个属比例稍大外, 菱形藻属、星杆藻属(Asterionella)在夏、秋两季也有一定的数量。从表 6可见, 具槽帕拉藻成为本区四季优势种, 而在前期报道仅在冬季1月占优[13,14]; 圆筛藻的优势地位全无, 海链藻(Thalassiosira)跃居成为优势种。
以往研究认为, 中肋骨条藻是厦门海区常见的优势种[8—14], 在季节变化上具有“暖水性”[27], 数量高峰期出现于夏季[11—14]。近期, 国内关于骨条藻分类研究有了突破, 并提供了厦门港骨条藻相关形态学特征及其季节变化[28]: 根据透射电镜分析, 在厦门港有六种骨条藻分布, 中肋骨条藻是唯一一种在周年每月均有分布的骨条藻, 水体中的中肋骨条藻在厦门港冬春季丰度较低, 夏秋季丰度较高, 占据优势地位。多尼骨条藻(Skeletonema dohrnii)主要在冬季和春季出现, 在水体中占据优势地位, 到了夏季和秋季则在水体中消失。曼氏骨条藻(Skeletonema menzelii)、热带骨条藻(Skeletonema tropicum)、亚当斯骨条藻(Skeletonema ardens)和桂氏骨条藻(Skeletonema grevillei)都是一般在夏季出现[28]。
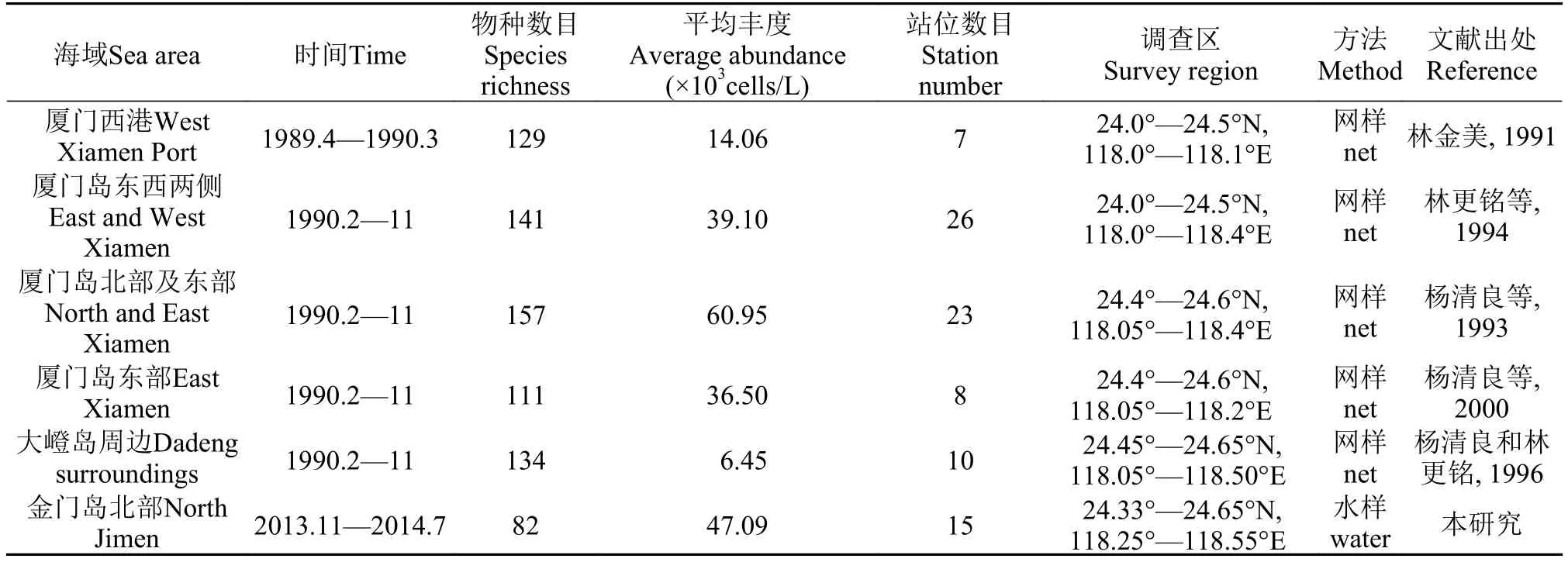
表 5 金门北部与毗邻海域的浮游植物生态特征值比较Tab. 5 Comparison of ecological characters of phytoplankton between sea waters of the Northern Jinmen Island and its adjacent sea regions
3.2 浮游植物丰度变动与环境因子的关联
浮游植物生态现状与环境因子密切相关。通过对浮游植物丰度与环境因子的Pearson 相关性分析(表 7)表明, 水温仅在11月与浮游植物丰度呈现显著的负相关, 盐度在11月和1月均与丰度呈现极显著的负相关, 悬浮物含量与1月丰度正相关关系明显, 与7月丰度负相关; 溶解无机氮和活性磷酸盐仅在1月对浮游植物丰度起极显著的促进增长作用。3月丰度与上述环境因子的关联作用不明显。
本次调查海区水温在13.21—26.92℃波动, 四季温差较大; 盐度在29.41—31.41变化, 盐度1月最低, 3月最高, 而水温7月最高, 3月最低。水温在11月适宜, 平均20.87℃, 利于浮游植物生长, 但本区浮游植物丰度却为四季最低值, 二者负相关显著。11月盐度接近于1月低值, 对浮游植物生长繁殖的促进作用不强, 近岸的盐度变化与大陆径流密切相关, 而大陆径流的输入必然带来丰富的营养, 在盐度的变化值并没有超过浮游植物耐受的程度时, 本区的浮游植物丰度随着盐度的下降而上升, 并一直延续到1月, 此时, 1月近岸(低盐)受陆源径流影响大[12,14,29], 营养盐含量通常比外海(高盐)丰富, 而浮游植物的生长、繁殖主要受制于水域中的营养盐,这样浮游植物丰度与盐度的关系以负相关的面貌出现, 而与溶解无机氮和活性磷酸盐的正面促进非常明显(表 7), 悬浮物是对1月浮游植物丰度显著正相关的一个作用因子。1月高营养盐含量为四季最高, 促进了浮游植物高丰度的形成。本区7月悬浮物含量最低(19.65 mg/L), 浮游植物丰度为四季次高值(41.05×103cells/L), 体现负相关关系; 1月悬浮物含量(26.93 mg/L)略高于7月含量少许, 浮游植物丰度为四季最高(73.28×103cells/L); 而3月和11月悬浮物含量处于较高水平(71.97 mg/L, 37.37 mg/L),浮游植物丰度是四季的低值(3月39.29×103cells/L, 11月34.73×103cells/L)。
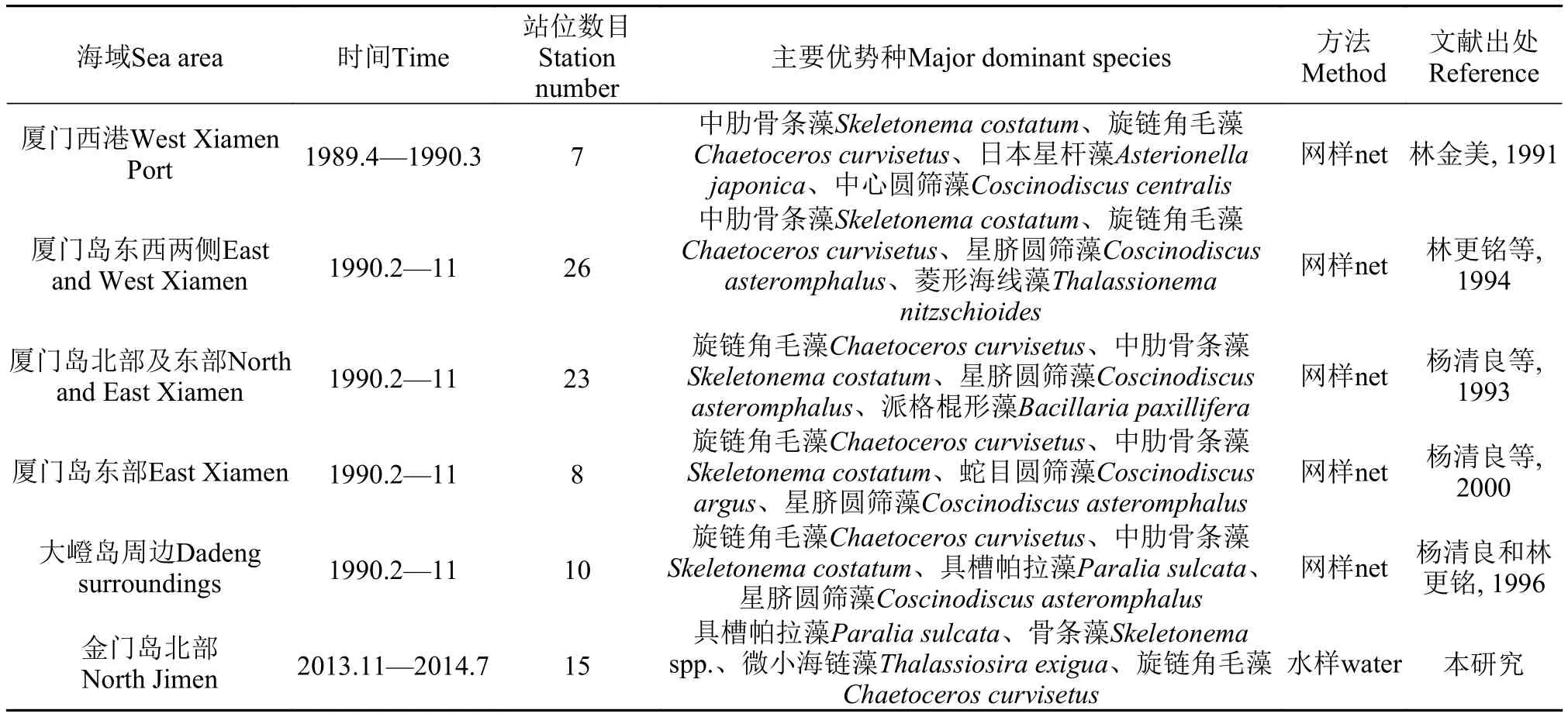
表 6 金门北部和毗邻海域四季的浮游植物优势种比较Tab. 6 Comparison of dominant phytoplankton in sea waters of the Northern Jinmen Island and its adjacent sea regions during one year
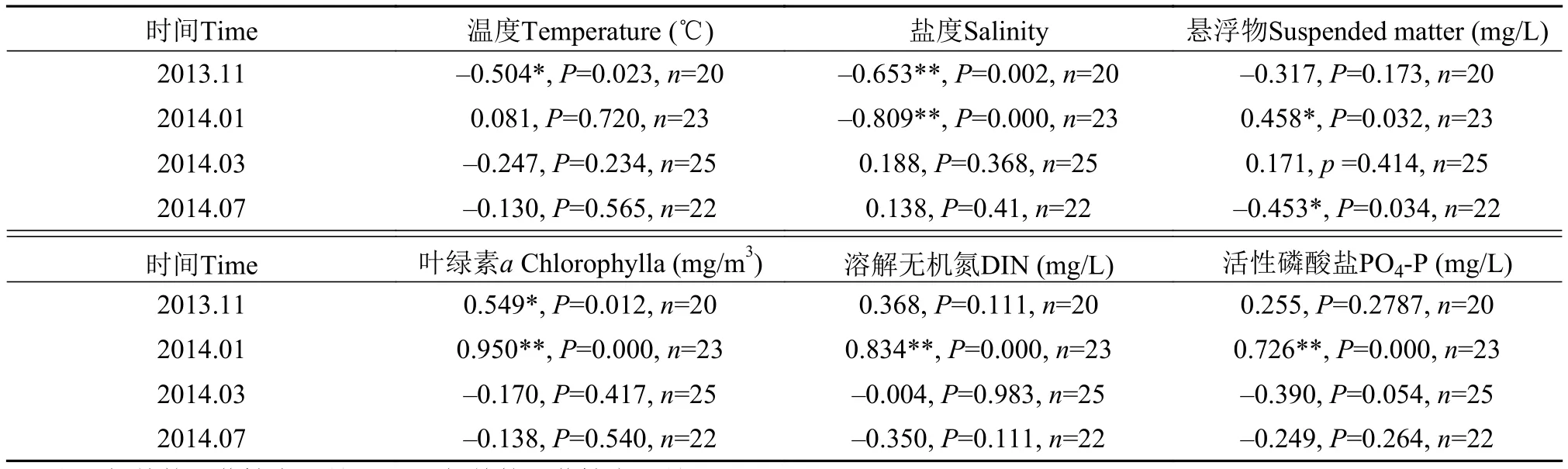
表 7 浮游植物丰度与环境因子的Pearson相关性Tab. 7 Pearson correlation analysis between phytoplankton abundance and environmental factors
将各季浮游植物丰度与同期调查的叶绿素a浓度进行线性回归分析, 发现除了11月和1月丰度与之有较好的正相关外, 浮游植物丰度与叶绿素浓度的关系不显著(表 7)。但1月浮游植物的高丰度与此时叶绿素a浓度高值的正相关关系显著的凸现出来,叶绿素a浓度与浮游植物丰度在高丰度发生期间(本海区1月丰度极高)的线性关联尤为明显[30,31]。
3.3 探讨海域工程对浮游植物群落的潜在影响
金门北部海域受到西部九龙江水、东部晋江支流小型水系以及台湾海峡外海水的共同影响, 这一区域潮大、流态复杂、含沙量较低[1,29]。在厦门机场二期工程施工过程中, 工程建设产生的非点污染源(广义指各种没有固定排污口的环境污染)主要来自于岛区码头及填海造地的建设施工、炸礁及航道疏浚等, 总悬浮微粒产生源为疏浚、抛泥和吹泥, 污染源包括岛上配套工程区生活污水和油污水等。这些污染都会对工程区及邻近海域海洋生态系统产生不同程度的影响[32,33]。其中, 主要影响因素为工程施工产生的大量悬浮物和冲击波。工程建设施工期的生活污水影响较小, 可以忽略。
研究认为[3,4,34], 近海海洋工程在施工过程中,通常产生一部分泥沙与海水混合, 形成悬沙含量很高的水团, 从而大大增加了水中悬浮物质的含量。根据2013—2014年同期悬浮物的监测数据分析, 本区悬浮物季节变化明显, 表底分布差异显著[29]。1月和7月悬浮物含量低, 浮游植物丰度较高; 而3月和11月悬浮物含量较高, 浮游植物丰度低。徐兆礼等[35]发现悬浮物质的增加, 最直接的影响是削弱了水体的真光层厚度, 从而降低了海洋初级生产力,使浮游植物丰度下降, 并造成优势种更替混乱。其次, 水中悬浮物质含量的增多, 对浮游植物物种的存活和繁殖有着明显的抑制作用[35,36]。但施工期所产生的悬浮物对海洋生物的影响在时间尺度上也是暂时的[35,37]。大量海洋工程案例[2—5,35—37]表明, 在施工期结束后, 水体中的悬浮物会很快恢复到施工期的浓度水平, 海洋生态系统也会很快恢复。再有, 水下爆破所产生的巨大冲击波对施工水域内的鱼类造成直接伤害外, 还会使安全区以外的鱼类迅速出逃, 起到大范围驱赶作用, 且对于生物幼体损害较大[5,36], 但对周围水域浮游生物的种类组成和丰度没有明显影响[36]。
本文结果可以作为厦门机场二期工程前期的本底背景, 待厦门机场二期工程完工之际再添加相应的浮游植物群落数据对比分析, 进而可对海洋工程开建的生态影响进行评估。
4 结论
由于金门岛特殊的地理位置和历史渊源, 在台海两岸的交流与互动中扮演着重要的角色和地位,对两岸生态环保的协同合作有示范作用。本文基于2013—2014年四季度月在金门岛北部海域获取的浮游植物样品分析, 共鉴定浮游植物3门43属82种(不含未定种), 群落构成以硅藻为主, 其次是甲藻, 蓝藻最少。物种组成以丰度的季节差异较大。四季优势种均为硅藻, 不同季节优势种有一定程度交错, 且变化较明显。6个优势种的优势度在表层小于底层。3月的浮游植物群落状态相对较脆弱。溶解无机氮及活性磷酸盐是促使1月丰度全年最高的原因。大量工程建设案例表明, 涉海工程施工期造成的浮游植物丰度下降趋势和优势种更替混乱在工程结束后能得以恢复。待厦门机场二期工程完工之际再添加相应的浮游植物群落数据对比分析, 进而评估工程开建的生态影响。
[1]Chen J R. Characteristic analysis of tidal current energy in the Xiamen Bay and Jinmen channel [J]. Marine Science Bulletin, 2013, 32(6): 641—647 [陈金瑞. 厦门湾海域及金门水道潮流能特征分析. 海洋通报, 2013, 32(6): 641—647]
[2]Wu H Y, Zhu W B, Chen X H, et al. Effects of enclosingbank and hydraulic fill projects on water environment [J]. Water Resources Protection, 2005, 21(2): 53—56 [吴英海, 朱维斌, 陈晓华, 等. 围滩吹填工程对水环境的影响分析. 水资源保护, 2005, 21(2): 53—56]
[3]Jia Y R, Sun Y L, Zhang X Q. Study on suspended particulate matter simulation during harbor dredging [J]. Technique of Port and Harbor, 2007, 3: 3—5 [贾怡然, 孙英兰, 张学庆. 港池疏浚过程悬浮物影响预测研究及应用.港工技术, 2007, 3: 3—5]
[4]Che H Y. Impact of suspended substance from the expansion engineering of Yingkou port on oceanic environment [J]. Journal of Meteorology and Environment, 2006, 22(2): 48—50 [车宏宇. 营口港扩建工程悬浮物对海域环境影响分析. 气象与环境学报, 2006, 22(2): 48—50]
[5]Cui Y, Song Y L, Lin Q L, et al. Effects of explosion in water column on marine organisms and marine environment [J]. Journal of Fishery Science of China, 1996, 3(4): 106—113 [崔毅, 宋云利, 林庆礼, 等. 水中爆破对海洋生物及海洋环境的影响研究. 中国水产科学, 1996, 3(4): 106—113]
[6]Gayoso A M. Long-term phytoplankton studies in the Bahia Blance estuary, Argentina [J]. Ices Journal of Marine Science, 1998, 55(4): 655—660
[7]Kamenir Y Z, Dubinsky T Z, Zohary T. Phytoplankton size structure stability in a meso-eutrophic subtropical lake [J]. Hydrobiologia, 2004, 520(1): 89—104
[8]Lin J M. Phytoplankton ecology in western Xiamen Harbour [J]. Journal of Oceanography in Taiwan Strait, 1991, 10(4): 345—350 [林金美. 厦门西海域浮游植物的生态. 台湾海峡, 1991, 10(4): 345—350]
[9]Chen Q H, Zeng Z W, Zhang S J, et al. Report on red occurred in Xiamen Harbour in 1987 [M]. In: Third Institute of Oceanography, SOA (Eds.), Collected Papers on Red Tide Survey and Study in Xiamen Harbour. Beijing: China Ocean Press. 1993, 1—18 [陈其焕, 曾昭文, 张水浸, 等. 厦门港1987年赤潮调查报告. 见: 国家海洋局第三海洋研究所编著, 厦门港赤潮调查研究论文集. 北京:海洋出版社. 1993, 1—18]
[10]Yang Q L, Lin G M, Lin J M. Ecological studies of phytoplankton in waters around Xiamen Amphioxus Reserve Area [J]. Journal of Oceanography in Taiwan Strait, 1993, 12(3): 205—217 [杨清良, 林更铭, 林金美.厦门文昌鱼自然保护区周围海域浮游植物的生态. 台湾海峡, 1993, 12(3): 205—217]
[11]Yang Q L, Lin G M, Cai B J. Species composition and distribution of phytoplankton in eastern Xiamen waters [J]. Journal of Oceanography in Taiwan Strait, 2000, 19(3): 337—343 [杨清良, 林更铭, 蔡秉及. 厦门东侧海域浮游植物的种类组成与分布. 台湾海峡, 2000, 19(3): 337—343]
[12]Lin G M, Yang Q L, Lin J M. Relationship between phytoplankton and environment factors in waters around Xiamen Island [J]. Marine Science Bulletin, 1993, 12(6): 40—45 [林更铭, 杨清良, 林金美. 厦门岛周围海域浮游植物与环境因子的关系. 海洋通报, 1993, 12(6): 40—45]
[13]Lin G M, Yang Q L, Lin J M. Abundance and species composition of phytoplankton in waters around Xiamen Island [J]. Journal of Oceanography in Taiwan Strait, 1994, 13(4): 353—358 [林更铭, 杨清良, 林金美. 厦门岛周围海域浮游植物的种类组成及丰度. 台湾海峡, 1994, 13(4): 353—358]
[14]Yang Q L, Lin G M. Research report of phytoplankton biodiversity in Xiamen Island [M]. In: The Multidisciplinary Oceanographic Investigation Office of Xiamen (Eds.), Marine Biodiversity Research in Xiamen Island and Neighbouring Area. Beijing: China Ocean Press. 1996, 112—118 [杨清良, 林更铭. 厦门市海岛资源综合调查研究报告-浮游生物部分. 见: 厦门市海岛资源综合考察办公室编著, 厦门市海岛资源综合调查研究报告. 北京: 海洋出版社. 1996, 112—118]
[15]State Bureau of Technical Supervise. The Specification for Oceanographic Survey-Marine Biological Survey (GB/T12736.6-2007) [S]. Beijing: China Standard Press. 2007, 30—35 [国家技术监督局.《海洋调查规范》(GB/T12736.6-2007). 北京: 中国标准出版社. 2007, 30—35]
[16]Sun J, Liu D Y. The preliminary notion on nomenclature of common phytoplankton in China Sea waters [J]. Oceanologia et Limnologia Sinica, 2002, 33(3): 271—286 [孙军, 刘东艳. 中国海区常见浮游植物种名更改初步意见. 海洋与湖沼, 2002, 33(3): 271—286]
[17]Chen J F, Gao Y H, Liang J R, et al. Research progress in species and gene diversity of Skeletonema, Bacillariophyta [J]. Advance in Natural Science of China, 2007, 17(5): 586—594 [陈金凤, 高亚辉, 梁君荣, 等. 骨条藻的种类与基因多样性研究进展. 自然科学进展, 2007, 17(5): 586—594]
[18]Gao Y H, Chen C P, Li Y. Marine nanophytoplankton diatoms from coastal waters of Hong Kong [C]. In: Morton B (Eds.), Perspective on Marine Environment Change in Hongkong and Southern China. Hongkong. 2003, 93—107
[19]Sun J, Liu D Y. The application of diversity indices in marine phytoplankton studies [J]. Acta Oceanologia Sinica, 2004, 26(1): 62—75 [孙军, 刘东艳. 多样性指数在海洋浮游植植物研究中的应用. 海洋学报, 2004, 26(1): 62—75]
[20]Liu L H, Zuo T, Chen R S, et al. Community structure and diversity of phytoplankton in the estuary of Yangtze River in Autumn [J]. Marine Fisheries Research, 2007, 28(3): 112—119 [柳丽华, 左涛, 陈瑞盛, 等. 2004年秋季长江口海域浮游植物的群落结构和多样性. 海洋水产研究, 2007, 28(3): 112—119]
[21]Chen Q C, Huang L M, Yin J Q, et al. Biodiversity of zooplankton in Nansha Islands [M]. In: The Multidisciplinary Oceanographic Expedition Team of Academia Sinica to Nansha Islands (Eds.), Marine Biodiversity Research in Nansha Islands and Neighbouring Area. Beijing: China Ocean Press. 1994, 42—50 [陈清潮, 黄良民, 尹建强, 等. 南沙群岛海区浮游动物多样性研究. 中国科学院南沙综合科学考察队. 南沙群岛及其邻近海区海洋生物多样性研究. 北京: 海洋出版社. 1994, 42—50]
[22]Meng S L, Chen J C, Hu G D, et al. Phytoplankton community characteristics and its eco-assessment on water quality in Lihu Lake, Taihu Lake [J]. Resources and Environmental in the Yangtze Basin, 2010, 19(1): 30—36 [孟顺龙, 陈家长, 胡庚东, 等. 太湖蠡湖浮游植物群落特征及其对水质的评价. 长江流域资源与环境, 2010, 19(1): 30—36]
[23]Sun J, Liu D Y, Qian S B. A quantitative research and analysis method for marine phytoplankton: An introduction to Utermöhl method and its modification [J]. Journal of Oceanography of Huanghai & Bohai Seas, 2002, 20(2): 105—112 [孙军, 刘东艳, 钱树本. 一种海洋浮游植物定量研究分析方法–Utermöhl方法的介绍及其改进. 黄渤海海洋, 2002, 20(2): 105—112]
[24]Hensen V.ü ber die Bestimmung des Planktonsoder desim Meeretreibenden Materials an Pflanzen und Thieren. Kommission zurwissenschaftlichen Untersuchung der deutschen Meere in Kiel, 1882—1886 [J]. V Bericht Jahrgang, 1887, 12—16, 1—107
[25]Lohmann H. Ueber das Fischenmit Netzen aus Müllergaze Nr 20 zu dem Zwecke quantitativen Untersuchungen des Auftribs. Wissenschaftliche Meeresuntersuchungen [J]. Abteilung Kiel Neue Folge, 1901, 5: 46—66
[26]Zhang F, Shangguan T L. Analysis on the ecological dominance of the Taxus mairei forest community in Shanxi [J]. Journal of Shanxi University (Natural Science), 1988, 11(3): 82—87 [张峰, 上官铁梁. 山西南方红豆杉(Taxus mariei)森林群落的生态优势度分析. 山西大学学报(自然科学版) , 1988, 11(3): 82—87]
[27]Jin D X, Chen J H, Huang K G. China Marine Planktonic Diatom [M]. Shanghai: Shanghai Science and Technology Press. 1965, 63—64 [金德祥, 陈金环, 黄凯歌. 中国海洋浮游硅藻类. 上海: 上海科学技术出版社. 1965, 63—64]
[28]Gu H F, Zhang X D, Sun J, et al. Diversity and seasonal occurrence of Skeletonema (Bacillariophyta) species in Xiamen Harbor and surrounding seas, China [J]. Cryptogamie Algologie, 2012, 33(3): 245—263
[29]Lin H, Ji W D. The investigation for oceanographic survey-observations of chemical parameters in the second phase project of the Xiamen Airport [R]. Third Institute of Oceanography, State Oceanic Administration. 2014, 19—25 [林辉, 暨卫东. 厦门机场二期工程海域的化学要素调查研究报告. 国家海洋局第三海洋研究所, 2014, 19—25]
[30]Yu H Y, Zhou B, Hu Z Y, et al. Study on correlation between chlorophyll a and algal density of biological monitoring [J]. Environmental Monitoring in China, 2009, 25(6): 40—44 [于海燕, 周斌, 胡尊英, 等. 生物监测中叶绿素a浓度与藻类密度的关联性研究. 中国环境监测, 2009, 25(6): 40—44]
[31]Rakocevic-Nedovic J, Hollert H. Use of chlorophyll-Secchi disk relationships, phytoplankton community and chlorophyll a as trophic state indices of Lake Skadar (Montenegro, Balkan) [J]. Environmental Science and Pollution Research, 2005, 12(3): 146—152
[32]Xu Z L, Zhang F Y, Chen Y Q. Assessment on fishery resource loss owing to mechanical entanglement and residual chlorine [J]. Marine Environmental Science, 2007, 26(3): 246—251 [徐兆礼, 张凤英, 陈渊泉. 机械卷载和余氯对渔业资源损失量评估初探. 海洋环境科学, 2007, 26(3): 246—251]
[33]Xu Z L, Chen H. Estimating economic costs of potential fishery losses caused by marine engineering [J]. Journal of Fishery Science of China, 2008, 15(6): 970—975 [徐兆礼, 陈华. 海洋工程环境评价中渔业资源价值损失的估算方法. 中国水产科学, 2008, 15(6): 970—975]
[34]Filipsson H L, Bjork G, McQuoid M R. A major change in the phytoplankton of a Swedish sill fjord-A consequence of engineering work [J]? Estuarine, Coastal and Shelf Science, 2005, 63: 551—560
[35]Xu Z L, Zhang F Y, Chen Y Q. Quantitative analysis on fishery resource loss owing to the influence of suspended substance and shock wave [J]. Journal of Fisheries ofChina, 2006, 30(6): 778—784 [徐兆礼, 张凤英, 陈渊泉.悬浮物和冲击波造成的渔业资源损失量估算. 水产学报, 2006, 30(6): 778—784]
[36]Jia X P, Lin Q, Cai W G. Evaluation for the impact of large explosion at Mabianzhou Island on the neighboring aquatic environment and marine organisms in Daya Bay [J]. Journal of Fisheries of China, 2002, 26(4): 313—320 [贾晓平, 林钦, 蔡文贵. 大亚湾马鞭洲大型爆破对周围水域环境与海洋生物影响的评估. 水产学报, 2002, 26(4): 313—320]
[37]Jiang M, Shen X Q, Yang H. Influence of underwater explosions on fishery organism [J]. Marine Fisheries, 2005, 27(2): 150—153 [蒋玫, 沈新强, 杨红. 水下爆破对渔业生物影响的研究. 海洋渔业, 2005, 27(2): 150—153]
SEASONAL DYNAMICS OF PHYTOPLANKTON COMMUNITY AND ITS RELATION WITH ENVIRONMENTAL FACTORS IN SEA WATERS OF THE NORTHERN JINMEN ISLAND
WANG Yu, XIANG Peng, YE You-Yin, LIN Geng-Ming, YANG Qing-Liang and LIN Mao
(Third Institute of Oceanography, State Oceanic Administration, Xiamen 361005, China)
Based on four cruise surveys carried out from 2013 to 2014, we analyzed phytoplankton community structure and seasonal pattern and compared them with those of the adjacent sea area. We also assessed the relationship between phytoplankton and environmental factors and potential influence by the second phase of the Xiamen Airport Project construction on phytoplankton. A total of 82 species (excluding unknown taxa) belonging to 43 genera of 3 classes were identified from 95 samples, in which diatoms were dominant in both the species composition and the abundance. Dinoflagellates was the second-largest group. Cyanobacteria was the least with only one species. Significant seasonal changes existed for phytoplankton species composition and abundance. Species richness was the lowest in March and the highest in November. The annual average phytoplankton abundance was 47.09×103cells/L with the peak in January and the minimum in November. The dominant species were diatoms including thirteen species: Guinardia delicatula, Ecampia zoodicaus, Skeletonema spp., Paralia sulcata, Thalassiosira exigua, Asterionella notula, Chaetoceros curvisetus, Nitzchia closterium, Bacillaria paradoxa, Chaetoceros abmormis, Leptocylindrus minutum, Pleurosigma angulatum and Pleurosigma formosum. In detail, Paralia sulcata and Skeletonema spp. were dominant species in four seasons. Shannon-wiener diversity index and Pielou’s evenness index indicated a more integrated and stable state of phytoplankton community. Compared with adjacent sea areas around Xiamen Island, the Northern Jinmen Island had medium level of richness of phytoplankton species, similar species composition but different dominant species and higher abundance. Pearson Correlation Analysis indicated that phytoplankton abundance was remarkably negatively associated with water temperature in November and was significantly negatively linked with salinity in January and November. Phytoplankton abundance were correlated positively to suspended substance in January but negatively linked in July. The phosphate and dissolved inorganic nitrogen were significantly positively linked with phytoplankton abundance in January. Chlorophyll a content was positively associated with phytoplankton abundance in November and significantly positive in January. Phytoplankton abundance in March was not clearly related to environmental factors. The second phase of Xiamen Airport Project mainly affected phytoplankton by massive suspended substance and shock wave during construction period. Moreover, ocean engineering cases indicated that phytoplankton abundance of investigated water began to rise and the disorder of dominants succession gradually recovered to the pre-construction status. More investigations of phytoplankton after completing ocean engineering projects are required to assess ecological affect by the second phase of Xiamen Airport Project.
Phytoplankton; Community structure; Seasonal dynamics; Environmental factors; Correlation analysis; Sea waters of the northern Jinmen Island
Q145+.2
A
1000-3207(2017)03-0712-12
10.7541/2017.89
2016-05-10;
2016-10-14
科技部基础专项(GASI-01-02-04); 科技部海洋公益性行业科研专项(201005015)资助 [Supported by the Key Project of Basic Research Program of Ministry of Science and Technology (GASI-01-02-04), the Marine Public Research Project of Ministry of Science and Technology (201005015)]
王雨(1981—), 男, 贵州长顺人; 副研究员; 主要从事浮游生物生态的研究。E-mail: wangyu@tio.org.cn
林茂, E-mail: linmao@tio.org.cn
