以PRDM1基因启动子为靶点的药物筛选模型的建立研究*
2017-05-03胡文坛王婷婷张世杰张靖宇刘红春
胡文坛,王婷婷,张世杰,何 鑫,张靖宇,刘红春△
(1.郑州大学第一附属医院检验科,郑州 450003;2.河南中医学院第一附属医院检验科,郑州 450002)
·论 著·
以PRDM1基因启动子为靶点的药物筛选模型的建立研究*
胡文坛1,王婷婷1,张世杰1,何 鑫2,张靖宇1,刘红春1△
(1.郑州大学第一附属医院检验科,郑州 450003;2.河南中医学院第一附属医院检验科,郑州 450002)
目的 以PRDM1基因启动子为靶点构建双荧光素酶报告基因载体,建立体外药物筛选细胞模型,并对中草药小分子化合物进行筛选。方法 将人PRDM1基因启动子序列(267、1 257 bp)克隆入荧光素酶报告基因载体pGL3-Basic中,构建重组质粒pGL3-PRDM1并与内参质粒pRL-TK瞬时共转染工具细胞,通过检测荧光素酶报告基因表达水平的变化反映PRDM1基因启动子的启动转录活性,并对共转染质粒比例、工具细胞选择等条件进行探索和优化,相关药物处理进行验证。结果 成功构建了荧光素酶报告载体pGL3-PRDM1。用0.5、1、2、4 μmol/L的5-Aza-CdR处理重组的U266细胞筛选模型,与0 μmol/L 5-Aza-CdR相比,增加5-Aza-CdR 的刺激,荧光强度及PRDM1基因启动子的活性呈剂量依赖性增强。与0 μmol/L相比,青蒿琥酯从5 μmol/L浓度开始对PRDM1基因启动子活性呈明显降低(P<0.05),在20 μmol/L浓度时降到最低;白芍总苷呈现小剂量促进PRDM1启动子活性,高剂量抑制PRDM1基因启动子活性。青蒿琥酯对细胞生长增殖影响较小,白芍总苷对细胞生长增殖的影响较复杂。结论 成功建立了以PRDM1基因启动子为靶点的药物筛选模型,并筛选出青蒿琥酯能抑制PRDM1基因启动子活性。
PRDM1基因;双荧光素酶报告基因;中草药;药物筛选
B淋巴细胞诱导成熟蛋白(B lymphocyte induced maturation protein,Blimp)-1,称为正向调节结构域锌指蛋白(positive regulatory domain zinc finger protein,PRDM)1,由PRDM1基因编码,具有5个锌指结构。Morgan等[1]研究结果证实,Blimp-1基因存在不同的转录起始位点和亚型,并于外显子1上游70 kb处存在1个附加外显子,使其具有2种亚型:Blimp-1α和-1β,分别由PRDM1α和PRDM1β编码,前者在调节浆细胞分化发挥重要的转录因子作用,后者由于功能缺陷失去了对多个靶基因的调控作用,所以传统意义上所说的Blimp-1蛋白是指Blimp-1α[2]。在B细胞系,PRDM1基因表达的Blimp-1特异地表达于所有抗体分泌细胞,包括浆细胞和浆母细胞[3],是维持骨髓中长寿浆细胞形成及Ig分泌所必需的转录因子,是长期维持抗原特异免疫反应所必需。正常情况下,Blimp-1参与机体内免疫应答的形成并维持稳定状态,但Blimp-1表达或调控异常时,将使免疫功能紊乱,多克隆B细胞激活、多种自身抗体大量产生,诱发系统性红斑狼疮(SLE)、类风湿关节炎(RA)、多发性硬化、肾小球肾炎、自身免疫性溶血性贫血等疾病,而干扰Blimp-1表达会使相关自身抗体水平降低,抑制B细胞向浆细胞分化,病情得到缓解,发病延缓[4-5]。本实验旨在通过建立以PRDM1基因启动子为靶点的高通量药物筛选模型寻求Blimp-1表达下调剂,在分离提取的中草药免疫活性成分中进行筛选。包括多糖类、皂苷类、类黄酮类、有机酸类、多酚类和萜类等多种成分被证明能调节机体免疫功能作用,在免疫相关疾病的治疗上也收到较好的临床疗效[6]。 根据相关文献报道,选取PRDM1基因转录起始位点上游267 bp和1 257 bp序列作为本模型的目标启动子序列,再结合双荧光素酶报告基因技术,构建Blimp-1活性抑制剂细胞筛选模型,双荧光素酶报告基因检测系统是目前从分子水平用于研究分析基因转录调控的有效工具,与RT-PCR相比更快速、灵敏、简便、适用于高通量筛选等优点。Mundy等[7]将含小鼠BMP-2启动子的荧光素酶报告基因质粒转入小鼠成骨细胞系,对3万多种化合物进行筛选,发现洛代他汀可增加报告基因活性,从而寻求到可刺激骨形成的新药物。国内外目前基于该方法构建以PRDM1/Blimp-1基因为靶点的免疫抑制药物筛选模型的报道尚少见。
1 材料与方法
1.1 主要试剂和仪器 293T人胚肾上皮细胞,U266、ARH77人多发性骨髓瘤细胞由本实验室保存。限制性内切酶NheⅠ、HindⅢ、KpnⅠ、T4 DNA连接酶购自Toyobo公司;质粒回收试剂盒、DNA片段回收试剂盒和MTT细胞增殖试剂盒购自上海生工生物工程公司,LipoFiterTM脂质体转染试剂购自汉恒生物科技公司。胎牛血清购自浙江天杭生物公司,PRMI1640、DMEM高糖培养基购自Hyclone公司。 荧光素酶报告基因载体pGL3-Basic、pRL-TK和pGL3-Control,Dual-Luciferase Raporter Assay System购于美国Promega公司。凝胶成像系统、基因扩增仪等。
1.2 方法
1.2.1 PRDM1基因启动子片段的克隆及重组载体的构建 按Ezup柱式法从培养的293T细胞中提取基因组DNA,并检测核酸浓度及纯度。根据GeneBank(Gene ID:639)中人类PRDM1基因序列设计引物,并在引物的上游和下游分别加入NheⅠ、HindⅢ及KpnⅠ、XhoⅠ酶切位点。合成PRDM1基因的267 bp启动子和1 257 bp启动子,其中267 bp启动子的上游引物:5′-CGG CTA GCG CTA GCA ATC TGG GGG AAA G-3′;下游引物:5′-CCA AGC TTA AGC TTC TCG GCG GTC CCT C-3′;1 257 bp启动子上游引物:5′-ATG CGG TAC CCC TAC CTA TGG TAA GGC AAG CA-3′;下游引物:5′-ATC GCT CGA GTC GTT CAT CTC TAC CCA GTC C-3′;下划线为设计的酶切位点。PCR反应采用50 μL体系,反应条件为94 ℃变性5 min;循环为94 ℃ 30 s(267 bp)/94 ℃ 45 s(1 257 bp),57 ℃ 30 s(267 bp)/60 ℃ 30 s(1 257 bp),72 ℃ 30 s,共进行35个循环,最后在72 ℃延伸2 min。取2 μL PCR产物在1.5%的琼脂糖凝胶电泳分析。利用DNA片段回收试剂盒回收PCR扩增片段,回收的PCR片段经过双酶切后送至上海生工生物公司进行测序,测序正确的人PRDM1基因启动子片段连接到pTA载体中,转化感受态大肠杆菌DH5α,筛选阳性克隆,提取质粒pTA2-PRDM1,纯化并分别克隆至经双酶切的pGL3-Basic载体,以T4 DNA连接酶16 ℃连接过夜,连接产物转化感受态大肠杆菌DH5α,双酶切鉴定筛选阳性克隆,获得重组质粒pGL3-PRDM1(267 bp)/(1 257 bp),送至上海生工生物公司测序鉴定。
1.2.2 重组质粒转染及双荧光素酶报告基因检测评价启动子活性 取对数生长期293T人胚肾上皮细胞,U266和ARH77人多发性骨髓瘤细胞,以每孔5.0×105接种于12孔细胞培养板中,每组3个复孔。待细胞生长至80%左右,按脂质体LipoFiterTM试剂说明书,采用脂质体法将重组质粒pGL3-PRDM1(267 bp)/(1 257 bp)和内参质粒pRL-TK分别共转染入293T、U266及ARH77细胞,同时以pGL-Basic及pGL-SV40和pRL-TK共转染作为阴性、阳性对照,并用optiMEM 10%,DMEM完全培养基对瞬时转染的细胞进行培养,24 h后更换成DMEM+10%FBS完全培养基。按照双荧光素酶检测试剂盒提供的操作方法检测在重组细胞293T、U266、ARH77细胞中pGL3-PRDM1的荧光素酶的活性。每孔加入2 mL新鲜培养液继续培养24~48 h后,每孔细胞用PBS洗涤2次,分别加入细胞裂解液,置摇床上,室温120 r/min,振摇20 min,收集细胞裂解液。取细胞裂解液上清20 μL,加入荧光素酶检测试剂Ⅱ 100 μL,用化学发光检测仪LumunometerTD-20/20(美国Turner Bio Systems 公司产品)检测萤火虫荧光素酶活性;再加入海肾荧光素酶检测试剂(Stop Glo 试剂)100 μL,将上述反应猝灭,并启动海肾荧光素酶反应,检测海肾荧光素酶活性。荧光素酶活性用每组萤火虫荧光素酶活性与海肾荧光素酶活性的比值来表示(即启动子活性)。
1.2.3 药物筛选模型的验证 DNA甲基化抑制剂5-氮杂-2′-脱氧胞苷(5-Aza-CdR)对PRDM1基因启动子有明显的诱导激活作用[8]。因此,本研究采用5-Aza-CdR来验证该筛选方法是否可以检测PRDM1启动子激活效应,同时用RT-PCR来检测Blimp-1 mRNA的表达变化。
1.2.4 筛选中草药小分子化合物对PRDM1启动子的作用 对已有研究报道的对动物体B细胞的增殖及抗体生成有调节作用的中草药小分子提取物[9-10],如黄芪(Astragalus)、人参皂苷(ginsenoside)、白芍总苷(Total glucosides of paeony,TGP)、雷公藤多苷(Tripterygium glycoside,TWP)、灵芝多糖(Ganoderma lucidum polysaccharides,GLPS)及青蒿素的衍生物青蒿琥酯(Artesunate,ASUN)等,在构建模型的基础上,分别进行检测其对重组细胞荧光素酶基因活性及PRDM1基因启动子的影响。
1.2.5 以MTT法检测对构建药物筛选模型所需细胞系生长曲线的影响 取生长状态良好的U266细胞,以每孔5×103接种96孔板中,实验组分别加入不同浓度筛选出的能抑制PRDM1基因启动子活性的小分子化合物。每24小时进行MTT法测定560 nm处吸光度值(OD值),并连续记录6 d,以培养时间为横轴,OD值为纵轴,做不同药物对U266细胞生长曲线的影响。

2 结 果
2.1 PRDM1基因启动子序列的克隆、鉴定及表达载体的构建 从293T细胞中提取总DNA为模板PCR扩增出PRDM1基因的启动子片段,通过琼脂糖凝胶电泳成像显示在约267 bp、1 257 bp处有特异性条带,与PRDM1启动子目的片段大小一致(图1A)。纯化的PCR产物送至上海生工生物公司进行DNA测序,结果显示与GenBank中登录的PRDM1基因序列完全一致。构建的重组载体pGL3-PRDM1,PRDM1启动子267 bp、1 257 bp成功插入到pGL3-Basic载体中(图1B、C)。
2.2 选取荧光素酶表达活性更高的模型进行优化 由于细胞的共转染效率受脂质体的量、脂质体与DNA的比例,两种质粒之间的比例和宿主细胞等多种因素的影响,为了获得较强的启动子活性,对pGL3-PRDM1与pRL-TK共转染比例进行探索,设置pGL3-PRDM1与pRL-TK的转染比例为1∶1、2∶1、4∶1及8∶1分别瞬时转染293T、U266、ARH77细胞,以pGL3-Basic、pGL3-Control作为阴、阳性对照,每次实验设两复孔,并重复3次。两段不同长度启动子构建的荧光素酶报告载体pGL3-PRDM1(267 bp/1 257 bp)且与内参质粒pRL-TK在转染不同细胞株时均以2∶1共转染比例时的荧光素酶表达活性更高(图2A~C),且以1 257 bp长度启动子构建的荧光素酶报告载体转染U266细胞系荧光素酶表达活性最高(图2D),因此以pGL3-PRDM1(1 257 bp)转染U266细胞为最终选定的荧光素酶报告载体细胞筛选模型,且与内参质粒pRL-TK以2∶1的比例共转染进行后续实验。
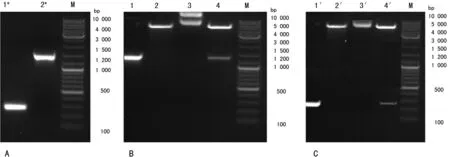
A:PRDM1基因启动子的克隆片段;B、C:构建的重组载体后经双限制性内切酶切后的片段;M:Maker DL10 000;1*:PRDM1启动子片段(267 bp);2*:PRDM1 启动子片段(1 257 bp);1:PRDM1 启动子(1 257 bp);1′:PRDM1启动子(267 bp); 2、2′:pGL3-Basic;3、3′:重组载体pGL3-PRDM1(1 257 bp/267 bp);4:1 257 bp启动子构成的重组载体酶切后pGL3-PRDM1/HindⅢ+KpnⅠ(4 800 bp+1 257 bp);4′:267 bp构成的重组载体酶切后pGL3-PRDM1/NheⅠ+HindⅢ(4 800 bp+267 bp)。
图1 PRDM1基因启动子的克隆片段及酶切片段
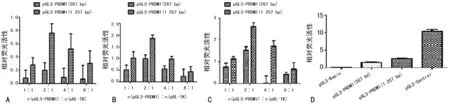
A:293T细胞;B:H929细胞;C:U266细胞;D:各组相对荧光活性。
图2 各组相对荧光活性比较
2.3 不同药物对PRDM1启动子的诱导作用
2.3.1 药物筛选模型的验证 用0.5、1、2、4 μmol/L的5-Aza-CdR处理重组的U266细胞筛选模型,与0 μmol/L 5-Aza-CdR相比,增加5-Aza-CdR 的刺激,荧光强度及PRDM1基因启动子的活性呈剂量依赖性增强,见图3。用RT-PCR检测Blimp-1 mRNA的表达水平也与之对应,随5-Aza-CdR剂量增加而表达增加,各剂量组Blimp-1基因mRNA分别为0.723±0.121、0.954±0.011、1.987±0.125、3.012±0.213、4.562±0.097,比较差异有统计学意义(P<0.05)。该筛选方法可以用于筛选针对PRDM1基因的小分子抑制剂的研究,双荧光素酶报告基因法检测PRDM1基因启动子活性具有极高的灵敏性。该方法可以采用微孔板处理细胞并直接测定荧光强度,可以高通量筛选针对Blimp-1的免疫抑制药物。
2.3.2 不同浓度青蒿琥酯和白芍总苷对PRDM1启动子活性的影响 经过反复的筛库工作及不断地重复和确定,最终得到对PRDM1启动子活性有影响的化合物,分别是对PRDM1启动子抑制作用的青蒿琥酯及对PRDM1启动子呈现低浓度增强、高浓度抑制的白芍总苷。结果显示,与0 μmol/L相比,青蒿琥酯从5 μmol/L浓度开始对PRDM1基因启动子活性呈明显降低(P<0.05),在20 μmol/L浓度时降到最低;白芍总苷呈现小剂量促进PRDM1启动子活性,高剂量抑制PRDM1基因启动子活性。见图4。
2.3.3 MTT法检测青蒿琥酯和白芍总苷对U266细胞生长增殖的影响 青蒿琥酯低剂量(≤20 μmol/L),其对细胞生长增殖影响较小,生长抑制率小于或等于10.5%;再增加剂量时,细胞毒性较明显,OD值显著小于0 μmol/L组(P<0.01),生长增殖明显受抑制。白芍总苷对细胞生长增殖的影响较复杂,≤5 μmol/L时,对细胞生长增殖的影响较小,对PRDM1启动子起增强激活的作用;≥20 μmol/L时,荧光强度明显减弱,对细胞生长增殖的抑制较明显,生长抑制率大于或等于31.3%。见表1、2。
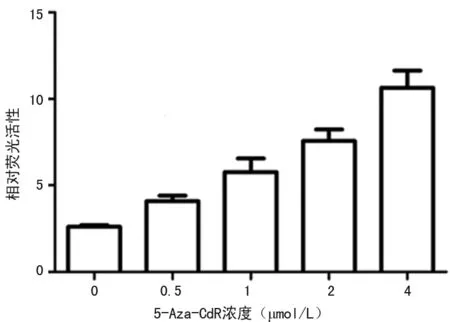
图3 在U266细胞中不同浓度的5-Aza-CdR 对PRDM1基因启动子活性的影响
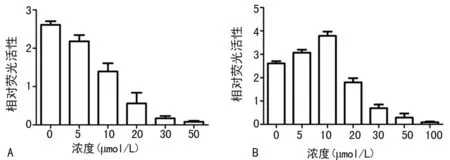
A:青蒿琥酯;B:白芍总苷。
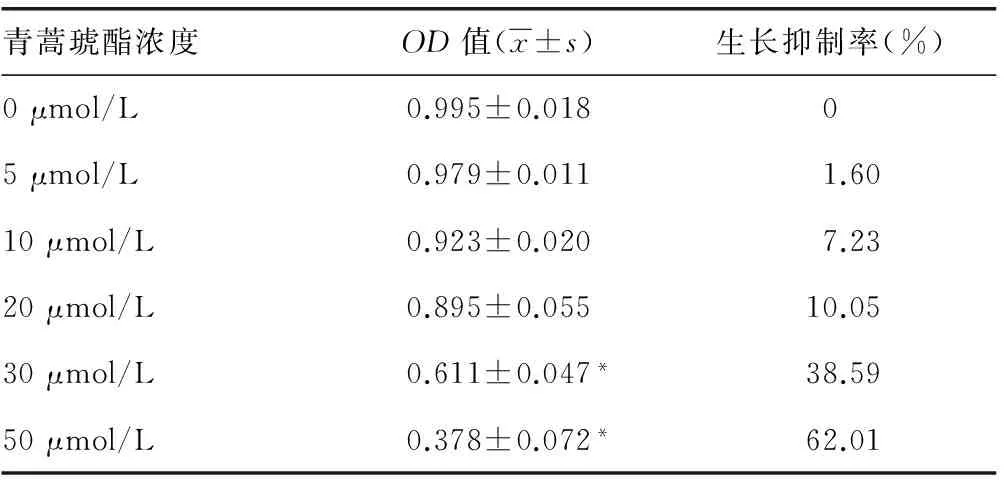
图4 不同浓度青蒿琥酯和白芍总苷对PRDM1 启动子活性影响表1 青蒿琥酯对U266细胞生长增殖的影响
*:P<0.01,与0 μmol/L比较。
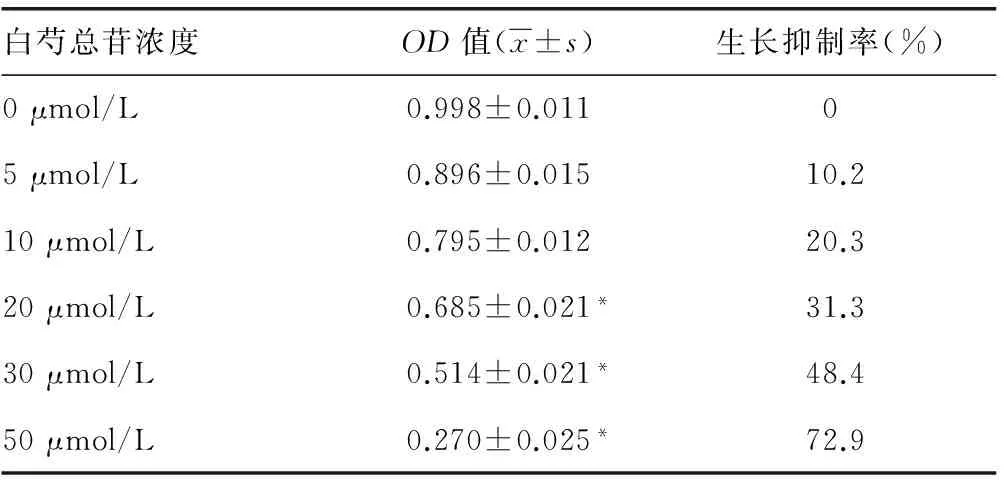
表2 白芍总苷对U266细胞增殖生长的影响
*:P<0.01,与0 μmol/L比较。
3 讨 论
Blimp-1在促进B细胞分化中起着关键作用,主要通过3个基因表达程序诱导了浆细胞发育[11-12]:(1)Blimp-1阻断B细胞增生程序,主要包括直接抑制c-myc,下调E2F-1及抗凋亡基因A1,上调cdk抑制子p18和p21;(2)Blimp-1上调促使Ig分泌的基因,包括Ig重链及轻链基因、J链、XBP-1等;(3)Blimp-1下调在形成生发中心和B细胞激活中起重要作用的基因,包括Pax-5、Bcl-6、AID、BCR信号转导相关基因等。以PRDM1基因启动子为靶点的抑制性分子将可能通过抑制浆细胞分化及抗体的产生,用于抗体所致自身免疫性疾病的治疗,并且由于成年后PRDM1的表达局限于生发中心晚期的B淋巴细胞和浆细胞[3],在其他组织器官无表达,以PRDM1为靶标的药物可能主要影响抗体的产生,而对尚存在补救途径的正常细胞免疫、固有免疫等免疫功能无太大影响,有可能成为高度特异、选择性的免疫性疾病治疗药物,为抗体所致免疫性疾病的治疗提供新的途径。
本研究通过构建重组质粒,转染细胞,双荧光素酶法检测,成功构建以PRDM1基因启动子为靶点的双荧光素酶报告基因药物筛选模型,根据信号比和灵敏度,确立了实验最佳条件:选择pGL3-PRDM1和pRL-TK的共转染比例为2∶1,以1 257 bp长度片段为活性较高启动子来构建荧光素酶报告载体,确定U266细胞为工具细胞进行后续药物筛选实验。鉴于PRDM1基因启动子CpG岛区域存在着较高程度的甲基化,使PRDM1基因表达受抑,而DNA甲基化抑制剂5-Aza-CdR可诱导PRDM1基因去甲基化再诱导转录激活[13]。因此,本实验采用5-Aza-CdR作为阳性药来验证该筛选方法是否可行,结果显示5-Aza-CdR能明显增强PRDM1基因启动子的活性,并与其mRNA的表达变化相符,说明药物筛选模型构建成功。对现已报道对动物体B细胞的增殖及抗体生成有调节作用的中草药小分子提取物如黄芪、人参皂苷、白芍总苷、雷公藤多苷、灵芝多糖及青蒿琥酯等200多种药物进行筛选试验,检测加入药物后双荧光素酶报告基因活性的表达变化,筛选出青蒿琥酯能降低荧光强度,抑制PRDM1基因启动子活性,而白芍总苷表现为低剂量促进、高剂量抑制的作用。
青蒿琥酯具有明显地减少B细胞过度活化,抑制自身抗体形成的作用,用于治疗MRL/lpr鼠后,发病延缓,发病程度减轻,血清抗dsDNA抗体和抗ANA抗体滴度降低,小鼠脾脏中BAFF mRNA表达水平显著下降,其机制可能是参与体内B细胞分化调控的关键环节[14]。但其是否参与调节PRDM1基因的表达及调节的分子机制尚不明了,其对PRDM1基因的下调作用是初始靶点还是其某个上游基因的后继效应呢?为此,本实验在筛选出最佳条件的作用下,加入不同浓度的青蒿琥酯处理,观察青蒿琥酯是否直接作用于PRDM1基因启动子,结果表明青蒿琥酯呈剂量依赖性抑制PRDM1基因启动子活性,并结合细胞的生长增殖曲线,≤20 μmol/L浓度时对PRDM1基因启动子活性的抑制程度较高且细胞生长状态影响较小,随着浓度的加大细胞生长增殖明显受抑制,可能与青蒿琥酯还影响细胞生长增殖的调控有关;白芍总苷也是传统的免疫调节药物,既能调节T细胞免疫,又能调节B细胞免疫,体外实验证实白芍总苷具有低浓度促进和高浓度抑制的双向调节作用,通过多种途径抑制自身免疫反应,对类风湿关节炎、系统性红斑狼疮等自身免疫性疾病有确切疗效,其作用机制可能是通过负调节腹腔巨噬细胞(PM)产生IL-1及产生大量PGE2而抑制B细胞增殖分化[15]。因此本实验也检测了白芍总苷是否抑制PRDM1基因的表达,结果显示小剂量可促进PRDM1基因表达,大剂量会抑制PRDM1基因启动子活性,而大剂量时对细胞生长增殖的影响较大,说明白芍总苷抑制PRDM1启动子活性可能与细胞毒性有关。
综上所述,本实验构建了重组质粒pGL3-PRDM1,将其与内参质粒pRL-TK瞬时共转染工具细胞,采用双荧光素酶法检测基因启动子活性,建立了以PRDM1基因启动子为靶点的药物筛选模型,为筛选以PRDM1基因为靶点的免疫抑制药物提供了一种有效的方法,同时也丰富和发展了中草药小分子提取物在治疗自身免疫性疾病方面的机制研究,为研发新的免疫抑制剂奠定基础。
[1]Morgan MA,Magnusdottir E,Kuo TC,et al.Blimp-1/Prdml alternative promoter usage during mouse development and plasma cell differentiation[J].Mol Cell Biol,2009,29(21):5813-5827.
[2]Györy I,Fejér G,Ghosh N,et al.Identification of a functionally impaired positive regulatory domain Ⅰ binding factor 1 transcription repressor in myeloma cell lines[J].J Immunol,2003,170(6):3125-3133.
[3]Shaffer AL,Lin KI,Kuo TC,et al.Blimp-1 orchestrates plasma cell differentiation by extinguishing the mature B cell gene expression program[J].Immunity,2002,17(1):51-62.
[4]Garaud JC,Schickel JN,Blaison G,et al.B cell signature during inactive systemic lupus is heterogeneous:toward a biological dissection of lupus[J].PLoS One,2011,6(8):e23900.
[5]Raychaudhuri S,Thomson BP,Remmers EF,et al.Genetic variants at CD28,PRDMl and CD2/CD58 are associated with rheumatoid arthritis risk[J].Nat Genet,2009,41(12):1313-1318.
[6]王爱萍,徐今宁.中药免疫调节作用研究进展[J].中国药业,2011,20(3):75-77.
[7]Mundy G,Garrett R,Harris S,et al.Stimulation of bone formation in vitro and in rodents by statins[J].Science,1999,286(5446):1946-1949.
[8]Patra SK,Bettuzzi S.Epigenetic DNA-(cytosine-5-carbon) modifications:5-aza-2′-deoxycytidine and DNA-demethylation[J].Biochemistry (Mosc),2009,74(6):613-619.
[9]储岳峰,李祥瑞,胡元亮.9种中药成分对小鼠免疫细胞活性的影响[J].南京农业大学学报,2004,27(1):97-100.
[10]Shan BE,Yoshida Y,Sugiura T,et al.Stimulating activity of Chinese medicinal herbs on human lymphocytes in vitro[J].Int J Immunopharmacol,1999,21(3):149-159.
[11]Boulet S,Daudelin JF,Labrecque N.IL-2 induction of Blimp-1 1 is a key in vivo signal for CD8+short-lived effector T cell differentiation[J].J Immunol,2014,193(4):1847-1854.
[12] Jackson DA,Elsawa SF.Factors regulating immunoglobulin production by normal and disease associated plasma cells[J].Biomolecules,2015,5(1):20-40.
[13] Watanabe Y,Toyota M,Kondo Y,et al.PRDM5 identified as a target of epigenetic silencing in colorectal and gastric cancer[J].Clin Cancer Res,2007,13(16):4786-4794.
[14]金鸥阳,张华勇,徐婷,等.青蒿琥酯治疗MRL/lpr狼疮鼠肾炎的病理变化及机制[J].实用临床医药杂志,2007,11(7):5-9.
[15]马丽,李作孝.白芍总苷的免疫调节功能及其临床应用[J].中国实验方剂学杂志,2010,16(17):244-246.
《重庆医学》杂志对运用统计学方法的有关要求
1.统计学符号:按GB 3358-1982《统计学名词及符号》的有关规定,统计学符号一律采用斜体。
2.研究设计:应告知研究设计的名称和主要方法。如调查设计(分为前瞻性、回顾性或是横断面调查研究),实验设计(应告知具体的设计类型,如自身配对设计、成组设计、交叉设计、析因设计、正交设计等),临床试验设计(应告知属于第几期临床试验,采用了何种盲法措施等);主要做法应围绕4个基本原则(重复、随机、对照、均衡)概要说明,尤其要告知如何控制重要非试验因素的干扰和影响。
4.统计学分析方法的选择:对于定量资料,应根据所采用的设`计类型、资料所具备的条件和分析目的,选用合适的统计学分析方法,不应盲目套用t检验和单因素方差分析;对于定性资料,应根据所采用的设计类型、定性变量的性质和频数所具备的条件及分析目的,选用合适的统计学分析方法,不应盲目套用χ2检验。对于回归分析,应结合专业知识和散点图,选用合适的回归类型,不应盲目套用简单直线回归分析;对于具有重复实验数据检验回归分析资料,不应简单化处理;对于多因素、多指标资料,要在一元分析的基础上,尽可能运用多元统计分析方法,以便对因素之间的交互作用和多指标之间的内在联系做出全面、合理的解释和评价。
5.统计结果的解释和表达:应写明采用统计学方法的具体名称(如:成组设计资料的t检验、两因素析因设计资料的方差分析、多个均数之间两两比较的q检验等),统计量的具体质(如:t=3.45,χ2=4.68,F=6.79等);在用不等式表示P值的情况下,一般情况下选用P>0.05、P<0.05和P<0.01 3种表达方式,无须再细分为P<0.001或P<0.000 1。当涉及总体参数(如总体均数、总体率)时,再给出显著性检验结果的同时,应再给出95%可信区间(CI)。
Establishment of drug screening model targeting PRDM1 gene promoter*
HuWentan1,WangTingting1,ZhangShijie1,HeXin2,ZhangJingyu1,LiuHongchun1△
(1.DepartmentofClinicalLaboratory,theFirstAffiliatedHospitalofZhengzhouUniversity,Zhengzhou,Henan450003,China;2.DepartmentofClinicalLaboratory,HenanCollegeofChineseMedicine,Zhengzhou,Henan450002,China)
Objective To construct a double luciferase reporter gene vector with PRDM1 gene promoter as target,and establish drug screening cell model in vitro,hope to find small molecule compounds in Chinese herbal medicine library by this model.Methods Total DNA was extracted from 293T cells and it was used for amplifying the fragment contained PRDM1 gene promoter(267 bp/1 257 bp) by PCR.The amplified product was inserted into pGL3-Basic vector.The PCR product and pGL3-PRDM1 vector were verified by sequencing and alignment.The pGL3-PRDM1 and pRL-TK vector were co-transfected into engineer cells.The activity of PRDMl gene promoter could be assayed by measuring luciferase.The method was optimized by changing ratio of two vectors.Results The highest transfection efficiency and luciferase activity were found in ratio of n(pGL-PRDM1)∶n(pRL-TK)=2∶1,and with the recombinant luciferase report gene vector contained the length of 1 257 bp amplified fragment transfecting into U266 cells.Moreover,the inductor (5-Aza-CdR) of PRDM1 gene was used for authenticating the method(P<0.01),the fluoresscence in tensity and promoter activity of PRDM1 gene were enhanced in a dose dependent manner with 5-Aza-CdR.The activity of the promoter of PRDM1 gene was significantly decreased from the concentration of 5 μmol/L of Artemisinid(P<0.05). The total glucosides of paeoniflorin promoted the promoter activity of PRDM1 gene at a low concentration, and inhibited the promoter activity of PRDM1 gene at a high concentration. Artesunate has no effect on cell proliferation. The effect of total glucosides of paeony on cell proliferation was more complicated.Conclusion A drug selection model targeting PRDM1 gene promoter has been successfully established,and artesunate has been screened to inhibit the promoter activity of PRDM1 gene.
PRDM1 gene;dual-luciferase report gene;drugs,Chinese herbal;drug screening
10.3969/j.issn.1671-8348.2017.11.002
河南省卫生厅计划指导项目(201304015);河南省教育厅科技研究重点项目(14A320026);郑州市科技局资助项目(141PPTGG437)。 作者简介:胡文坛(1990-),检验技师,在读硕士,主要从事自身免疫性疾病的发病机制及分子诊断的研究。△
,E-mail:xingyunerliu@163.com。
R965.1
A
1671-8348(2017)11-1445-05
2016-11-03
2017-01-19)
