基于CNKI的分子生物学技术在湖北省林业研究中的应用与进展分析
2017-04-18彭婵杨杉陈慧玲张新叶��
彭婵++杨杉++陈慧玲++张新叶��
摘要:以CNKI中国知网数据库为文献数据源,采用文献调研的方法分析近 10年来分子生物学技术在湖北省林业研究方面的报道,结果表明:分子标记技术和基因工程技术是湖北省林木分子生物学研究的主要技术,分子标记的开发与体系构建、遗传基础及多样性评价、功能基因克隆与转化是研究的热点领域。最后,结合湖北省林业现状和分子生物技术的发展趋势对以后的研究提出了意见和建议。
关键词:分子生物学;林业研究;湖北
中图分类号:S718文献标识码:A文章编号:1004-3020(2016)06-0001-09
Application Progress on Molecular Biotechnologyof Forestry Research in Hubei Province:Based on CNKI
Peng ChanYang ShanChen HuilingZhang Xinye
(Hubei Academy of ForestryWuhan430075)
Abstract: The data was collected from CNKI,the literature research method was used to retrieve all the papers about the recent process in molecular biotechnology of forestry research in Hubei Province,The results showed that:molecular marker technology and gene engineering technology are the key techniques on molecular biotechnology research in Hubei forestry.Development of Molecular Markers and System Construction ,Genetic Basis and genetic diversity evaluation, cloning and transformation of functional gene are three new research hot topics.Finally Some suggestions for future resarch on molecular biotechnology research in Hubei forestry were put forward.
Key words:Molecular biotechnology;Forestry research;Hubei
分子生物学是近半个世纪以来迅猛发展的前沿学科,该学科以研究遗传信息(核酸)及其载体(蛋白质)等生物大分子的组成结构、生物功能为主要目标,揭示各种生物大分子在生物特征和生命活动中的相互作用关系,并在分子水平对其进行利用和改造,因其具有简便高效、低耗低害等特色已被各国广泛应用于医药、卫生、食品、化工、环保、能源、司法等领域。
林业研究与其它行业相比具有其特殊性。首先林木普遍的生长周期长,几年到几十年不等,见效慢;其次林木基因组大(挪威云杉基因组是人类基因组的七倍,是拟南芥的一百多倍)[1],且林木基因组杂合度高遗传背景复杂;另一方面许多林木树种的表型特征必须在达到其特定的成熟期才能显现出来;最后是林木本身体积大,生长面积和分布范围也大,实地调查费时费力。林业研究的特殊性决定了常规研究方法很难实现短时间内提升林木生产应用价值、达到林木速生高抗多性状综合改良需求,而分子生物学技术从分子水平研究林木生长成才机理,完全突破了传统研究思维的限制,是林业发展的全新研究方向。湖北省森林覆盖率达3961%,林木种质资源丰富,为了解分子生物学技术在湖北省林业研究的应用情况,本文从中国知网全文数据库资源调查入手,检索了近10年来有关湖北分子生物学在林木研究上的文献资料,系统归纳和梳理分子生物技术在湖北林业研究中的应用历程,了解研究现状以及当前学科发展态势,为分子生物技术在湖北林业研究中发挥更大作用提供理论依据。
1材料与方法
以中国知网(http://www.cnki.net/) “中国学术期刊网络出版总库”、“中国博士学位论文全文数据库”和“中国优秀硕士学位论文全文数据库”为数据源,检索日期为200611至20161121日,采用标准检索方式,根据林业学科涉及范围,学科领域选择 “农业科技”中“林业”子学科,“基础学科”中“生物学”下的“植物学”子学科,以“分子”“DNA”或“基因”为主题关键词,以湖北省内科研单位及大专院校为第一作者单位,剔除综述、评论、文库重复等共检索出文献105篇,其中包括46篇硕博论文,59篇学术期刊论文。
湖北林业科技第45卷
第6期彭 婵,等:基于CNKI的分子生物学技术在湖北省林业研究中的应用与进展分析
2结果与分析
2.1文献发布树种及技术类型
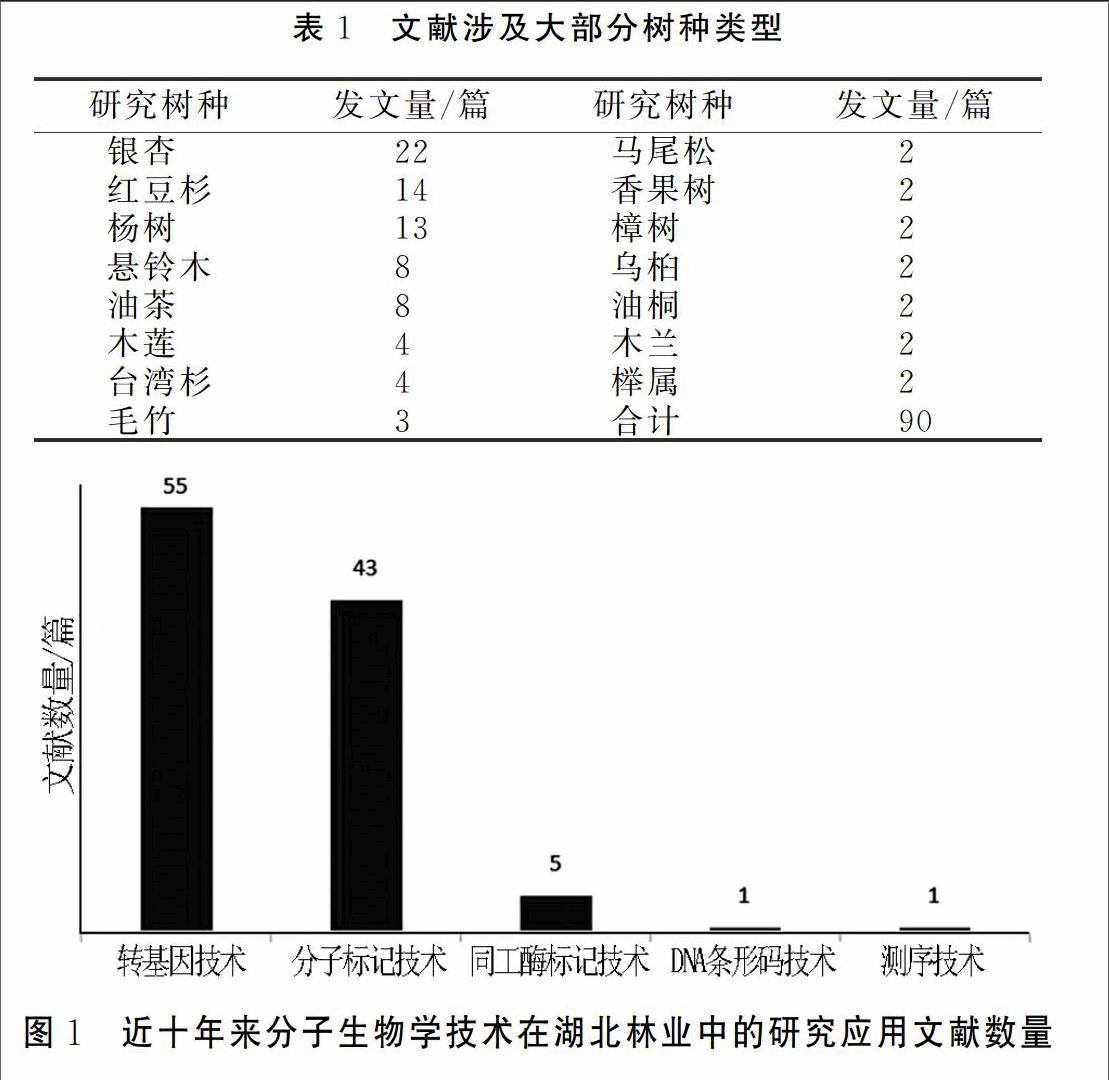
对105篇文献进行统计分析,涉及到树种30类,大部分树种如表1所示,除表1另有珙桐、栓皮栎、白皮松、刺槐、水杉、杜仲、枫杨等15余种树木。研究文献所涉及主要技术手段:其中55篇文献为基因工程转基因技术相关文献,43篇文献以分子标记筛选与应用为主要内容,DNA 条形码技术、测序技术文献各一篇,同工酶技术文献5篇(见图1)。
2.2分子生物学技术在湖北林业研究中的应用
2.2.1同工酶技术的应用
同工酶(等位酶)技術就是通过对同种酶不同分子形式的电泳谱带分析,来识别控制这些谱带的基因位点和等位基因,同工酶技术与DNA标记技术相比存在信息量少、且无法从DNA水平上检测遗传变异等缺点, 但是在早期林木研究中曾被广泛应用。
李军民[2]、丁小飞等人利用同功酶(等位酶)技术对湖北省部分地区的栓皮栎[2]、檫木[3]、马尾松[4]和白皮松[5]天然群体进行了遗传结构和遗传多样性分析,结果表明:长阳栓皮栎群体和马尾松群体都具有较高的遗传变异性水平,而檫木及南漳白皮松群体遗传变异性水平均低于其他同类平均水平,同时结果发现马尾松群体和檫木群体内杂合体不足,纯合体过量,处于非平衡状态,故对其适宜采取原地保护和异地保存相结合的保护策略。
丁小飞[6]等人利用等位酶分析结合外部形态(叶片、树皮、小枝)比较,从表现以及基因型鉴别了鹅掌楸属内 2 个种及种间杂种,并发现在Pgm和Shdh的基因位点分别存在特征谱带,可以用于今后从基因水平识别鹅掌楸开杂交育种和杂种苗。
2.2.2分子标记技术的应用
随着分子生物学及其技术的快速发展,分子标记技术因其能直接从DNA 水平反映个体遗传差异且不受植株发育时空和环境及其他因素影响所以得到了非常广泛的应用,是现今最为理想的遗传标记技术。近年来湖北省与林木相关的许多新分子标记被开发,并在种质资源种群结构、遗传变异、亲缘关系鉴定与分类、系统进化、以及育种等领域发挥重要作用。
(1)遗传多样性的分析。遗传多样性是生物所携带的遗传信息总和,一个群体的遗传多样性越高,其对环境的适应能力越强,越容易拓展分布范围,反之就越容易濒临灭绝。因而遗传多样性的分析测定对了解物种的遗传基础、环境适应性、防止资源过度开发和制定科学有效的保护策略具有举足轻重的意义。在湖北省林木资源遗传多样性研究应用领域,主要涉及以下4 个方面:①运用分子标记技术对同一树种不同群体的遗传基础进行评价。大量研究表明,湖北林木种质资源具有丰富的遗传多样性。研究人员通过AFLP[7]、SRAP[8]以及ISSR+SRAP[9]的标记组合分析了湖北省内各地区大面积栽培的油茶群体,结果显示均具有较高的遗傳多样性。在优良单株研究上彭婵[10]、李良[11]等分别对湖北省林科院收集的鄂、湘林、长林等市场优良品种和全省范围内初选优良单株进行了SSR和ISSR分析显示多样性丰富。郭新安[12]、赵西梅[13]等人分别对红安、罗田、麻城、巴东、随州和安陆等地银杏古树进行ISSR标记研究,结果一致发现在物种水平上这些地区的银杏古树群体具有较高的遗传多样性,结合形态标记和分子标记对部分优良单株的遗传基础评价也认为总体水平上遗传基础较宽[14]。②运用分子标记技术探究物种濒危原因并提出科学保护措施。杨琴军[1516]、李江伟[1718]等在建立稳定的台湾杉RAPD、ISSR、NSSR和CPSSR反应体系基础上对包括湖北星斗山在内几个台湾杉原生居群的研究结果显示各地种源遗传多样性高低程度不一,居群间的地理距离与遗传距离之间无明显相关性,认为台湾杉生长原生境直接破坏及其自身的自然更新不良是导致台湾杉种群濒危的可能原因。珙桐[19]、香果树[20]也存在同类问题,对此类树种建议:建立髙效的种子萌发技术体系,保护现有资源禁止砍伐,种植园内迁移保存。而白豆杉[21]与黄梅秤锤树[22]群体则面临近交衰退风险,亟待开展迁地保护工作, 包括构建资源库保存关键基因型。武汉植物园邓建云[23]研究古树资源发现一株有特异AFLP标记的杜仲古树,并推测这一杜仲古树是原始野生植株,在现今野生杜仲资源几乎灭绝的情况下这株古树急需进行切实的保护和深入研究。③运用分子标记技术对品种遗传稳定性进行评价。物种遗传稳定性关系到林木种质保护,利用分子标记进行遗传稳定性分析尤为重要,可以解决林木经过多代、多次无性繁殖后种苗是否依旧保有其原品种的遗传性状,以及林木种苗对外界环境干扰的变异性等问题。熊丹[24]等对野生香果树体细胞胚繁殖中不同继代的胚性愈伤组织和再生植株之间的遗传稳定性进行RAPD标记研究, 发现香果树的胚性愈伤组织会发生体细胞无性系变异,但随着继代次数的增多,体细胞胚的变异率不是逐渐升高,而是先升高后下降。而姚军[25]等选取处于不同立地条件下少果毛一球悬铃木不同年份的嫁接单株进行ISSR 分子标记研究显示,一球悬铃木嫁接单株之间遗传差异极小,遗传稳定性高,多年嫁接后仍与母本性状保持一致。王瑞静[26]采用不同剂量60Co-γ对黑派4个杨树品种辐射敏感度进行研究,发现各剂量组间平均基因相似系数变化无规律,并未因辐射剂量增大而增大,辐照处理后获得的杨树植株在DNA水平上存在多态性差异,圣山杨对辐射较为敏感。④运用分子标记技术对各个群体进行聚类分析,探讨不同群体的亲缘关系与地理位置关系。各树种遗传多样性的普遍研究[2730]结果显示相同区或地域邻近的居群遗传距离也相对较小、亲缘关系较近;地理距离较远的居群,其遗传距离也相对较大、亲缘关系较远,说明各群体之间亲缘关系的亲疏与地理分布的远近呈较强的正相关性。但木荷[31]个别无性系与相同产地无性系并未聚在起,推测其遗传基础的变化可能是人为因素的干扰所致。复杂的山形地貌的阻隔也可能对同一地区不同居群间基因流造成影响,袁珊[32]对神农架4 个居群66 个连香个体调查发现,在小尺度的山体隔离下也可能产生物种不同程度的分化,从而影响植物居群的遗传结构。
(2)核心种质构建。不断深入的林木研究工作需要广泛收集种质资源和保存丰富的遗传资料,但随着种质资源的不断积累,尤其是多数林木自身体积量大,其种植生长面积需求量更大,庞大的资源群体数量给种质资源的保存带来了压力。为解决这一难题,并提高有限空间里种质资源研究和利用的效率,以尽可能小数量的收集品种代表尽可能多的物种遗传性状,核心种质的构建成为现今林业研究的一个重要方向。
文靓[33]在ISSR分子标记结果的基础上,采取逐步聚类随机取样法分别湖北乡土毛白杨、响叶杨、山杨和大叶杨种质资源构建核心种质,t检验结果各项遗传指标与初始种质差异不显著,说明构建的核心种质能够很好代表原始种质的遗传多样性。郑瑜[34]研究了银杏核心种质构建的最佳构建策略,认为分组应按来源产地,组内以对数比例法确定取样量,25%的取样比例,最后以组内类平均法(UPGMA)聚类抽样的策略最佳。并以此法筛选出52份核心种质样品,各项遗传多样性参数明显高于保留种质相比,说明所构建的核心种质对原始种质群体遗传多样性具有较高的代表性。
(3)种质鉴定。优良的种质资源是林木研究、生产和育种的基础,林木品种鉴定是确保优良种源的一个重要环节。传统的植株形态鉴定虽然简单直观,但也受限于表观特征数量少、环境影响大、主观因素干扰、植株生长周期季节限制等因素,难以正确反映品种之间固有的亲缘关系。而采用分子标记技术进行杂种纯度鉴定,是从基因组DNA的水平检测杂交种及其亲本的差异,鉴定速度快、准确率高、非生物因素影响小,确保了被鉴定品种的真实性。杨树是中国工业人工林主栽树种之一,目前各地栽培的杨树品种繁多且混杂,但杨树种苗形态上可作为品种鉴定的特征标记数量有限,尤其是亲缘关系相近的杨树黑杨品种间的形态学差异甚微,张亚东[35]等人利用新型分子标记EST-SSR鉴定湖北省内目前主栽的 8 个黑杨品种,得到7 对多态性引物在不同品种间有特异谱带,而只需4对就可以完全区别这8个近缘关系种,该结果证明EST-SSR标记完全可以应用于鉴定林木品种,且鉴别效率较高。付秀芹[36]联合形态学特征、光合作用特点并RAPD分子标记鉴定了乌桕杂种的真实性,进一步证实了分子标记完全可以应用于林业实践生产中。
(4)分子系统学研究。系统学是运用DN A 分析技术从基因、个体及居群水平上,来探讨物种的多样性、亲缘关系及其之间进化关系的科学,可了解物种的起源与传播途径、系统位置和分类地位等,利于资源保护保存和可持续利用[3738]。现在许多林木的系统发育关系至今仍不清楚,分类和系统发育关系以及起源问题,还没有达成共识。王丽娜[37]、王璐[39]等分别对山毛榉科水青冈属7个物种以及部分外类群进行了系统发育关系和地理分布格局研究,前者结果认同泛生物地理学的研究,即水青冈属的分子系统发育与之一致,都认为水青冈属植物在进化过程中受大陆漂移等地质历史事件的影响。后者研究结果也与榉属地理分布极其吻合,因此假設榉属的原始分布区或起源中心为北太平洋的某个地区(中国或日本北部)。陈仁芳[38]综合分子系统学和其他三门科学研究桑属属内种数和系统发育关系,对桑种植资源鉴定、育种亲本选择以及桑的分子育种等技术问题有很大帮助。
2.2.3其他技术应用
DNA条形码技术是建立在DNA序列分析技术之上,即用一个或几个标准的DNA片段来完成物种分类和鉴定工作,利用该方法进行木材分类是传统木材鉴别方法的重要突破,湖北大学杨路路[40]以水杉为例,探寻了保存1 a、2 a和6 a的水杉木材及阴沉木DNA提取方法,并用DNA条形码技术筛选分析DNA序列片段,证实了序列ITS-2、trnL-F均适合杉科DNA条形码技术要求,对于已降解材料ITS-2更为适用,继而在此基础上建立了一套适于杉科分子水平的木材鉴定体系.
表达序列标签(EST)和基因组测序也是功能基因组学中常用的高通量研究方法,湖北省林科院张新叶[4142]等人首先利用现有数据库中来自美洲黑杨及欧美杨的20 023条EST序列开发了杨树EST-SSR标记,此后又结合EST序列和生物信息学方法进行了杨树SNPs标记开发,并测序验证得到13个候选SNPs位点可以用于杨树进一步相关研究。利用所开发的标记,杨彦伶[43]等人基于柳树和杨树的亲缘关系,探究了杨树SSR和ESTSSR标记在柳树中的通用性,发现ESTSSR标记通用性较好,而基因组SSR标记多态性较好。此后,张新叶[44]等人又用黑斑病高抗与高感的美洲黑杨为材料建立了2个cDNA文库,通过克隆测序鉴定出177个抗病候选基因,该研究为进一步从基因水平探究黒杨的抗黑斑病机制及功能基因的发现奠定了基础。
利用EST技术开发的分子标记来自于基因编码序列,因其更为保守多态性相对量也较少,因此开发全基因组标记对林木研究也不可或缺。目前,第 2 代高通量测序技术已被广泛应用,水杉是中国一级保护植物,发源地湖北利川,张新叶[45]利用第二代高通量测序平台 ROCHE-454 GLX对水杉基因组进行测序,在得到的全基因组信息上对所含微卫星重复序列进行了特征和组成情况分析,发现1 965个微卫星其中921个可开发的SSR标记位点,引物验证结果也表明所开发标记的高效性,该结果不仅丰富了水杉群体的标记资源,更对保存遗传学与分子标记辅助育种有重要意义。
2.2.4林木基因工程研究
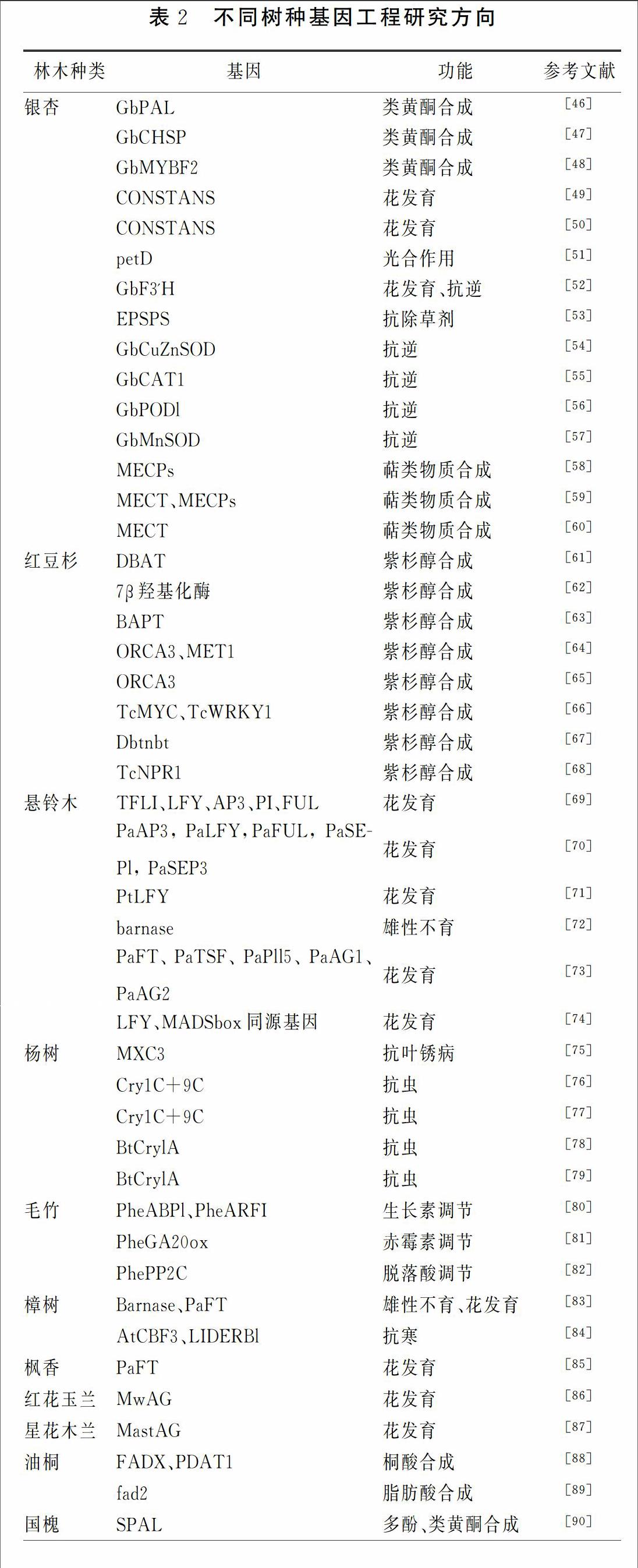
通常林木基因工程是指将克隆的外源优良目的基因利用DNA重组等技术导入植物细胞或组织进行表达,以有效改良品种的不良性状定向获得有利性状的植物新品种的技术,该技术不仅克服了植物有性杂交的限制性而且无限扩大基因交流的范围。在过去的 10年里,湖北省林木基因工程已在花期调控、雄性不育、次生代谢物合成、抗病虫害、和抗逆等发面取得了很大进展,详情见表2。
(1)银杏基因工程进展。银杏叶中活性化合物不仅对植物自身生长和抗逆性有重要作用,也是植物药制剂的重要材料,我省银杏基因工程研究主要围绕银杏叶黄酮类和萜内酯类合成代谢调控以及银杏逆境胁迫耐受力展开。
李金宝[46]、李琳玲[47]从等结构基因表达序列方向,分别对银杏叶中类黄酮生物合成过程中的关键酶PAL(苯丙氨酸解氨酶)、CHSP(查尔酮合成酶)基因启动子序列进行了表达载体构建和功能分析。袁红慧[59]人等通过基因步移法克隆得到萜内酯合成 MEP 途径中两个关键酶基因GbMECT和 GbMECPs的启动子序列,并对其进行了生物信息学分析,为后续萜内酯或其前体物质生产建立了分子基础。
李琳玲[54]、程华[5557]等人首次利用RACE技术克隆得到银杏叶绿体铜锌超氧化物歧化酶基因GbCuZnSOD、锰型超氧化物歧化酶GbMnSOD、过氧化氢酶基因CAT1、POD1的cDNA序列,采用Northern杂交及进化树分析基因起源,并用RTPCR技术探明这些基因在植物中的表达情况,以期揭示其对环境适应能力及抗逆性分子机理,为林木遗传改良提供理论依据和基因资源。
(2)红豆杉基因工程。
紫杉醇是红豆杉的重要次生代谢产物,已有研究证明紫杉醇能够使细胞周期停滞于有丝分裂期从而抑制癌细胞的有丝分裂,对各种癌症都有很好的疗效[64,67]。研究其在红豆杉中生物合成代谢及机制显得尤为重要。
紫杉醇合成约需要 20 步酶促反应,探究参与各合成反应的酶及其编码基因的功能, 并通过基因工程途径调节紫杉醇的合成代谢是目前的一个主要研究方向。燕丽娜[62]等以南方红豆杉为材料, 获得了紫杉烷7β羟基化酶基因的全长序列,并对其进行了进化分析。张志建[63]等对曼地亚红豆杉的3氨基3苯基丙酰转移酶(BAPT)基因进行了克隆,并研究其在大肠杆菌中的表达。谢莎[68]从中国红豆杉细胞中克隆TcNPR1基因,研究了其响应SA诱导大量合成紫杉醇的途径和分子进化情况。张鹏[67]克隆了紫杉醇合成的最后一个关键酶紫杉烷侧链苯甲酰转移酶基因Dbtnbt,转化红豆杉细胞过表达后发现紫杉醇产量提高约37%。
紫杉醇合成系列关键酶基因的表达及其活性受转录因子影响,通过改造关键酶基因的启动子或转入相应的转录因子可激活酶的基因表达。邓慧[61]克隆曼地亚红豆杉细胞紫杉醇合成途径中关键酶基因 DBAT 启动子,为后续克隆相应紫杉醇转录因子打下基础。于小青[64]将转录因子ORCA3和MET1RNA分别导入到红豆杉细胞中发现紫杉醇产量明显升高。李书涛[66]使用酵母单杂交技术分离了转录因子TcMYC和TcWRKY1,下一步是分析其在SA和 MeJA 介导紫杉醇合成过程中起到的作用。
(3)悬铃木基因工程。悬铃木是湖北省重要的园林绿化树种,华中农业大学围绕定向培育无毛或者雄性不育新品目标进行了一系列相关研究。黄文俊[72]将导致雄性不育的核糖核酸酶基因barnase导入到悬铃木得到5%的稳定转化率。李志能[74]构建了悬铃木雌花cDNA文库,并结合同源序列法和RACE技术克隆了花发育LFY基因和部分MADSbox基因,为下一步不育悬铃木研究奠定了基础。易双双[70],张佳琪[73]分别通过抑制成花转换、花器官形成等关键基因PaFT、PaLFY的表达,以及时空表达分析以及异源表达实验来研究这些关键基因的生物学功能,并通过表型观测验证了部分开花相关基因转化遗传稳定性。孔宇飞[69]在前人基础上利用GUS报告基因对影响以悬铃木合子胚为受体的农杆菌介导遗传转化条件进行探索认为Tm的浓度为300 mg/L,最适合作为转化体系的抑菌浓度。菌液浓度OD值0.6~0.8之间,农杆菌的侵染能力最强,农杆菌EHA105适合作为侵染细菌。
围绕无果、雄性不育的新品种培育方向,施雪萍[83]针对樟树在成功转化GFP基因植株基础上,又进行了Barnase雄性不育基因和PaFT早花基因的转化并得到了转化Barnase的植株,证明了樟树可以通过基因工程进行遗传改良。其后刘冉冉[85]也将悬铃木PaFT基因转入枫香叶片再生体系开拓了乡土树种枫推广应用新方向。
(4)杨树基因工程。病虫害一直是侵害林木生长的重要因素,林木抗性研究即通过基因工程手段引入抗病虫害基因,使其在植株内稳定地遗传和表达获得相应抗性。湖北省气候温暖湿润也是各种病害高发区,杨树是主要造林以及林业产业树种,熊瑞婷[75]以杨树抗锈病基因MXC3为对象构建RNAi载体,以南林895杨的组培苗叶片作为受体材料进行遗传转化发现外源基因影响了杨树内源MXC3基因的表达;徐金艳[76]、卢小三[77]分别构建了SN0431、LH0143、LH0417杨树无性系再生体系并探究了其在不同条件下对转化抗虫基因CrylC+9C转化效率的影响。康薇[78]、郑进[79]等针对中嘉8号杨存在抗虫性不强的缺陷,建立了中嘉8号杨原生质体再生及转化体系,并发现转BtcrylA基因杨树植株具有明显的抗虫活性。肖硕[91]首次将BtcrylA基因转入刺槐,并建立和优化了农杆菌介导的刺槐离体茎段的转化体系,但对转化植株在田间通过抗性试验有待更进一步的检验和筛选。
(5)其他树种相关基因工程。张艳[80]、梁晓静[81]、郭少玲[82]利用RACE技術分别从毛竹中克隆出与生长素、赤霉素、脱落酸相关基因PheABPl+PheARFI、PheGA20ox、PhePP2C,并用QRTPCR技术分析了这些基因在不同剂量的60Coγ射线处理中的相对表达量。三峡大学景丹龙[86]、张博[87]分别对木兰科红花玉兰和星花木兰花发育AGAMOUS(AG)同源基因MwAG和MastAG的结构和功能进行分析,认为MwAG基因是雌、雄蕊发育的关键基因,仅表达于雌、雄蕊中而不在幼叶和花被片中,基因表达量与其雌、雄蕊发育快慢呈强相关性,而MastAG基因参与心皮的发生,能抑制A类基因活性。任景[89]利用SNP技术对控制油桐脂肪酸代谢关键基因fad2基因的变异情况进行研究认为该基因编码区的SNP是导致不饱和脂肪酸和桐酸含量的增加的主要原因。许锋[90]等利用反义RNA技术克隆国槐苯丙氨酸解氨酶基因SPAL并构建反义表达载体,导入拟南芥后对获得的抗性植株进行Northern杂交鉴定及成分含量分析,结果表明SPAL基因对多酚和类黄酮有明显抑制作用。湖北大学田良涛[92]首次构建了乌桕三倍体培育体系并进一步研究了乌桕胚乳转基因的方法,为提高乌桕产能利用率打下基础。
3结论与讨论
本研究以CNKI中国知网全文期刊数据库和硕博士学位论文数据库为数据源,检索和分析2006~2016年分子生物学在湖北林业研究中的应用情况,发现其中分子标记技术和转基因技术是林业研究重点,主要涉及三大方面: 分子标记的开发与体系构建、遗传基础及多样性评价、功能基因克隆与转化,而有关核心种质构建、DNA条形码、cDNA文库建立、基因组测序等研究成为新兴的热点研究方向。虽然分子生物技术已经在湖北林业研究的各个领域中得到广泛应用, 但与其它行业相比, 无论是在技术成熟度还是应用范围来讲都是刚刚起步。根据湖北林业研究的可持续发展的需要,结合分子生物技术的学科特性, 建议未来湖北林木分子研究从以下几个方面着手开展研究工作:
(1)充分利用省内资源优势, 加强本土优良、珍贵林木种质资源发掘与收集,同时着力开展已有资源高产、优质、多抗等相关性状的系统评价, 储备优质遗传资源。
(2)不断改进和利用新的分子标记技术,在进行新性状 DNA 分子标记筛选的同时,积极寻找与林木重要性状的主效基因连锁更为紧密的标记,实现功能基因快速准确追踪。
(3)构建高密度遗传连锁图、物理图谱寻求更为快速、简捷的克隆功能基因的方法。
摸索建立可操作性强的转基因体系,对性状改良基因进行重点研究,改善一些具经济价值性状如观赏、食用、药用、环保、工业用等以获得更大的经济价值。在已有高产、优质品种中导入抗逆性的关键基因,创制一批拥有自主知识产权的高产多抗广适新品种。
(4)基于林木植物基因功能分析的现有成果, 通过同源基因序列分析, 利用基因克隆与表达技术、基因芯片技术、RNAi/DNAi技术以及蛋白质组学技术, 在基础研究层面深入揭示重要林木种发育与代谢过程的分子机制, 深入解析林木生长发育与环境相互适应的分子机制等。
虽然分子生物学手段相比传统研究方法已显示出更大优势,但是通过基因层面对树种的研究只是一种新的方向,并不能完全取代传统方法,仍然需要将分子生物学技术与常规手段紧密配合,加速林木科研进程, 为湖北林业的发展发挥推动作用。
参考文献
[1]Nystedt B., N.R. Street, A. Wetterbom,等. The Norway spruce genome sequence and conifer genome evolution[J]. Nature, 2013,497(7451): 579584.
[2]李军民, 丁小飞, 陈红林,等.长阳栓皮栎天然群体遗传多样性的等位酶分析[J].湖北林业科技,2012(01):34+12.
[3]丁小飞,陈红林,曹健,等.檫木三个群体的遗传结构初探[J].湖北林业科技,2006(05):12.
[4]丁小飞,曹健,陈红林,等.湖北省马尾松天然群体的遗传变异和遗传多样性研究[J].安徽农业科学,2006(13):30573059.
[5]丁小飞,杨桂芬,董梅,等.白皮松天然群体遗传多样性的等位酶分析[J].广东林业科技,2011(01):812.
[6]丁小飞,曹健,陈红林,等.同功酶技术在鹅掌楸属树种分类中的应用研究[J].湖北林业科技,2009(02):58+14.
[7]张婷,刘双青,梅辉,等.湖北省不同地区油茶遗传多样性的AFLP分析[J].安徽农业科学,2011(23):1407014071+14075.
[8]左雪枝.SRAP标记技术优化与湖北油茶遗传多样性研究[D].湖北:华中农业大学,2012.
[9]张婷,刘双青,董妍玲.湖北省油茶种质资源的遗传基础研究[J].河南农业科学,2011(11):5356.
[10]彭婵,李振芳,陈慧玲,等.湖北油茶种质资源SSR分析[J].湖北林业科技,2013(05):14+32.
[11]李良.湖北省油茶优良单株遗传多样性及品质初步分析[D].湖北:华中农业大学,2010.
[12]郭新安.湖北省三大区域群体银杏古树遗传多样性的ISSR分析[D].湖北:华中农业大学,2006.
[13]赵西梅.湖北省大别山区银杏古树种核变异的研究[D].湖北:华中农业大学,2007.
[14]王婷婷.银杏优良单株的遗传基础分析[D].湖北:华中农业大学,2014.
[15]杨琴军,陈光富,刘秀群,等.湖北星斗山台湾杉居群的遗传多样性研究[J].广西植物,2009(04):450454+567.
[16]杨琴军,袁继林,付强,等.台湾杉ISSR反应体系的建立及检测[J].华中农业大學学报,2011(04):432437.
[17]李江伟,杨琴军,刘秀群,等.台湾杉遗传多样性的ISSR分析[J].林业科学,2014(06):6166.
[18]李江伟.基于NSSR和CPSSR标记的台湾杉遗传多样性研究[D].湖北:华中农业大学,2014.
[19]罗世家.珙桐遗传多样性与保护生物学研究[D].湖北:华中农业大学,2012.
[20]熊丹,陈发菊,李雪萍,等.神农架地区濒危植物香果树的遗传多样性研究[J].西北植物学报,2006(06):12721276.
[21]王艇,苏应娟,欧阳蒲月,等.利用RAPD标记分析濒危植物白豆杉种群的遗传结构[J].生态学报,2006(07):23132321.
[22]阮咏梅,张金菊,姚小洪,等.黄梅秤锤树孤立居群的遗传多样性及其小尺度空间遗传结构[J].生物多样性,2012(04):460469.
[23]邓建云,李建强,黄宏文.一株具有特异AFLP指纹图谱的杜仲古树[J].武汉植物学研究,2006(06):509513.
[24]熊丹,谢伟,陈发菊,等.香果树组织培养过程中遗传变异的RAPD分析[J].植物生理学通讯,2008(01):3741.
[25]姚军,张丽芳,涂炳坤,等.基于ISSR标记的少果毛一球悬铃木遗传稳定性分析[J].中南林业科技大学学报,2014(02):611.
[26]王瑞静.黑杨派4个杨树品种~(60)Coγ辐射效应研究[D].湖北:华中农业大学,2009.
[27]黎曙光.湖北省枫杨种质资源的收集及遗传多样性分析[D].湖北:华中农业大学,2007.
[28]万爱华,徐有明,管兰华,等.马尾松种子园无性系遗传结构的地理变异[J].东北林业大学学报,2006(04):1214+48.
[29]肖黎,李晓玲,王玉兵,等.22种木莲属植物亲缘关系的ISSR分析[J].植物研究,2011(04):489494.
[30]肖黎,马太洋,李晓玲,等.22种木莲属植物亲缘关系的SRAP分析(英文)[J].西北植物学报,2011(11):21782184.
[31]辛娜娜.木荷家系遗传及其育种亲本特性的研究[D].湖北:华中农业大学,2014.
[32]袁珊,孟爱平,李建强,等.神农架山体对濒危植物连香树遗传结构影响的研究[J].植物科学学报,2012(04):358365.
[33]文靓.湖北乡土杨树的核心种质构建研究[D].湖北:华中农业大学,2013.
[34]郑瑜.银杏核心种质构建初探[D].湖北:华中农业大学,2010.
[35]张亚东,胡兴宜,宋丛文.利用新型分子标记ESTSSR鉴定湖北省内的主栽黑杨品种[J].分子植物育种,2009(01):105109.
[36]付秀芹,宋兆建,蔡明锋,等.能源植物乌桕杂种苗期形态与光合特性及分子鉴别[J].湖北林业科技,2011(06):1721.
[37]王丽娜.山毛榉科水青冈属(FagusL.)的分子系统发育关系和生物地理研究[D].湖北:华中师范大学,2012.
[38]陈仁芳.桑属系统学研究[D].湖北:华中农业大学,2010.
[39]王璐,雷耘,张明理.基于序列trnLtrnF和ITS的榉属系统发育与地理分布格局的初步分析[J].植物生态学报,2013(05):407414.
[40]杨星宇,杨路路,余志伟,等.水杉木材DNA提取及条形码分子鉴定[J].湖北大学学报(自然科学版),2011(04):397403.
[41]张新叶,宋丛文,张亚东,等.杨树ESTSSR标记的开发[J].林业科学,2009(09):5359.
[42]张新叶,宋丛文,杨彦伶,等.基于EST序列的杨树候选SNPs标记分析[J].华中农业大学学报,2009(06):741745.
[43]杨彦伶,张亚东,张新叶.杨树SSR标记在柳树中的通用性分析[J].分子植物育种,2008(06):11341138.
[44]张新叶,宋丛文,黄敏仁.杨树抗黑斑病相关基因表达谱分析(英文)[J].林业科学,2011(01):8594.
[45]张新叶,张亚东,彭婵,等.水杉基因组微卫星分析及标记开发[J].林业科学,2013(06):160166.
[46]李金宝,唐寅,许锋,等.银杏GbPAL基因启动子表达载体的构建[J].贵州农业科学,2012(05):1719.
[47]李琳玲,程华,程水源,等.银杏查尔酮合成酶基因启动子(GbCHSP)调控元件及功能分析[J].园艺学报,2010(12):19191928.
[48]許锋,孙楠楠,张威威,等.银杏GbMYBF2启动子克隆及序列分析[J].贵州农业科学,2014(04):1620.
[49]龚付全,熊超,陈柳吉,等.银杏CONSTANS基因的克隆与序列分析[J].湖北农业科学,2010(12):29452948.
[50]张威威,宁迎晶,陈柳吉,等.银杏CONSTANS基因植物表达载体构建[J].生物技术,2010(06):810.
[51]许锋,张威威,唐寅,等.银杏叶绿体petD基因的克隆与表达[J].植物生理学通讯,2010(01):3741.
[52]李琳玲,程华,陈小玲,等.银杏类黄酮3羟化酶基因的克隆与表达分析[J].园艺学报,2015(04):643654.
[53]程华,李琳玲,王燕,等.银杏EPSPS基因克隆及表达分析[J].西北植物学报,2010(12):23652372.
[54]程华,许锋,王燕,等.银杏叶绿体铜锌超氧化物歧化酶基因GbCuZnSOD的克隆与表达[J].林业科学,2010(06):3542.
[55]程华,李琳玲,许锋,等.银杏过氧化氢酶基因CAT1的克隆及表达分析[J].林业科学研究,2010(04):493499.
[56]程华,李琳玲,王燕,等.银杏过氧化物酶基因POD1的克隆及表达分析[J].华北农学报,2010(06):4451.
[57]程华,李琳玲,许锋,等.银杏锰型超氧化物歧化酶GbMnSOD基因的克隆与表达[J].园艺学报,2009(09):12831290.
[58]袁红慧,程华,李琳玲,等.银杏MECPs基因启动子克隆及其植物表达载体的构建[J].贵州农业科学,2013(05):1015.
[59]袁红慧.银杏MECT、MECPs基因启动子克隆及植物表达载体构建[D].湖北:武汉工程大学,2013.
[60]袁红慧,程华,李琳玲,等.银杏MECT基因启动子克隆及序列分析[J].湖北农业科学,2015(07):17461750.
[61]邓慧.紫杉醇生物合成酶DBAT启动子的克隆与植物表达载体的构建[D].湖北:华中科技大学,2007.
[62]燕丽娜,苏应娟,王艇.南方红豆杉紫杉烷7β羟基化酶基因全长序列的克隆和进化分析[J].中山大学学报(自然科学版),2009(05):120124.
[63]张志建,郭佳玉,张鹏,等.紫杉醇合成关键酶BAPT基因的克隆及原核表达[J].现代生物医学进展,2010(16):30113014.
[64]于小青.红豆杉细胞遗传转化体系优化及转基因细胞株ORCA3、MET1RNAi基因表达分析[D].湖北:华中科技大学,2011.
[65]于小青,杨海燕,张蒙,等.ORCA3转录因子对红豆杉细胞中紫杉醇生物合成的影响[J].山地农业生物学报,2012(03):189193.
[66]李書涛.调控紫杉醇合成转录因子TcMYC和TcWRKY1的克隆及功能研究[D].湖北:华中科技大学,2012.
[67]张鹏,李书涛,付春华,等.过表达Dbtnbt基因提高中国红豆杉细胞的紫杉醇含量[J].中国生物化学与分子生物学报,2014(04):377382.
[68]谢莎,许想平,张蒙,等.中国红豆杉TcNPR1基因的克隆与功能研究[J].植物科学学报,2014(04):383393.
[69]孔宇飞.悬铃木开花相关基因遗传转化研究[D].湖北:华中农业大学,2014.
[70]易双双.二球悬铃木花发育相关基因功能的初步验证及遗传转化[D].湖北:华中农业大学,2010.
[71]彭文明.不同基因型悬铃木的离体培养及遗传转化研究[D].湖北:华中农业大学,2009.
[72]黄文俊.农杆菌介导不育基因转化悬铃木及长期继代培养植株的遗传稳定性分析的研究[D].湖北:华中农业大学,2007.
[73]张佳琪.二球悬铃木花发育相关基因的克隆及功能研究[D].湖北:华中农业大学,2010.
[74]李志能.二球悬铃木LFY及MADSbox同源基因克隆、功能验证及其系统进化研究[D].湖北:华中农业大学,2008.
[75]熊瑞婷.杨树MXC3基因RNAi载体构建及遗传转化研究[D].湖北:华中农业大学,2015.
[76]徐金艳.两种杨再生体系建立及转Cry1C+9C基因的研究[D].湖北:华中农业大学,2011.
[77]卢小三.两个杨树无性系再生体系的建立及抗虫基因Cry1C+9C转化的初步研究[D].湖北:华中农业大学,2010.
[78]康薇.中嘉8号杨基因转化受体系统的建立及转BtCry1A基因植株检测[D].湖北:华中师范大学,2007.
[79]郑进.BtCry1A基因转化中嘉8号杨的研究[D].湖北:华中师范大学,2006.
[80]张艳.毛竹生长素相关基因的克隆及辐射对其表达的影响[D].湖北:华中农业大学,2010.
[81]梁晓静.毛竹赤霉素相关基因的克隆及辐射对其表达的影响[D].湖北:华中农业大学,2011.
[82]郭少玲.毛竹脱落酸相关基因的克隆及辐射对其表达的影响[D].湖北:华中农业大学,2013.
[83]施雪萍.樟树体细胞胚再生体系的优化和转化Barnase、PaFT基因的研究[D].湖北:华中农业大学,2009.
[84]陈甘明.樟树遗传转化体系的优化及抗寒基因的转化[D].湖北:华中农业大学,2014.
[85]刘冉冉.枫香遗传转化体系的优化及转化PaFT基因的研究[D].湖北:华中农业大学,2010.
[86]景丹龙,马江,张博,等.红花玉兰MwAG基因在花发育不同时期的表达[J].植物学报,2013(02):145150.
[87]张博.星花木兰AGAMOUS同源基因的可变剪接与功能分化[D].湖北:三峡大学,2015.
[88]李虹侠.油桐种子油体发育规律和FADX、PDAT1基因克隆表达[D].湖北:中国科学院研究生院(武汉植物园),2015.
[89]任景,油桐的再生体系建立及其fad2基因克隆与多态性分析[D].湖北:华中农业大学,2011.
[90]许锋,朱俊,张风霞,等.国槐苯丙氨酸解氨酶基因的克隆、反义表达载体构建及遗传转化[J].林业科学研究,2008(05):611618.
[91]肖硕,刺槐高频再生体系的建立及转BtCry1A基因的初步研究[D].湖北:华中师范大学,2009.
[92]田良涛,乌桕胚乳三倍体植株再生与转基因研究[D].湖北:湖北大学,2011.
(责任编辑:唐 岚)
