1960年以来太湖水生植被演变*
2017-04-12周彦锋蒋兆林张秀山王国祥
赵 凯,周彦锋,蒋兆林,胡 娟,张秀山,周 洁,王国祥
(1:南京师范大学环境学院,南京 210023)(2:安庆师范大学资源环境学院,安庆 246011)(3:中国水产科学研究院淡水渔业研究中心,无锡 214081)(4:江苏省太湖渔业管理委员会办公室,苏州 215104)
1960年以来太湖水生植被演变*
赵 凯1,2,周彦锋3,蒋兆林4,胡 娟2,张秀山2,周 洁2,王国祥1**
(1:南京师范大学环境学院,南京 210023)(2:安庆师范大学资源环境学院,安庆 246011)(3:中国水产科学研究院淡水渔业研究中心,无锡 214081)(4:江苏省太湖渔业管理委员会办公室,苏州 215104)
太湖的富营养化污染日益严重,针对太湖水生植被的研究工作非常重要,然而全面的太湖水生植被调查已经有将近二十年未见报道. 基于2014年夏季全湖水生植被调查结果,结合历史资料,比较分析1960年以来太湖水生植被演变情况. 结果表明,1960年以来,共有23种水生植物从太湖消失,其中1981、1997和2014年分别消失7、4和12种. 从分布区面积来看,1960年以来太湖水生植被总体呈北部湖区水生植被消失,东北部、东部及南部湖区水生植被分布区面积持续扩张的态势,1981年全湖水生植被分布区面积占8%,到2014年已经有33.82%的水面有水生植被分布. 从生物量组成来看,太湖水生植被先升后降,从1960年的10×104t,持续上升到1988年的44.72×104t,1997年下降到36×104t,2014年进一步下降到29.09×104t. 但挺水植被以外的水生植被,尤其是浮叶植被的生物量一直保持上升态势. 总生物量的下降与东太湖挺水植被大面积消失有关,到2014年全湖挺水植被生物量比重仅占5.15%,东太湖沼泽化问题已不复存在. 从群落组成变化情况来看,苦草(Vallisnerianatans)群落分布区面积锐减,马来眼子菜(Potamogetonmalaianus)和荇菜(Nymphoidespeltatum)分布区持续扩张. 目前太湖水生植被管理面临的主要问题是北部湖区水生植被恢复和东部湖区水生植被过量生长.
太湖;水生植被;植被演变;植被管理;水草收割
水生植被作为湖泊生态系统的主要初级生产者,在维持水生态健康过程中扮演着至关重要的角色[1-3]. 自从湖泊二相理论[4-6]被广泛应用到富营养化湖泊治理中以来,越来越多的生态学家相信维持水生植被的生长是健康水体生态修复的关键手段及目标所在[7-8].
太湖作为一个得到广泛关注的富营养化水体,其水生植被的研究工作也得到了广泛的开展. 历史上针对太湖的全面植被调查大体分为三个阶段:第一阶段是建国后为了开发利用太湖水生生物资源进行的综合性调查[9];第二阶段是改革开放以后从渔业生产的合理开发角度进行的水生植物生产力调查*曹萃禾.东太湖水生维管束植物调查报告(铅印本),1981.*许兆明.西太湖水生维管束植物调查报告(铅印本),1981.[10-11];第三阶段是1990s以后随着富营养化污染的日益严重而进行的富营养化机理研究中涉及的植被调查[12]. 此后的太湖植被调查仅限局部湖区[13-17],针对全湖水生植被的全面系统调查已经有近二十年未见报道. 在太湖水污染现状日益严峻的今天,作为太湖水生态过程中的重要环节之一,针对太湖全湖的水生植被现场调查紧迫而且必要.
本文基于2014年夏季太湖全湖水生植被调查结果,结合历史资料,比较分析1960年以来太湖水生植被演变情况,试图全面了解太湖水生植被的分布及演变,为太湖水资源管理提供理论依据.
1 方法
1.1 断面设置
调查时间为2014年8月3-14日. 调查范围仅包括太湖岸线以内常年被水淹没的区域,不包括沿岸湖滨带的芦苇群落. 由于太湖面积太大,为便于对植被分布状况进行描述,将全湖划分成9个调查区域分别进行调查,每个调查区内根据预调查结果设置断面,要求断面尽量均匀分布,且能穿过典型水生植被分布区(图1). 调查区内,每条直线视为一个断面,共设置91条断面,总长度899.63 km(表1). 此外,考虑到太湖中大型岛屿周边可能存在水生植物分布,在西洞庭山、乌龟山、三山、漫山等岛屿周边另设样线绕岛航行.
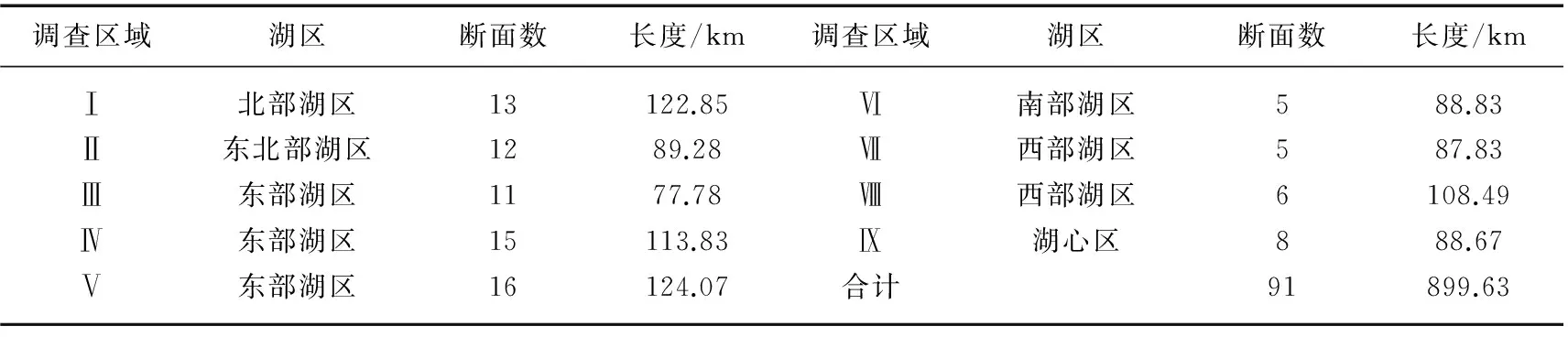
表1 各调查区域断面数及长度
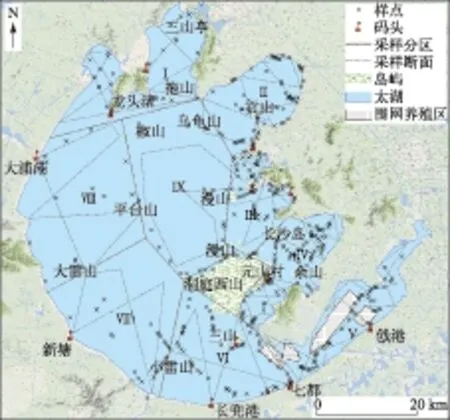
图1 太湖水生植被调查分区及断面设置
1.2 调查方法
调查范围为湖区所有水生植物物种及群落组成,另有少量湿生植物也在湖区内有分布,一并纳入调查范围. 水生植物与湿生植物区别在于:水生植物是指能在淹水环境中完成整个生活史过程的植物;湿生植物是指能在过湿环境中完成整个生活史过程的植物,部分物种因其较强的耐涝性可在淹水环境正常生长,但不能完成整个生活史过程. 根据水生植物各层片的优势种组成对群落进行命名,同一层片的优势种之间用“+”相连,不同层片之间的优势种用“—”相连.
利用船载法对水生植被进行调查,尽量沿断面行船,根据水深及风浪条件判断可能的水生植物分布区,在目测无水生植物分布的水域,每5 km设1个采样点,测量水深、透明度、浊度等环境因子,然后用自制铁钩拖行100~200 m判断水下有无植被分布. 发现有植物分布的水域直接用GPS打点标记,估算盖度,并在典型水生植物群落分布区通过样方法调查物种及群落组成. 挺水植被样方大小为1 m×1 m,直接用镰刀收割样方内所有物种的地上部分,记录物种组成,估算盖度,称取生物量;挺水植被以外的水生植被通过水草采样夹(0.15 m2)进行群落调查. 每个调查区内每种群落设置的样方数至少为5个.
在ArcGIS软件中完成植被图的绘制并计算各群落分布区面积,具体方法为:将全湖标记的485个样点(图1)导入软件中,根据每个样点的水生植被分布情况进行群落分类,先用直线将相邻断面上相同的植物群落连接,再通过同时期遥感影像进行校正,若遥感影像无法分辨群落差别则通过水深、透明度、风浪等水文条件估测可能的水生植被分布区边界. 生物量集中在水底的沉水植物往往无法通过目测估计其盖度,采取的办法为默认其盖度为100%,根据多个样方生物量平均值乘以总分布区面积得出该群落的总生物量. 生物量集中在水面附近的浮叶植物和沉水植物,其水平结构上往往不是均匀分布的,采取的办法为在生物量最大处设置样方,再根据样方处的盖度和总植物群落盖度换算出单位面积生物量,乘以该群落的分布区面积得出总生物量.
2 调查结果
2.1 物种组成
本次调查共记录到维管植物22科31属39种(表2),包括蕨类植物3科3属3种,双子叶植物11科12属14种,单子叶植物8科17属22种. 其中挺水植物11种,沉水植物13种,浮叶植物5种,漂浮植物7种,湿生植物3种.
2.2 群落组成
太湖水生植被总体呈北部湖区和西部湖区裸露,东北、东部、南部湖区广泛分布的格局,湖心仅西竹岛附近有零散的马来眼子菜群落分布,另北部湖区梅梁湖近湾口偶见单株马来眼子菜. 根据水生植物群落优势种组成,将太湖水生植被划分成10种群落类型(图2). 由西向东以马来眼子菜群落逐步过渡到荇菜-马来眼子菜群落,再过渡到其他群落类型,单位面积生物量逐渐增加,群落组成也逐渐复杂.

表2 2014年太湖水生维管植物名录
以挺水植物为主要优势种的群落类型主要分布在东部湖区的东太湖和胥湖近岸区,其他湖区近岸区也有小面积分布,主要优势种有芦苇、菰和莲.
浮叶植物为主要优势种的群落类型以荇菜-马来眼子菜群落最为多见,以南部湖区分布面积最大,东部湖区和东北部湖区也有大面积的片状分布. 荇菜+菱-聚草-金鱼藻群落分布于东部湖区的东太湖苏州湾以西水域、冲山以东水域及胥湖航道北侧水域,适宜淤泥底质的静水水域.
以沉水植物为主要优势种的群落类型包括马来眼子菜群落、苦草群落、微齿眼子菜群落、马来眼子菜-微齿眼子菜群落、微齿眼子菜+黑藻群落、马来眼子菜+聚草-金鱼藻群落. 其中,马来眼子菜群落分布区面积最大,从东北部湖区一直到南部湖区均有大面积连片分布,但该群落或呈零星分布,或呈斑块状分布,总体盖度一般小于1%. 苦草群落分布区位于东西山之间水域. 微齿眼子菜群落分布区位于贡湖东部的羊湾水厂取水口附近水域. 微齿眼子菜+黑藻群落及马来眼子菜-微齿眼子菜群落分布于胥湖近岸水域,生物量较低. 马来眼子菜+聚草-金鱼藻群落大面积分布于东太湖及东西山之间水域.
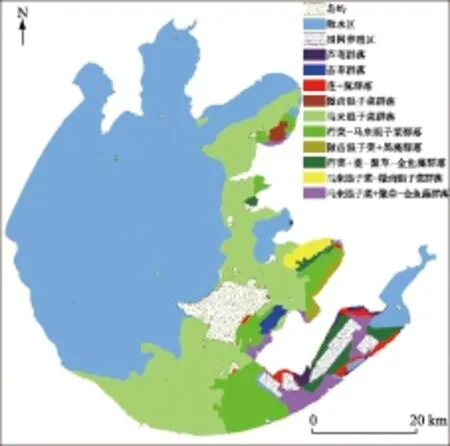
图2 2014年太湖水生植被分布
2.3 生物量分布
2014年夏季太湖湖区水生植被分布区面积共计790.76 km2,占太湖总水域面积的33.82%,夏季总生物量分别为29.09×104t(鲜重)、2.96×104t(干重). 各植物群落分布区面积及生物量分布情况见表3.
从生态型来看,挺水植物群落分布区面积9.87 km2,占太湖水域总面积的0.42%;浮叶植物群落分布区面积168.84 km2,占太湖水域总面积的7.22%;沉水植物群落分布区面积为612.05 km2,占太湖水域总面积的26.18%.
从群落类型分布情况来看,马来眼子菜群落分布区面积最大,占总植物分布区面积的62.64%,但大多分布于与敞水区交界的区域,多成零散分布,总盖度仅为0.3%,因此其生物量总量不大,仅占总生物量的6.21%. 荇菜-马来眼子菜群落是太湖现存植被中生长最为旺盛的群落类型,二者常以面积不等的斑块在水平结构上镶嵌排列,分布区面积居第二位,占总植物分布区面积的18.37%,生物量却达到总生物量的43.84%,居第一位. 马来眼子菜+聚草-金鱼藻群落分布区面积居第三位,占总分布区面积的2.91%,占总生物量的6.48%,也居第三位. 荇菜+菱-聚草-金鱼藻群落见于东部湖区的东太湖、胥湖及冲山附近水域,分布区面积居第四位,但植被生长非常茂盛,占总植被分布区面积虽然只有2.98%,生物量占比却能达到17.97%,居第二位.
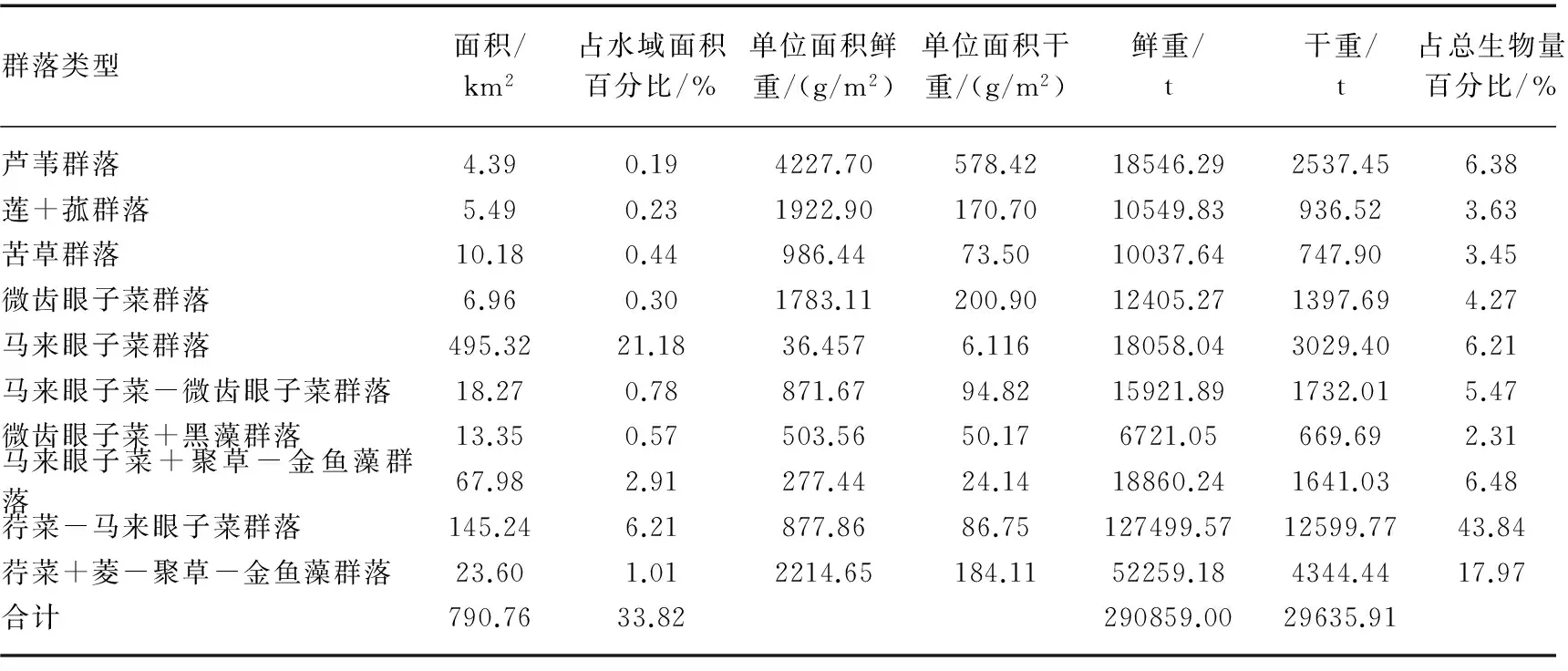
表3 2014年太湖各水生植物群落生物量分布情况
3 水生植被演变
3.1 物种组成变化
1960年的调查共记录66种维管植物[9],1981年为61种,1997年为66种[12],对照物种名录将这些物种进行生态型划分:1960年调查到49种水生植物、17种湿生植物,1981年有45种水生植物、16种湿生植物,1997年水生和湿生植物分别有43和23种. 本次调查(2014年)共记录水生植物36种,湿生植物3种.
对比1960年以来太湖水生植物物种组成变化情况,根据《中国植物志》的分类体系将不同时期调查到的太湖高等植物物种名录进行重新划分,将佛朗眼子菜(Potamogetonfranchetii)并入眼子菜(Potamogetondistinctus)[18],其他种名和学名也进行相应调整. 在此基础上对1960年以来太湖水生植物物种分布情况进行比较分析(表4). 结果表明,共有26种水生植物分布情况发生变化,其中挺水植物10种,浮叶植物4种,漂浮植物2种,沉水植物10种. 这26种植物中,有23种为逐渐消失的物种,其中有7种在1981年调查时消失,4种在1997年调查时消失,12种在2014年调查时消失. 有3种植物为外来迁入物种,其中伊乐藻为1986年人工引入[19],水盾草为外来物种入侵[17],篦齿眼子菜迁入原因不明,鉴于该物种较强的耐污性,可能与水质恶化有关.
3.2 生物量变化
从总生物量变化情况来看,1960年以来,太湖水生植被生物量保持先升后降的态势,从1960年的10×104t上升到1981年的36.82×104t,增加3倍多,到1987年进一步上升到44.46×104t,1988年维持在44.72×104t,1997年下降到36×104t,2014年进一步下降到29.09×104t. 但从生物量组成来看,挺水植被变化趋势与总生物量一样,先升后降,挺水植被以外的沉水植被和浮叶植被则一直保持上升的态势(图3).
1960年的调查中,挺水植被共8×104t,占总水生植被生物量的80%;1981年挺水植被生物量达30.17×104t,占比达81.94%;1987和1988年分别为34.8×104和33.02×104t,分别占78.27%和73.83%;1997年,挺水植被生物量减少到22.5×104t,占比62.5%;到2014年,挺水植被生物量下降到2.91×104t,仅占湖区水生植被生物量的10.00%. 挺水植被以外的水生植被在1960年仅2×104t,占总生物量的20%;到1981年上升至6.65×104t,占比18.06%;1987和1988年达到9.66×104和11.70×104t,分别占总生物量的21.73%和26.16%;1997年进一步上升至13.5×104t,占37.50%;2014年更是达到26.18×104t,占总生物量的90.00%.
从水生植物分布区范围来看,1960年仅在东部湖区的东太湖和东西山之间有大面积水生植被分布[9]. 到1988年,东部湖区的胥湖、光福湾、镇湖湾,东北部湖区的贡湖,北部湖区的梅梁湖、竺山湖等水域均出现不同面积的水生植被,水生植被分布区面积占总水域面积的8%[11]. 1997年北部湖区的水生植被明显退化,仅竺山湖有少量残存,东北部湖区及东部湖区水生植被反而持续扩张,盖度>1%的植被分布区面积占全湖
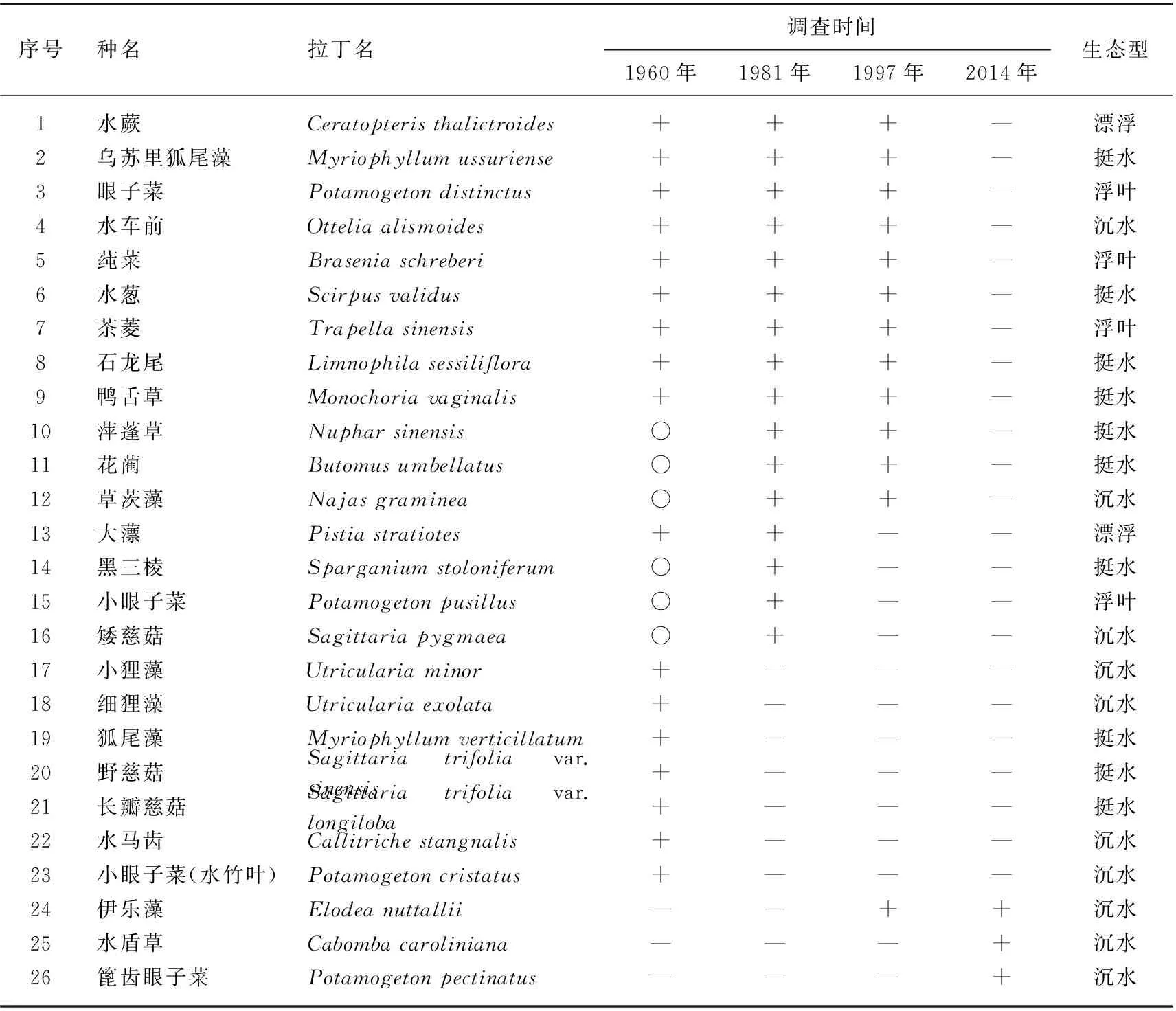
表4 1960年以来太湖水生植物物种变化
*“+”表示有分布;“—”表示无分布;“○”表示推测有分布.
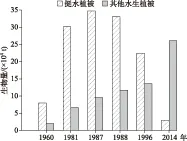
图3 太湖历年夏季水生植被生物量(鲜重)变化[9-12]
水域面积的19%[12]. 2014年夏季,北部湖区植被完全消失,东部湖区水生植被则进一步扩张,胥湖和东西山之间几乎全部有水生植被分布,南太湖更是扩张明显,几乎全部水域都有水生植被分布,甚至蔓延到西部湖区的东侧,全湖水生植被分布区面积达到33.82%.
3.3 群落分布变化
结合历史资料绘制1960年以来历年太湖水生植被空间分布图(图4). 从植被分布区来看,沉水植被和浮叶植被的增加趋势并没有生物量这么明显,这是由盖度差异导致的. 例如,1960年东太湖以外的湖区水生植被分布区范围并不小,但几乎全部为单株或零星分布,生物量不足15 g/m2[9].
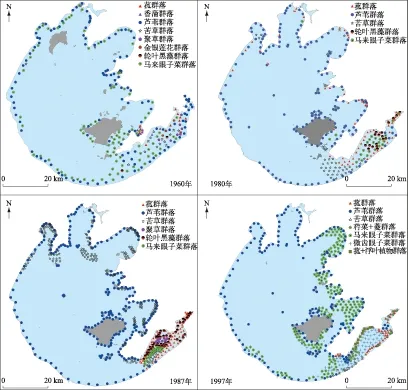
图4 太湖1960[9]、1980[21]、1987[21]和1997年[21]水生植被空间分布
从东太湖水生植被群落组成来看,1960年东太湖优势种为挺水植被菰和芦苇,沉水植被优势种为马来眼子菜和苦草. 1981年挺水植被中菰的优势度上升,芦苇仅占少量,沉水植被中马来眼子菜优势度下降显著,但苦草分布区面积显著上升. 1987年和1988年菰分布区面积和生物量均远大于芦苇,沉水植被中苦草和马来眼子菜为共优种. 1997年东太湖芦苇和菰的分布区面积均有所上升,沉水植被优势种变为微齿眼子菜,浮叶植被荇菜和菱也开始成为建群种,但分布区范围非常有限. 从2002年开始,东太湖菰和芦苇分布区面积锐减,浮叶植被和沉水植被分布区面积及生物量显著上升,荇菜、伊乐藻、马来眼子菜等成为优势种[20]. 2014年东太湖挺水植被分布区面积进一步下降,仅沿岸有少量芦苇群落分布,菰和莲仅在围网外围有人工栽培,其他区域以马来眼子菜和荇菜分布最为广泛.
东太湖以外水域在1960年仅在近岸区的芦苇群落外围有少量马来眼子菜和苦草分布. 1981年,北部湖区的竺山湖和东部湖区的杨湾附近水域均出现大量苦草群落,马来眼子菜群落分布区面积则锐减,生物量仅为1960年的11%,同时,在围网外围开始出现少量荇菜群落. 1988年苦草、聚草和马来眼子菜成为优势种,其中苦草优势度最大,集中分布在北部湖区的竺山湖和东部湖区的杨湾附近水域,聚草则分布于东部湖区的胥湖,马来眼子菜在各湖区零散分布. 1997年北部湖区水生植被基本消失,东北部湖区、东部湖区及南部湖区以马来眼子菜和微齿眼子菜为主要优势种. 2014年北部湖区无水生植被分布,在东部、东北部及南部湖区马来眼子菜群落呈现出由东向西扩张的态势,成为分布区最大的水生植物,而荇菜更是一跃成为分布区第二广泛的水生植物,南部湖区、东部湖区、东北部湖区,均可见一望无垠的荇菜群落,场面蔚为壮观. 微齿眼子菜仍然是主要优势种之一,但分布区转移到了东北部湖区的贡湖和东部湖区的胥湖. 此外,菱、聚草、金鱼藻和黑藻均在东部湖区有成片分布.
4 讨论
自1960年以来,一共有23种水生植物从太湖消失. 从消失的物种组成来看,不仅包括了水蕨、莼菜这样的珍稀濒危物种,也包括了矮慈菇、野慈菇、眼子菜、石龙尾、茶菱这样的广布种. 其余物种如萍蓬草、水车前、乌苏里狐尾藻、狐尾藻、水马齿等曾经都是分布非常广泛的物种,如今在整个长江下游地区都已变得非常罕见,由此推断,太湖水生植物物种多样性的丧失具有一定的普遍性. 此外,莼菜和水葱在太湖周边有大量人工栽培,茶菱、矮慈菇、野慈菇、鸭舌草在周边的农田水网仍有大量分布. 这说明部分消失的水生植物可以通过人工引种和生境修复恢复生长. 在所有消失的物种中,有7种在1960-1981年间消失,4种在1981-1997年间消失,12种在1997-2014年间消失. 鉴于不同时期太湖环境变迁的主要驱动力不同,1960-1981年间消失的物种可能是大规模围湖造田和建闸后水位变化趋于稳定造成的,而1981年以后消失的16个物种则是水质持续恶化和围网养殖、围堰取土、人工水位调控等人为干扰共同导致的.
水生生态系统包括水质浑浊的藻型和水质清洁的草型两种稳态,当水体营养盐上升到一定的阈值后会发生草型湖泊向藻型湖泊的转换,从而造成水环境恶化、生态功能丧失[5]. 大量文献报导了富营养化导致太湖水生植被消失[14,16,22],而太湖的富营养化污染更是已经持续二十余年并保持加剧态势[23-24]. 本研究结果表明,自1960年以来,伴随着愈演愈烈的水体富营养化过程,太湖的水生植被不仅没有消失,相反其分布区面积和生物量却持续上升. 但是,从植被分布区变化格局来看,水生植被分布区上升仅发生在水质相对较好的东部湖区,北部湖区水生植被在1997年以后已经完全消失. 湖泊二相理论从1993年提出以后受到了多方面的挑战,该理论的提出者Scheffer本人于2007年对其进行了完善,提出生态系统的二相转换是受多方面因素调控的,除了营养盐水平之外,还应包括空间异质性、水位变化、湖泊面积、气候变化等因素[6]. 对于太湖这样的超大型湖泊而言,其复杂的空间异质性是毋庸置疑的. 同时,太湖的开阔湖面非常广阔,风浪也成为影响植被分布的重要影响因子之一[12]. 此外,太湖地处经济活跃的苏南平原,人为活动密集,除营养盐输入之外,增殖放流、渔业生产、蓄水调水、水草收割等都对太湖水生植被分布格局造成影响. 因此,对于太湖这样的超大型湖泊而言,其水生植被的分布格局应该是非常复杂的,准确描述其水生植被演变的机制还有待进一步研究.
在2002年之前的研究工作中,东太湖沼泽化一直是被广泛关注的热点问题[15-16,25]. 历史上东太湖一直是一个受人为干扰较小、保持自然演替格局的荒芜水域,挺水植被芦苇分布广泛. 1960年以后开始的大规模围湖造田使得水域面积下降,芦苇群落大面积消失. 围湖造田造成的水流减缓和随后人工引种栽培的菰群落大量繁衍,加速了东太湖的沼泽化进程,到1980s,东太湖挺水植被生物量已超过20×104t. 2000年以后,随着大规模围网养殖,东太湖水生植被遭到严重破坏,尽管沼泽化趋势仍然持续,但生物量已经显著下降[9]. 2009年以后,规范化水产养殖使得东太湖苏州湾以西水域围网面积显著下降,挺水植被在此区域几乎完全消失,浮叶和沉水植被分布区得到部分恢复. 2012年开始,地方政府陆续在东太湖苏州湾及以东水域挖深取土以供城市建设,使得该水域水深普遍达到4 m以上,局部区域甚至达到9 m. 这一系列人为干扰对东太湖水生植被群落组成造成了显著影响,广袤的挺水植被已不复存在,苏州湾及以东水域几无水生植被分布,以西部分几乎全被浮叶和沉水植被占据. 到2014年夏季,东太湖挺水植被仅9.18 km2,占东太湖总水域面积的7.01%,东太湖沼泽化问题已不复存在.
到调查时为止,太湖已经成为一个典型的“草藻混合型湖泊”,即东部、东北部和南部湖区以草型湖泊为主,有大面积水生植被分布,其他湖区则为藻型湖泊,水草消失,蓝藻水华恶性暴发. 综合分析来看,太湖的水生植被演变的主要驱动力包括围湖造田、渔业生产、人工水草收割、富营养化污染和人工水位调控等. 围湖造田的影响主要是在1960s-1990s期间,以东太湖为例,1960-1997年间,围湖造田导致东太湖面积减少120 km2,导致沿岸芦苇群落大面积衰退,湖区的浮叶植物和沉水植物开始新的演替过程[12]. 渔业生产对水生植被的影响体现在1980s后太湖范围内广泛的围网养殖、草食性鱼类的大量投放及破坏底泥的捕捞作业等. 围网养殖导致湖区风浪减小,有利于水生植物生长的同时也加速了湖泊沼泽化进程[12],竺山湖1995-1998年的草食性鱼类养殖可能是导致该水域水草消失的主要原因[12],本次调查发现的光福湾水生植被消失可能与该水域大量投放中华绒螯蟹(Eriocheirsinensis)有关. 人工水草收割对太湖水生植被的影响主要体现在1960s-1980s,此时周边百姓大量捞取太湖水草用作绿肥,芦苇也被大量收割,这直接导致了太湖水生植被生物量的严重下降,1983年以后太湖严禁水草打捞,水生植被开始有所恢复[10]. 此外,受目前东部湖区水生植被大量生长的影响,水生植被集中腐烂,导致水质快速恶化,影响水资源利用的现象时有发生[17]. 2012年开始,太湖苏州部分重新开启了大规模水草打捞,捞草船和人工打捞同步进行,这对太湖水生植被的影响评估尚未见报道. 水位通过影响水生植被的光合作用对其分布格局存在显著影响[26-27],2014年8月对122个样点进行的水深测定显示,太湖平均水深为243.12 cm,对水生植被分布区内74个样点的水深测定结果显示,水生植被分布区平均水深209.22 cm,这个数值已经逼近水生植被分布范围的临界值[28],这可能会使得太湖的水生植被对水位变化变得非常敏感,针对该假设的研究还有待进一步开展.
最近十几年来,遥感技术取得了长足进步,并被应用到太湖植被调查中[29-30],对于长期监控植被演变具有重要意义. 该技术不需要繁重的野外实地调查,仅需进行简单的现场校正即可,可弥补野外实地调查中断面之间水域植被分布情况完全未知的缺陷. 但该技术受天气、光照、风浪、季节等的影响较大,受光谱分辨率的影响较为明显,对于生物量集中在水底的沉水植被难以成功解译,且无法获得生物量数据. 此外,在太湖分布广泛但盖度较小的稀疏马来眼子菜群落也难以通过该方法区分,这使得该方法可能会低估水生植被分布情况. 而野外实地调查则难免存在主观性,例如沿断面行船时,在一个水生植被不呈均匀分布的水域内,难免会刻意驶入密度较大、植被分布较旺盛的水域,在水生植被分布过于旺盛的区域,由于行船困难又不得不绕行,这些都使得野外实地调查可能会高估水生植被的分布情况. Zhao等通过遥感技术对太湖1981-2010年30年间的水生植被分布情况进行分析[30],其主要结果与本文研究结果基本一致,主要体现在三方面:首先,水生植被1981、1995和2010年的分布区面积变化趋势与本文研究结果一致;其次,2010年遥感解译的水生植被分布区面积为341.3 km2,而本次调查虽然发现水生植被分布区面积高于这一数值,但若将本次调查中盖度小于1%的水生植被分布区剔除,2014年太湖水生植被的分布区面积为295.44 km2,二者相差不大;第三,浮叶植物分布区面积持续上升的结果是一致的,即2010年146.2 km2,2014年168.83 km2. 以上分析结果提示,未来太湖的水生植被研究工作需要将两种调查手段有效结合,彼此扬长避短,方可更加精确地获知太湖水生植被分布状况.
5 结论
1)1960年以来累计有23种水生植物从太湖消失,生境变化和人为干扰是物种多样性丧失的主要原因. 建议在局部湖区引种栽培已经消失的水生植被,划定一定水域恢复原始生境,以维持太湖水生植物物种多样性,为将来的水生植被恢复奠定物种基础.
2)在持续富营养化污染二十余年之后,太湖北部湖区水生植被已经完全消失,东部及东北部湖区挺水植被以外的水生植被保持分布区面积和生物量持续上升的态势,尤其以浮叶植被上升最为显著. 太湖是一个人为干扰强烈、影响因素复杂的大型浅水湖泊,其水生植被演变机制受多方面因素调控,针对太湖水生植被的研究工作还需要进一步开展.
3)经过半个世纪的人为活动之后,东太湖的沼泽化问题已经不复存在. 如今的太湖水生植被管理面临的主要问题是北部湖区水生植被消失和东部湖区水生植被过量生长,下一步工作重点应该围绕北部湖区水生植被恢复和东部湖区水草控制展开.
4)未来太湖水生植被分布的研究工作应从以下方面展开:第一,围绕水位变化对太湖水生植被的影响展开全面评估;第二,准确评估2012年开始的大规模水草收割,科学指导太湖的水生植被管理;第三,结合遥感技术和野外实地调查,对太湖水生植被分布格局展开更加精准的研究.
致谢:本次野外调查持续12 d,水上航程近千公里,没有太湖渔政各个中队的支持,不可能顺利完成,在此表示深深的谢意. 长时间的野外调查对样品的及时处理是一个考验,感谢中国水产科学研究院淡水渔业研究中心提供实验室供我们及时分析样品.
[1] Kolada A. The use of aquatic vegetation in lake assessment: Testing the sensitivity of macrophyte metrics to anthropogenic pressures and water quality.Hydrobiologia, 2010, 656(1): 133-147. DOI: 10.1007/s10750-010-0428-z.
[2] Kreiling RM, Richardson WB, Cavanaugh JCetal. Summer nitrate uptake and denitrification in an upper Mississippi River backwater lake: The role of rooted aquatic vegetation.Biogeochemistry, 2011, 104(1-3): 309-324. DOI: 10.1007/s10533-010-9503-9.
[3] Søndergaard M, Johansson LS, Lauridsen TLetal. Submerged macrophytes as indicators of the ecological quality of lakes.FreshwaterBiology, 2010, 55(4): 893-908. DOI: 10.1038/35098000.
[4] Scheffer M, Carpenter S, Foley JAetal. Catastrophic shifts in ecosystems.Nature, 2001, 413(6856): 591-596. DOI: 10.1038/35098000.
[5] Scheffer M, Hosper S, Meijer Metal. Alternative equilibria in shallow lakes.TrendsinEcology&Evolution, 1993, 8(8): 275-279. DOI: 10.1016/0169-5347(93)90254-M.
[6] Scheffer M, van Nes EH. Shallow lakes theory revisited: Various alternative regimes driven by climate, nutrients, depth and lake size.Hydrobiologia, 2007, 584(1): 455-466. DOI: 10.1007/s10750-007-0616-7.
[7] Qin BQ, Yang LW, Chen Fetal. Mechanism and control of lake eutrophication.ChineseScienceBulletin, 2006, 51(19): 2401-2412. DOI: 10.1007/s11434-006-2096-y.
[8] Wang GX, Zhang LM, Chua Hetal. A mosaic community of macrophytes for the ecological remediation of eutrophic shallow lakes.EcologicalEngineering, 2009, 35(4): 582-590. DOI: 10.1016/j.ecoleng.2008.06.006.
[9] Nanjing Institute of Geography, Chinese Academy of Sciences ed. A preliminary report on the comprehensive investigation of Lake Taihu. Beijing: Science Press, 1965:51-54. [中国科学院南京地理研究所. 太湖综合调查初步报告. 北京: 科学出版社, 1965: 51-54.]
[10] Cao Zuhe. Changes of aquatic vascular plant resources in Taihu and their effects on fish productivity.FreshwaterFisheries, 1990, 6: 30-32. [曹萃禾. 太湖水生维管束植物资源变动及其对鱼产力的影响. 淡水渔业, 1990, 6: 30-32.]
[11] Bao Jianping, Miao Weimin, Li Jiefuetal. Preliminary study on the aquatic weeds of the Tai Lake and their rational development and utilization.JournalofDalianFisheriesCollege, 1991, 6(1): 13-20. [鲍建平, 缪为民, 李劫夫等. 太湖水生维管束植物及其合理开发利用的调查研究. 大连水产学院学报, 1991, 6(1): 13-20.]
[12] Qin Boqiang, Hu Weipin, Chen Weiminetaleds. Evolution process and mechanism of water environment in Taihu. Beijing: Science Press, 2004. [秦伯强, 胡维平, 陈伟民等. 太湖水环境演化过程与机理. 北京: 科学出版社, 2004.]
[13] Gu Xiaohong, Zhang Shengzhao, Hu Weipingetal. Evolution of community structure of aquatic macrophytes in East Lake Taihu and its wetlands.ActaEcologicaSinica, 2005, 25(7): 1541-1548. [谷孝鸿, 张圣照, 胡维平等. 东太湖水生植物群落结构的演变及其沼泽化. 生态学报, 2005, 25(7): 1541-1548.]
[14] Liu Weilong, Hu Weiping, Chen Yonggenetal. Temporal and spatial variation of aquatic macrophytes in West Lake Taihu.ActaofEcologicaSinica,2013, 27(1): 159-170. [刘伟龙, 胡维平, 陈永根等. 西太湖水生植物时空变化. 生态学报, 2013, 27(1): 159-170.]
[15] Zhang Shengzhao, Wang Guoxiang, Pu Peiminetal. Succession of hydrophytic vegetation and swampy tendency in the East Lake Taihu.JournalofPlantResourcesandEnvironment, 1999, 8(2): 1-6. [张圣照, 王国祥, 濮培民等.东太湖水生植被及其沼泽化趋势. 植物资源与环境, 1999, 8(2): 1-6.]
[16] Wu Qinglong, Hu Yaohui, Li Wenchaoetal. Tendency of swampiness of East Lake Taihu and its causes.ActaScientiaeCircumstantiae, 2000, 20(3): 275-279. [吴庆龙, 胡耀辉, 李文朝等. 东太湖沼泽化发展趋势及驱动因素分析. 环境科学学报, 2000, 20(3): 275-279.]
[17] Zhao Kai, Li Zhenguo, Wei Hongnongetal. The distribution of aquatic vegetation in Gonghu Bay, Lake Taihu, 2012.JLakeSci, 2015, 27(3): 421-428. DOI:10.18307/2015.0308.[赵凯, 李振国, 魏宏农等. 太湖贡湖湾水生植被分布现状(2012年). 湖泊科学, 2015, 27(3): 421-428.]
[18] Sun Xiangzhong ed. Flora of China (8 volumes). Beijing: Science Press, 1992: 68. [孙祥钟. 中国植物志(第八卷). 北京: 科学出版社, 1992: 68.]
[19] Yang Qingxin, Li Wenchao. Introductions ofElodeanuttalliiin East Tahihu Lake.CollectedPapersofNanjingInstituteofGeographyandLimnology,ChineseAcademyofSciences,1989, 6: 84-92. [杨清心, 李文朝. 伊乐藻在东太湖的引种. 中国科学院南京地理与湖泊研究所集刊, 1989, 6: 84-92.]
[20] Gu Xiaohong, Zhang Shengzhao, Bai Xiulingetal. Evolution of community structure of aquatic macrophytes in East Lake Taihu and its wetlands.ActaEcologicaSinica, 2005, 25(7): 1541-1548. [谷孝鸿, 张圣照, 白秀玲等.东太湖水生植物群落结构的演变及其沼泽化. 生态学报, 2005, 25(7): 1541-1548.]
[21] Taihu Basin Authority, Ministry of Water Resources, Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences eds. Atlas of ecology and environment of the Lake Taihu. Beijing: Science Press, 2000. [水利部太湖流域管理局, 中国科学院南京地理与湖泊研究所. 太湖生态环境地图集. 北京: 科学出版社, 2000.]
[22] Lei Zexiang, Chen Guangrong, Xie Yifaetal. Management of aquatic macrophytes in Lake Taihu.EnvironmentalScienceandTechnology, 2009, 32(6): 189-194. [雷泽湘, 陈光荣, 谢贻发等. 太湖大型水生植物的管理探讨. 环境科学与技术, 2009, 32(6): 189-194.]
[23] Chen YW, Fan CX, Teubner Ketal. Changes of nutrients and phytoplankton chlorophyll-a in a large shallow lake, Taihu, China: An 8-year investigation.Hydrobiologia, 2003, 506(1): 273-279. DOI: 10.1023/B:HYDR.0000008604.09751.01.
[24] Zhu Guangwei. Eutrophic status and causing factors for large, shallow and subtropical Lake Taihu, China.JLakeSci, 2008, 20(1): 21-26. DOI:10.18307/2008.0103. [朱广伟. 太湖富营养化现状及原因分析.湖泊科学, 2008, 20(1): 21-26.]
[25] Li Wenchao. “Yellow water” in East Lake Taihu caused byZizanialatifoliaand its prevention.JLakeSci, 1997, 9(4): 364-368. DOI:10.18307/1997.0412.[李文朝. 东太湖茭黄水发生原因与防治对策探讨. 湖泊科学, 1997, 9(4): 364-368.]
[26] Wantzen KM, Rothhaupt KO, Mörtl Metal. Ecological effects of water-level fluctuations in lakes: An urgent issue. In: Ecological effects of water-level fluctuations in lakes. Netherlands:Springer, 2008: 1-4. DOI: 10.1007/978-1-4020-9192-6_1.
[27] Paillisson JM, Marion L. Water level fluctuations for managing excessive plant biomass in shallow lakes.EcologicalEngineering, 2011, 37(2): 241-247. DOI:10.1016/j.ecoleng.2010.11.017.
[28] Zhang YL, Zhang B, Ma RHetal. Optically active substances and their contributions to the underwater light climate in Lake Taihu, a large shallow lake in China.FundamentalandAppliedLimnology, 2007,170(1): 11-19.
[29] Ma Ronghua ed. Remote sensing of lake environment. Beijing: Science Press, 2010. [马荣华. 湖泊水环境遥感. 北京: 科学出版社, 2010.]
[30] Zhao D, Lü M, Jiang Hetal. Spatio-temporal variability of aquatic vegetation in Lake Taihu over the past 30 years.PLoSOne, 2013, 8(6): e66365. DOI:10.1371/journal.pone.0066365.
Changes of aquatic vegetation in Lake Taihu since 1960s
ZHAO Kai1,2, ZHOU Yanfeng3, JIANG Zhaolin4, HU Juan2, ZHANG Xiushan2, ZHOU Jie2& WANG Guoxiang1**
(1:CollegeofEnvironment,NanjingNormalUniversity,Nanjing210023,P.R.China)(2:CollegeofResourceandEnvironment,AnqingNormalUniversity,Anqing246011,P.R.China)(3:FreshwaterFisheriesResearchCenterofChineseAcademyofFisherySciences,Wuxi214081,P.R.China)(4:FisheriesManagementCommitteeOfficeofLakeTaihu,Suzhou215104,P.R.China)
Comprehensive investigations on aquatic vegetation have been scarcely implemented in Lake Taihu during the past two decades, which is of special importance with the severe eutrophication and pollution. The change in aquatic vegetation in Lake Taihu since 1960 were analyzed based on the exhaustive survey of vegetation in summer, 2014. The results indicated that 23 species disappeared in Lake Taihu since 1960, in which 7 species disappeared in 1981, 4 species disappeared in 1997, and 12 species disappeared in 2014. From the perspective of spatial distribution, aquatic vegetation showed a trend of expansion in the east part and a trend of shrinking in the west of the lake. The coverage rate of aquatic vegetation to the entire lake area was 8% in 1981, whereas, this value increased to 33.82% in 2014. The biomass of aquatic vegetation increased from approximately 1×105t in 1960 to 4.472×105t in 1997, and then decreased from 3.6×105t in 1997 to 2.909×105t in 2014, respectively. However, excluding the emergentPhragmitiesaustraliscommunity, the biomass of other aquatic vegetation, particularly that of the floating-leafed vegetation, continuously increased. A rapid decrease of biomass was closely related to the diminishing of emergent plants in the eastern lake, which accounted for only 5.15% of the total vegetative biomass in Lake Taihu in 2014. It suggests that now the trouble of swampiness does not exist in this area any longer. The change in community composition revealed that the area ofVallisnerianatanssharply decreased, while that ofPotamogetonmalaianusandNymphoidespeltatumprogressively increased. Currently, the main issues of aquatic vegetation management in Lake Taihu are restoring the aquatic vegetation in the northern lake and controlling the overgrowth of aquatic vegetation in the eastern lake.
Lake Taihu; aquatic vegetation; vegetation changes; vegetation management; waterweed harvest
*中央级公益性科研院所基本科研业务费专项(2015JBFM03)、国家自然科学基金项目(41573061)、江苏省太湖污染防治专项(TH2015202,TH2014402)联合资助.2016-04-09收稿;2016-06-27收修改稿.赵凯(1984~),男,博士研究生,讲师;E-mail:zhaokai1911@126.com.
J.LakeSci.(湖泊科学), 2017, 29(2): 351-362
DOI 10.18307/2017.0211
©2017 byJournalofLakeSciences
**通信作者; E-mail:wangguoxiang@njnu.edu.cn.
