餐厨垃圾厌氧消化起泡现象研究
2017-04-11赵小飞王小铭彭绪亚重庆大学三峡库区生态环境教育部重点实验室重庆400045
何 琴,李 蕾,彭 爽,赵小飞,瞿 莉,王小铭,彭绪亚(重庆大学三峡库区生态环境教育部重点实验室,重庆 400045)
餐厨垃圾厌氧消化起泡现象研究
何 琴,李 蕾,彭 爽,赵小飞,瞿 莉,王小铭,彭绪亚*(重庆大学三峡库区生态环境教育部重点实验室,重庆 400045)
以餐厨垃圾中温厌氧消化反应器为研究对象,考察泡沫事件对反应器性能如比沼气产率(SBP)、比甲烷产率(SMP)和挥发性固体(VS)去除率等的影响;并通过分析泡沫前、后系统稳定性参数如挥发性脂肪酸(VFAs)浓度、VFA/总碱度(TA)值和氨氮(TAN)浓度等的变化情况,以及泡沫前、后(包括泡沫层与液体层)的细菌群落结构变化,解析泡沫事件产生的可能原因.结果表明,稳定期的SBP、SMP和VS去除率分别为(0.950 ± 0.104) m3/kg VS、(0.574 ± 0.072) m3CH4/kg VS和(87.14 ± 2.76)%,而泡沫事件显著影响了反应器效率(t检验,95%的置信区间),SBP、SMP和VS去除率分别降为(0.717 ± 0.100) m3/kg VS、(0.432 ± 0.070) m3CH4/kg VS和(84.24 ± 4.44)%.泡沫发生前,系统内出现了VFAs快速积累现象,且易被产甲烷菌消耗的乙酸比例下降,而对泡沫趋势具有增强作用的丙酸比例上升.并且泡沫出现后丝状菌Longilinea arvoryzae和Levilinea,以及黏细菌Cytophaga fermentans的条带强度明显增大,而丝状菌的丝状结构以及粘细菌产生的粘性物质对起泡有一定程度的贡献. 综上,泡沫的产生可能是由系统内大量VFAs积累以及特定微生物大量繁殖的联合作用引起的.
餐厨垃圾;厌氧消化;起泡现象;微生物
餐厨垃圾具有高有机质含量、高含水率及低热值的特性,相比于常规的固体废物处理方法如焚烧和填埋,常选择厌氧消化技术处理餐厨垃圾并回收能源——沼气[1].然而,泡沫问题是困扰厌氧消化系统稳定运行的一大难题[2-3].过去大量的研究者指出不适当的运行条件如超负荷、搅拌不当等会引起泡沫[2];也有不少研究者表明表面活性物质如蛋白质、脂质、VFAs等的积累,会减小表面张力,从而强化泡沫潜能[4].近年来也有研究者转向分析关键微生物的影响,以期寻找泡沫产生的根本原因.如研究者[5-7]发现污泥厌氧消化反应器起泡后大量的 Gordonia或 Microthrix出现,并指出这两种丝状菌是诱导泡沫形成的主要原因.Kougias等[8]研究了经受泡沫事件的粪便消化反应器中微生物群落结构变化,却发现除了常见的泡沫特征细菌 Nocardia 和Desulfotomaculum以外,可产生生物表面活性剂的Lactobacillus和Bacillus以及可减小溶液表面张力的Micrococcus和Streptococcus等的相对丰度在泡沫后显著升高.然而,现有研究大多都专注于活性污泥系统,或污水污泥、粪便厌氧消化系统的泡沫研究.餐厨垃圾厌氧消化系统也极易出现起泡现象[9].但目前,研究者大多只报导了起泡现象的存在,起泡原因及对反应器性能及微生物群落的影响还鲜有进一步的研究.
本研究针对餐厨垃圾厌氧消化反应器的泡沫事件,根据所监测的理化指标,评估泡沫事件对反应器性能的影响;结合稳定性参数的分析,同时采用PCR-DGGE技术对比分析泡沫前、泡沫期(包括泡沫层与液体层)的细菌群落,以期寻找餐厨垃圾厌氧消化反应器中可能的起泡因子.
1 材料与方法
1.1 消化底物和接种污泥
消化底物为取自重庆大学某学生食堂的餐厨垃圾.在取样前,估算运行30d所需的餐厨垃圾量,按照估算量分3份连续3d同一时间同一地点进行取样,将取得的3份餐厨垃圾在大容器中混合均匀.人工去除骨头、塑料袋和纸巾等杂质,用粉碎机制浆并混合均匀,过 10目筛,保证其粒径≤2mm.采用四分法取样测定理化性质,剩余餐厨垃圾用2L的容器分装并密封,于-18℃条件冷冻保存,使用前1d于4℃解冻.反应器运行期间共取餐厨垃圾4次,每次取回后均测定餐厨垃圾理化性质,4次测定结果相差不大,情况如下:pH值为(6.31 ± 0.21),总固体含量(TS)为(28.20 ± 3.41)%,挥发性固体含量(VS)为(26.61 ± 3.25)%,VS/TS为(93.61 ± 1.54)%,碳氮比(C/N)为(14.73 ± 0.34).
接种污泥取自重庆市白市驿某户农家沼气池的室温消化污泥.该消化污泥在接种前过10目筛,去除其中的大颗粒混杂物;并在(37 ± 1)℃预孵化2周,以去除其中残留的原有机消化底物.接种污泥 pH 值为(7.35 ± 0.13),TS为(8.28 ± 0.63)%,VS为(5.54 ± 0.38)%,VS/TS为(67.05 ± 2.62)%,C/N为(10.06 ± 0.43).
1.2 反应器及运行
试验在全自动的完全搅拌釜式反应器(CSTR)中进行,反应器总容积50L,工作容积30L.搅拌转速 90r/min,采用间歇搅拌方式,搅拌时间3h,间歇时间 3h.通过与循环加热水箱相连的容器夹套维持消化污泥温度为(37 ± 1)℃.
一次性向反应器内投加 30L经预孵化后的接种污泥,采用每天进、出料一次的半连续方式运行.反应器运行阶段划分为:驯化期(1~14d)、稳定期(15~70d)、扰动期(71~86d)、泡沫期(87~129d)和恢复期(130~138d).
驯化期和稳定期的运行有机负荷(OLR)均为3.0kg VS/(m3·d).扰动期内,实验楼分别在反应器运行的第71d、72d和75d发生了事故停电事件,时长分别为7h、10h和10h,造成反应器温度波动(污泥温度从设定的(37±1)℃降至(30±1)℃)和搅拌中断.该事件发生后反应器内液面出现了少量的泡沫.为了观察泡沫对系统性能及稳定性的影响,在第83d将OLR提升为4.0kg VS/(m3·d)以强化起泡现象.运行第87d反应器内出现了密集而稳定的泡沫,并持续了 43d.泡沫期间每日记录反应器内所形成泡沫的体积,然后通过加强搅拌和多次从进料口手动回流消化液消除当天形成的泡沫.泡沫中后期和恢复期为了消泡并恢复系统正常运行,采用了降低进料负荷为 2.0kg VS/m3且进料间隔延长为2d的策略.
1.3 物化参数分析
pH值、产气量和气体成分(CH4和CO2)通过在线监测.在每日进料前取 80mL污泥用于其他物化参数的测定.TS、VS采用烘干法测定;总碱度(TA)和总挥发性脂肪酸(TVFA)浓度采用滴定法测定;总氨氮(TAN)采用标准方法[10]测定;C/N采用元素分析仪测定(Elementar VarioELⅢ元素分析仪,德国).VFAs(如乙酸、丙酸、丁酸、异丁酸、戊酸和异戊酸)采用气相色谱法测定:气相色谱仪(Agilent 7890A GC,Agilent,美国),FID检测器,载气为N2,DB-FFAP毛细管柱(30m×0.25mm I.D.,膜厚 0.5μm),进样器和检测器温度分别为250和275℃,毛细管柱升温程序:80℃维持2min,然后以 10℃/min的速率升温至 200℃,并在200℃维持2min.每个指标测定设3个平行样,结果取平均值.
1.4 DNA提取和PCR-DGGE
为了考察泡沫前后微生物群落变化情况,寻找反应系统中的泡沫特征微生物,分别对稳定期、泡沫初期(上层泡沫和下层污泥)、泡沫中期和末期及恢复期的反应器取样进行微生物分析(对应编号1#~6#),取样后立即于-80℃冻存,直至DNA提取处理.采用试剂盒E.Z.N.A.®Soil DNA Kit(OMEGA,美国)提取总DNA,并根据试剂盒说明书进行操作.
根据文献资料,泡沫特征微生物均为细菌[5-8],故只对细菌进行扩增和序列分析.细菌引物采用341F(5′-CCTACGGGAGGCAGCAG-3′)和907R(5′-CCGTCAATTCCTTTGAGTTT-3′)用于细菌16S rDNA的PCR扩增,其中341F的5’端带有40-bp的GC-clamp[11].反应体系50μL,包含各 1.0μL的上游和下游引物(10μmol/L),0.3μL Takara Ex 8Taq HS(5U/μL;Takara Bio,Shiga,日本), 5.0μL 10× Ex Taq Buffer,4.0μL dNTP 9Mixture (2.5mmol/L),20ng纯化后的DNA模板,最后用无菌超纯水补充体积至50μL.扩增反应在PCR仪(Veriti® 96-WellThermal Cycler,Applied Biosystems® Inc.,美国)中进行:94℃预变性 4min; 94℃变性 1min, 59℃退火 1min,72℃延伸 1.5min,循环 30次;在72℃延伸7min;最后于4℃结束反应.
利用DcodeTM基因突变检测系统(Bio-Rad,美国)对 PCR扩增产物进行电泳分离.以去离子甲酰胺和尿素为变性剂,聚丙烯酰胺凝胶浓度为6%(质量浓度),变性梯度35%~70%.将30μL PCR产物与15μL 5×Loading Buffer混合均匀后加样,电泳条件为60℃,120V预电泳20min,80V电泳14h.使用含有 SYBR Green I 染料的 1×TAE buffer(Molecular Probe,美国)染色 30min,然后在GelDoc凝胶成像系统(Bio-Rad,美国)中进行成像分析.
切取代表性条带和优势条带,捣碎后置于30μL灭菌超纯水中,于 4℃隔夜保存.以溶出的DNA为模板,以不含 GC-clamp的引物 341F/ 907R(细菌)进行第2次PCR扩增,反应体系及条件不变.PCR产物经纯化试剂盒(E.Z.N.A.®Gel Extraction Kit,OMEGA,美国)纯化后,克隆到pUM19-T Vector(Vazyme,美国)中,根据产品说明书进行操作.克隆后的基因片段送到北京鼎国昌盛生物科技有限公司测序.测序结果提交到GenBank,通过 NCBI基因序列数据库(http: //www.ncbi.nlm.nih.gov/BLAST/)进行序列相似度搜索.
1.5 数据处理与分析
沼气产量换算成标准状态(0ºC,101.325kPa)下的气体体积.VS去除率和游离氨(FAN)浓度计算方法参考文献[12].比沼气产率(SBP)表示系统添加单位有机质(以1kg VS计)所产生的沼气量(m3/kg VS);比甲烷产率(SMP)表示系统添加单位有机质(以1kg VS计)所产生的甲烷量(m3CH4/kg VS).使用SPSS软件(version 18.0)分析各个理化指标的均值和标准偏差.
使用 Quantity One软件(version 4.6.2)对DGGE图谱中条带的位置及亮度进行数字化,通过非加权配对算术平均法对图谱进行聚类分析;使用SPSS软件对图谱进行主成分分析(PCA);根据公式计算Shannon-Wiener指数和Simpson指数
[13],用以表征样品中微生物的丰富度和均匀度.
2 结果
2.1 厌氧消化反应器效能
从图1可看出,反应器以3.0kg VS/(m3·d)的OLR启动,经历了前期效能参数波动较大的驯化期(1~14d)后,系统在 3.0kg VS/(m3·d)稳定运行56d(稳定期,15~70d).与驯化期相比,稳定期的SBP和SMP显著提高,而VS去除率则变化不明显(t检验,95%置信区间)(表1).
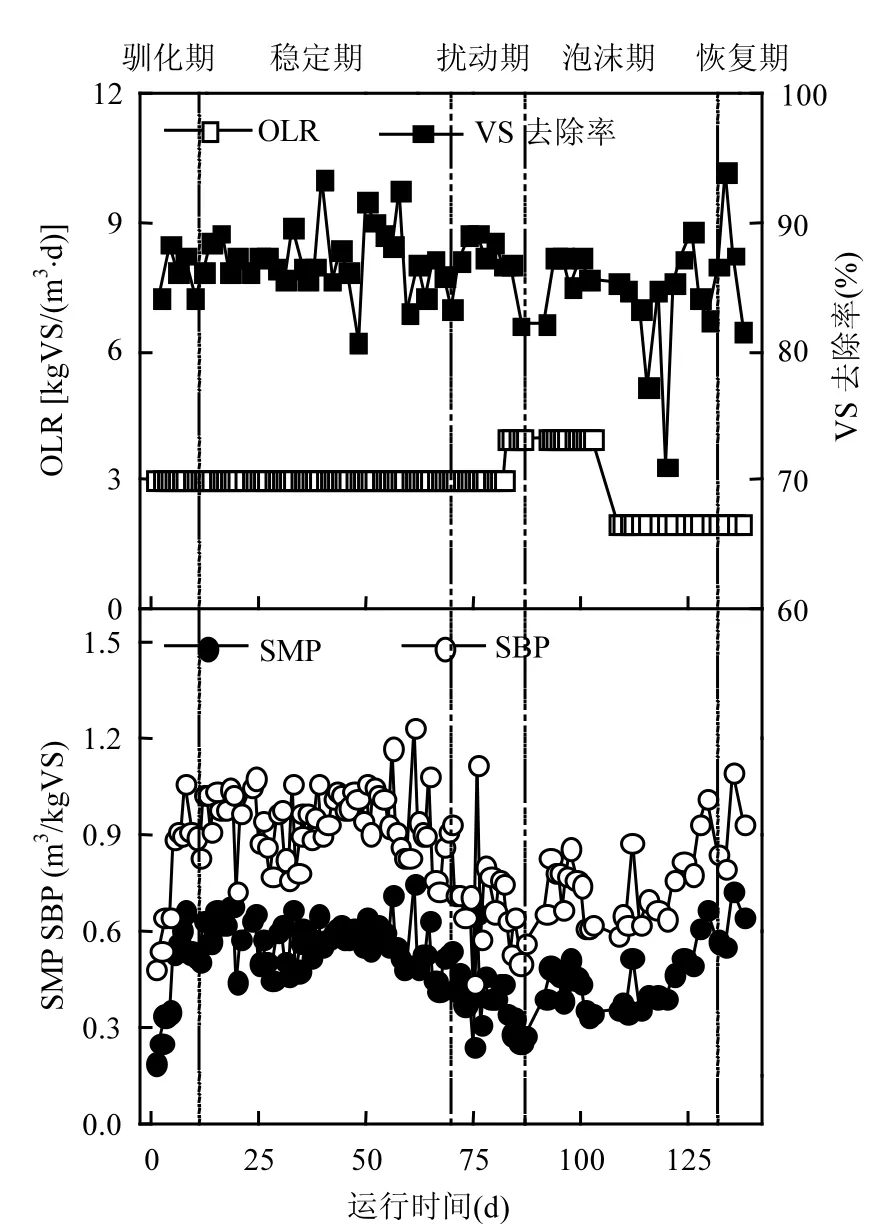
图1 厌氧消化反应器效能参数Fig.1 Performance parameters of the studied anaerobic digester“┆”表示运行阶段的划分界线

表1 各阶段厌氧消化反应器效能参数Table 1 Performance parameters in different stages of the studied anaerobic digester
在经历扰动前期的温度波动和搅拌中断过程中(71~75d),系统SBP和SMP迅速降低,分别从70d的0.923m3/kg VS和0.544m3CH4/kg VS降至0.444m3/kg VS和0.247m3CH4/kg VS(75d),降幅分别达到51.9%和54.6%.而VS去除率仍然没有明显变化.在第76d,SBP和SMP出现了较大反弹,分别增加到了1.122m3/kg VS和0.661m3CH4/kg VS,增幅高达152.7%和167.6%;但在此之后再次下降.分析可能是由于停电期间温度降低短暂影响了微生物酶活性,使得系统进料中的有机物降解不完全,部分残留有机物在温度和搅拌恢复后的系统中被降解,从而使SBP和SMP在短时间内增大;而此后的再次下降,则可能是系统内有机酸积累、pH值降低(图3(a)和(c)),微生物活性受到抑制,系统效能受影响.在第83d将OLR提升为4.0kg VS/(m3·d)后,SBP、SMP和VS去除率持续降低,至第86d,降幅分别为33.1%、42.4%和5.3%.
运行第 87d反应器内出现了大量密集而稳定的泡沫,并持续了 43d (泡沫期,87~129d).泡沫期间,反应器内物质明显分层,上层为棕色泡沫层,下层为黑色污泥层.反应器侧面中上部出料口出料,对上层泡沫进行取样,可观察到上层泡沫实际由大量棕色细小气泡构成,在空气中较稳定、不易破裂;反应器底部出料时可观察到下层污泥中也有大量细小气泡不断向上涌出.记录反应器内泡沫含量(FC)变化,绘制曲线如图 3(d).根据 FC的大小,将泡沫期分为泡沫前期(87~103d,FC>30%)、泡沫中期(104~114d,13%<FC<30%)和泡沫后期(115~129d,10%<FC<13%).泡沫期的SBP、SMP和 VS去除率与稳定期相比,均显著降低(t检验,95%置信区间) (表1).而泡沫后期进料负荷降低,系统效能开始有所恢复,FC也逐渐减小,最后系统进入恢复期(130~138d).与泡沫期相比较,恢复期的 SBP和 SMP均有显著提高(t检验,95%置信区间)(表 1).而恢复期的效能参数与稳定期相比较,没有显著性的差异,证明系统效能恢复.
从图2中可看出,稳定期的累积SBP曲线前段斜率较大,说明进料后微生物能够迅速有效地降解有机物进行产气活动.约 9h后,由于可用的底物开始受限,产气速率逐渐减缓.驯化期的产气曲线与稳定期类似,但前段斜率更小,说明驯化期的微生物产气活性不如稳定期高.扰动期内,停电扰动后产气曲线在进料后出现了约3h的停滞期,可能由于微生物受停电扰动带来的温度波动影响从而活性降低,电力恢复后微生物活性需要一定时间进行修复.负荷提升后,产气曲线斜率较负荷提升前有所降低,可能由于负荷提升导致系统抑制性物质积累,一定程度上抑制了产甲烷微生物的活性.而泡沫期的3条累积SBP曲线的最大斜率呈初期>后期>中期的趋势,说明泡沫初期的产气活性最大,泡沫中期产气活性最小,而泡沫后期通过采用降低负荷等策略,系统产气活性得到了一定程度的恢复.总体来说,产气活性大小呈:稳定期>驯化期>泡沫初期>泡沫后期>停电扰动>泡沫中期>负荷扰动.
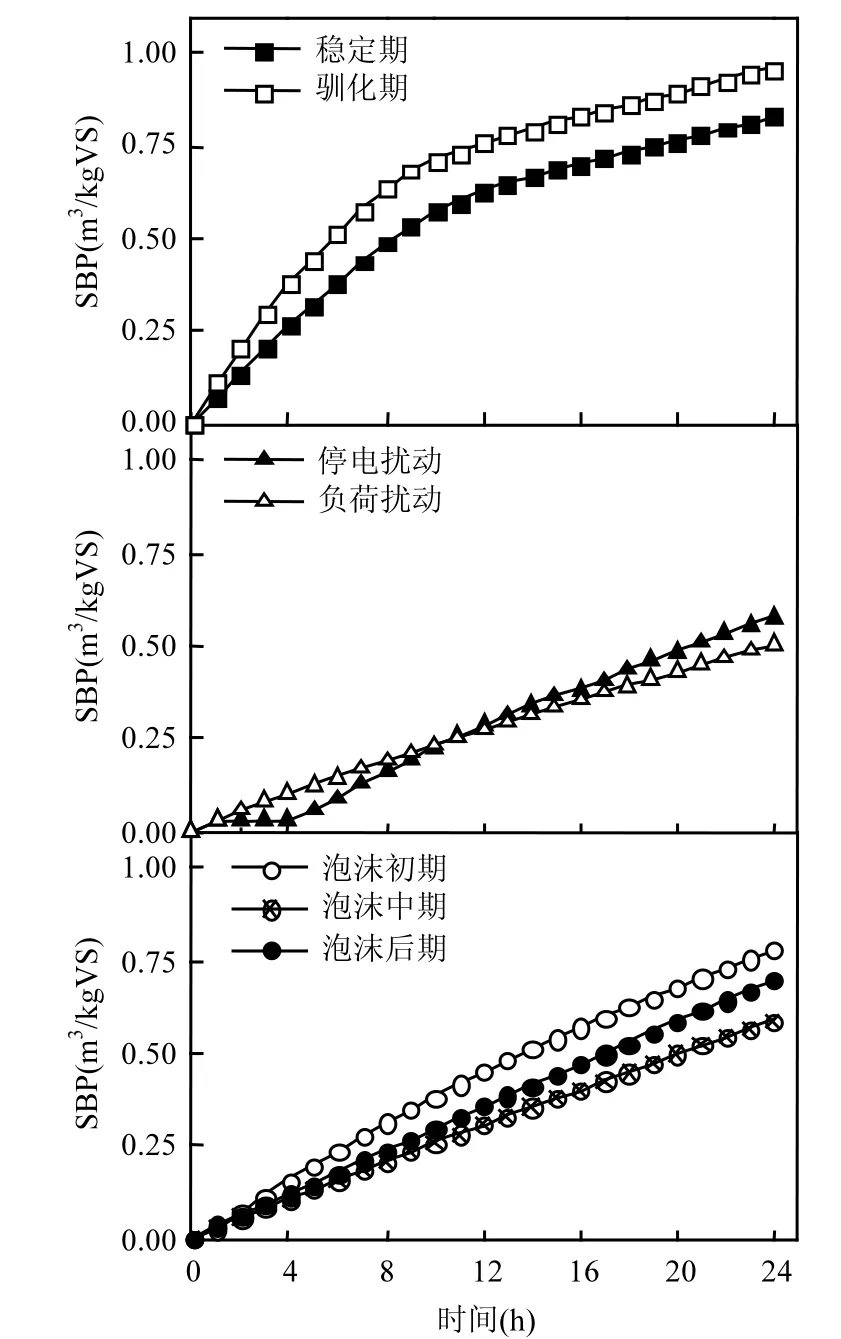
图2 各阶段代表性24h累积SBPFig.2 Typical cumulative specific biogas production for a 24-h period in different stages
2.2 厌氧消化反应器稳定性
从图3(b)可看出,TAN在整个运行期间的总体趋势是逐渐上升的,但在第 35~38d,112~117d, 125~130d出现了3个低峰,每次低峰过后TAN水平均再次升高,运行结束时,TAN达3477mg/L.从图3(a)~(c)可看出,FAN由于受pH值的影响,其变化趋势与pH值的类似,而pH值变化趋势也明显受系统内TVFA浓度的影响,且VFA/TA值的变化趋势也与TVFA类似.
驯化期,TVFA维持在较低水平(1331±279) mg/L,此时,VFA/TA、pH值和FAN分别维持在(0.32±0.06)、(7.32±0.06)和(23±4)mg/L.稳定期,除TAN外的各个稳定性参数均维持比较稳定的状态,TVFA和VFA/TA较驯化期明显降低,分别为(605±392)mg/L和(0.08±0.05),而pH值和FAN均较驯化期有所上升,分别为(7.65±0.08)和(100 ±33)mg/L.扰动期,TVFA的持续上升,导致pH值持续下降,从稳定期末的7.67降到了6.98,均值为7.42,产甲烷菌活性受抑制;VFA/TA在扰动期间迅速增大,停电扰动之后马上超过了稳定性阈值0.350[12],达到 0.434(第 75d),表明系统受抑制;之后该比值继续增大,最高达0.803,均值0.528.泡沫初期,TVFA和VFA/TA值继续迅速增大,即使停止进料(89~92d,图 1(a))也未能使其降低;泡沫中期,第105~109d系统停止进料(图1(a)),TVFA和VFA/TA值有所回落,但第 110d恢复进料 2.0kg VS/(m3·d)后,再次增大;最后将进料间隔延长为2d从而进一步降低日均负荷后(112~129d,图1(a)),TVFA和VFA/TA才逐渐降低,而pH值也逐渐恢复.恢复期时,VFA/TA值降至0.350以下,维持在(0.244±0.054)的水平;此时 TVFA仍在继续降低,pH值继续上升.但pH值的升高,以及TAN的持续升高,一起引起了 FAN的升高,从泡沫期的(83±36)mg/L 上升到恢复期的(323±85)mg/L,最终运行结束时高达 415mg/L,虽然高出了文献中的抑制阈值 200mg/L[14],但从稳定的效能等参数来看,此时的高FAN浓度并没有对系统产生明显抑制.
从图 3(c)和(d)可看出,驯化期,VFAs以乙酸和丙酸为主要成分,分别占 TVFA 的(35.68± 10.21)%和(37.60±4.87)%,其次是正戊酸、异丁酸、异戊酸和正丁酸.而稳定期,乙酸所占比例上升,丙酸相对下降,分别为(54.88±10.68)%和(27.36±9.75)%;正戊酸占(15.70±4.27)%,且正丁酸、异丁酸和异戊酸维持在较低的水平,比例均值均小于 2%.扰动期内,乙酸浓度最初大幅升高,但在达到顶峰后再迅速下降,此时丙酸替代乙酸迅速上升,并使得TVFA继续呈上升趋势,乙酸、丙酸浓度在扰动期分别为(3257±601)和(749±14) mg/L.泡沫期,丙酸浓度迅速升高,远远高于乙酸,成为主导 VFAs((67.21±11.84)%),最高值为13978mg/L,均值9715mg/L.而正戊酸也迅速升高,并在第 94d开始超过乙酸,最高值为 3017mg/L,均值2054mg/L.而其他VFAs也有所升高,但浓度水平仍然不高.泡沫后期,乙酸缓慢回升,其他VFAs尤其是丙酸和正戊酸则迅速降低.直至恢复期,乙酸再次成为主导 VFAs((53.06±26.31)%),丙酸比例(35.53±22.81)%,其他VFAs比例均值均小于5%.
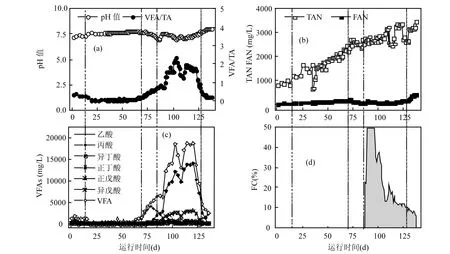
图3 厌氧消化反应器稳定性参数Fig.3 Stability parameters of the studied anaerobic digester“┆”表示运行阶段的划分界线;图3(d)中,第89~96d,实际泡沫界面高出了反应器侧边玻璃视窗顶部,无法准确测量泡沫层实际高度,统一以视窗顶部高度计算泡沫含量,计算值为49.46%,但其实际含量应>49.46%
2.3 PCR—DGGE结果分析
对系统泡沫前后的污泥样品中的细菌进行PCR-DGGE分析,利用Quantity one软件对细菌DGGE图谱进行条带识别,得到泳道/条带识别图,如图4所示.从图4可以看出,不同阶段细菌群落组成基因图谱均表现出了明显的差别.再分别从多样性、聚类和主成分等几方面分析对比泡沫前后系统内细菌群落结构变化,并寻找与泡沫相关的特征细菌.
2.3.1 多样性分析 对所识别的条带图谱进行数字化,计算各阶段样品细菌多样性指数,结果见表2.从表2可以看出,稳定期的细菌Simpson指数值最高,为0.92,表示稳定期的样品中细菌群落较均匀;随着反应器的运行,细菌Simpson指数逐渐降低,尤其是泡沫中后期,下降至 0.82,表明细菌群落均匀度降低;而恢复期又稍微回升. Shannon-Winner指数在泡沫前期的泡沫层中值最大为2.74,但与稳定期的2.62相差不大,随着运行的进行逐渐减小,最后在恢复期有一个回升,说明泡沫层样品中细菌多样性反而最高,泡沫后期最低,而恢复期有所回升.综上,两个指数得出的结果一致,即稳定期的细菌群落多样性最高,系统失稳以及泡沫出现后多样性逐渐降低,但在系统恢复后多样性重新回升.
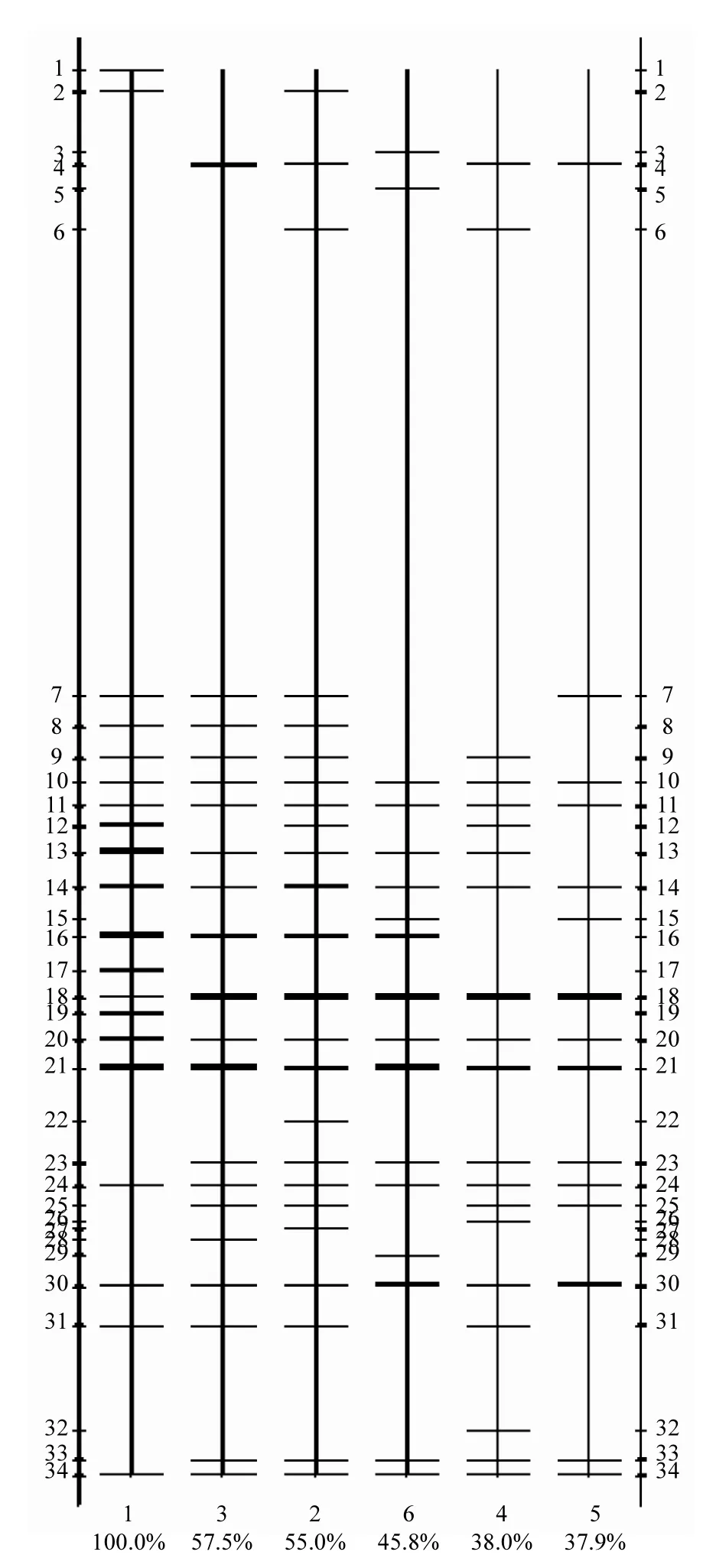
图4 各阶段样品泳道/条带识别Fig.4 Lane images of samples from different stages条带识别均以1#样品为基准,两侧数据表示泳道内条带数,图下方的百分数表示各个样品与1#样品的相似性程度
2.3.2 聚类分析和主成分分析 从图 5(a)可以看出,泡沫初期泡沫层2#样品和污泥层3#样品细菌群落结构相似性最高,聚类指数为0.84,泡沫中期 4#样品与泡沫后期 5#样品相似度也比较高,聚类指数为0.82.来自泡沫期的2#、3#与4#、5#样品的聚类指数也相对较高,为0.71.而恢复期6#样品与泡沫期样品的聚类指数为0.69,稳定期1#样品与其他样品的聚类指数最低,仅为0.47,说明稳定期的细菌群落结构在经历了泡沫抑制后发生了较大的变化.从PCA分析结果图5(b)中也可看出,除2#与3#、4#与5#,以及2#、3#与4#、5#距离较近以外,其他的点与点之间的距离都较大,验证了上述聚类分析的结果,即细菌群落结构经历了泡沫抑制后发生了较大变化.
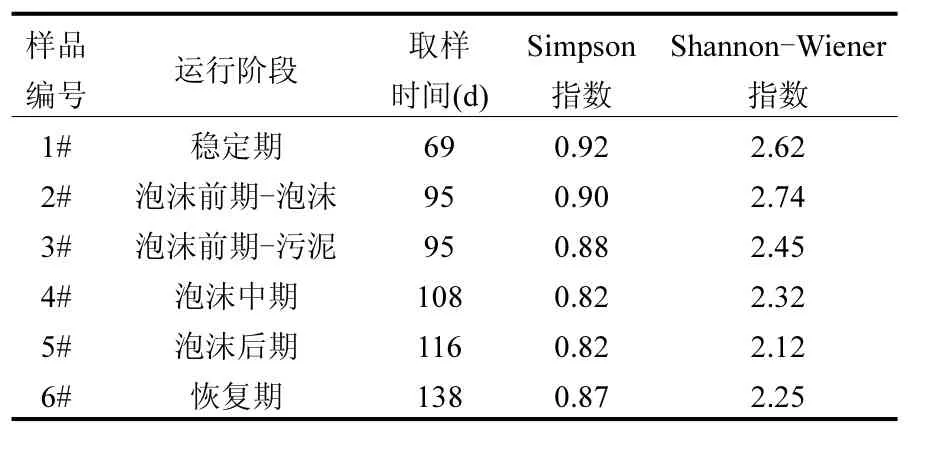
表2 各阶段样品细菌多样性参数Table 2 Bacteria diversity in different stages
2.3.3 测序结果分析 为了寻找泡沫特征细菌,对代表性条带和优势条带进行测序.然后,通过NCBI搜索测序所得16S rRNA基因序列的相似序列,结果见表3.
从表3可看出,所测条带序列大多属于拟杆菌门,拟杆菌门的细菌主要涉及水解和酸化过程,可生成脂肪酶、蛋白酶和纤维素酶以及其他胞外酶[15].例如,检测到的属于 Natronoflexus属的条带 9,可将多种碳水化合物发酵生成乙酸和琥珀酸
[17].条带10和11分别属于Alkalitalea属和Mangroviflexus属,其最终发酵产物主要为丙酸和乙酸[18-19].Petrimonas sulfuriphila(条带 13)在稳定期条带强度最高,可发酵多种单糖和多糖生成乙酸、H2和CO2的中温厌氧发酵细菌[20].条带20所属的Solitalea属是典型的酸化细菌,可发酵葡萄糖、水解明胶等[21].而 Cytophagafermentans (条带 21)是属于拟杆菌门的兼性厌氧黏细菌,可在发酵过程中产生大量粘液[22].另外,也检测到一些厚壁菌门的成员,如条带 2只存在于稳定期和泡沫前期污泥样品中,属于Dethiobacter属,可利用短链脂肪酸和H2作电子供体[16].条带 31的最近亲缘属于厚壁菌门Sporobacter属,是典型的同型产乙酸菌[25].此外,条带24的最近亲缘是Longilinea arvoryzae;条带25的最近亲缘属于Levilinea属,均为绿弯菌门的严格厌氧丝状菌,并具有长度超过 100μm的多细胞丝状结构[23-24].
而由于L. arvoryzae和Levilinea的丝状生长的生理特性,C. fermentans产生大量粘性代谢产物的特性,以及在泡沫期样品尤其是泡沫层样品中的更高条带强度,这些微生物被认为是本试验中的泡沫特征微生物.

图5 细菌聚类分析和PCA图Fig.5 Cluster analysis and PCA of bacteria
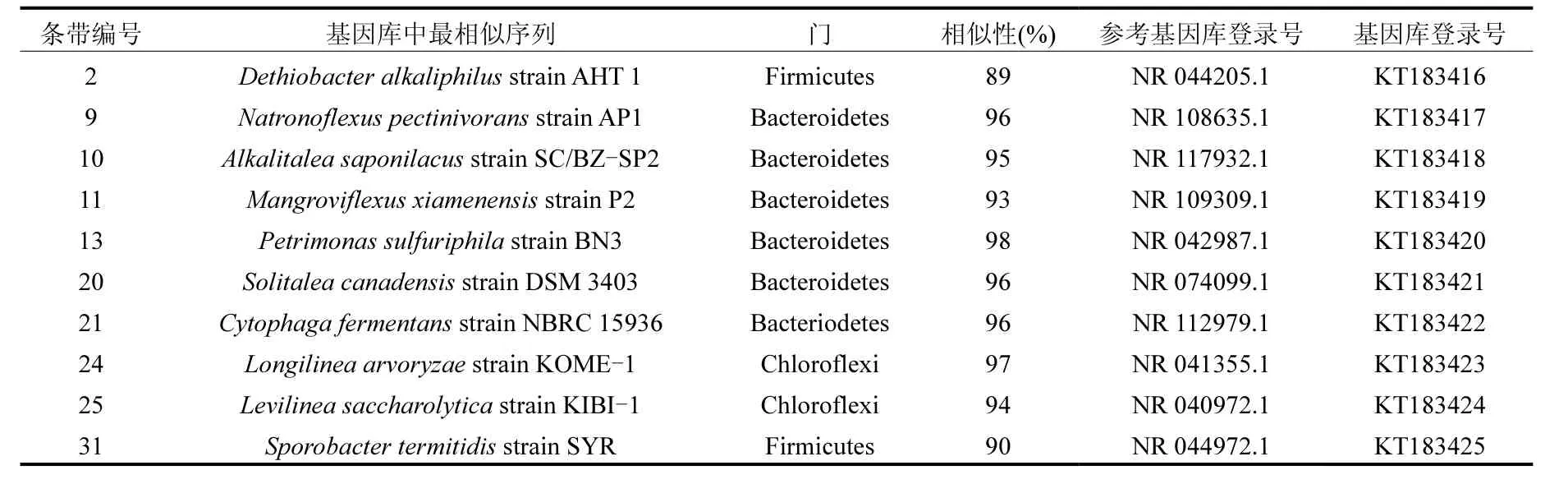
表3 细菌DGGE条带16S rRNA基因序列的亲缘关系比对结果Table 3 Phylogenetic affliation of the 16S rRNA gene sequences from DGGE bands of bacteria.
3 讨论
3.1 泡沫对厌氧消化反应器效能的影响
第15~70d系统在3.0kg VS/(m3·d)负荷下稳定运行,SBP和SMP分别稳定在(0.950 ± 0.104) m3/kg VS和(0.574 ± 0.072) m3CH4/kg VS,与Li等[12]所报道的水平相近.VS去除率稳定在(87.14 ± 2.76)%.反应器分别在71、72和75d经历了连续 3次由实验室事故停电引起的温度波动和搅拌停止,再将OLR从3.0提升到4.0kg VS/(m3·d)后,反应器在第87d形成了大量密集泡沫,甚至在第 89~96d,泡沫层体积超过了污泥层体积的49.46%.Kougias等[26]在对橄榄油厂废水和猪粪联合厌氧消化的研究过程中,在相近的负荷3.6kg VS/(m3·d)下也观察到了泡沫现象.
泡沫期间,由于产甲烷过程的抑制作用,以及大量气泡滞留在污泥内部尤其是上层泡沫中, SBP和 SMP波动较大,分别为(0.717 ± 0.100) m3/kg VS和(0.426 ± 0.075) m3CH4/kg VS,与稳定期相比,均显著降低(t检验,95%置信区间),均值分别下降24.5%和24.8%.相比之下,VS去除率虽然只比稳定阶段低3.3%,但经t检验,变化仍然显著(95%置信区间).Kougias等[26]报道称其反应器在经历泡沫事件后VS去除率降低了11.22%,并将此归因于固体有机颗粒在气液界面积累,造成有机物不能得到有效降解.但本研究中的 VS去除率降低则更可能是由于微生物活性受抑制,从而系统对有机物的降解效率下降.
3.2 泡沫可能的原因简析
3.2.1 VFAs积累 在密集泡沫出现前,反应器内观察到了一定程度的VFAs积累现象,总VFA最高积累到 7372mg/L(86d,其中乙酸主导).随后在泡沫期发生了严重的过酸化现象,总VFA最高达到 18658mg/L (121d).而且在泡沫期,泡沫层中的 VFAs浓度高于液体层(例如,第 95d,泡沫层14284mg/L,液体层12356mg/L);另外丙酸迅速替代乙酸成为VFAs的主要成分(图3(c)).Zhang等[9]的餐厨垃圾厌氧消化反应器在经历了泡沫后,也观察到了同样的 VFAs成分转变.而这种转变可能是由于系统氢积累后,酸化细菌利用高挥发性脂肪酸如丙酸、戊酸来积累电子[27].
猜测 VFAs积累可能对泡沫的引发有贡献,因为VFAs(如丙酸)的羧基端,表现出表面活性剂性质,它们趋于随微小的沼气气泡上升,并在气液界面积累,然后减小消化液的表面张力,增强泡沫趋势[3].而且,Ganidi等[5]也报道称VFA积累是污泥消化反应器中起泡的原因之一.
3.2.2 泡沫特征微生物 对比泡沫期与稳定期的细菌DGGE图谱,发现一些可能与泡沫相关的微生物.L. arvoryzae最初在稳定期的1#样品中被检测到具有较低的条带强度,随后在泡沫初期尤其是泡沫层样品中条带强度显著增大,随后在泡沫中期和后期随起泡强度的减小而逐渐减小.Levilinea最初在泡沫期被检测到,且在泡沫层中条带强度要高于液体层,其条带强度也随泡沫比例的下降而减小.它们都是严格厌氧的丝状菌,具有长度超过100μm的多细胞丝状结构,猜测其丝状结构对泡沫的产生有一定的贡献.因丝状菌特有的丝状结构,使其捕捉到细小的沼气气泡,而其细胞表面的疏水性驱使其随沼气气泡一同上升至气液界面,从而丝状菌在气液界面积累,并同时产生生物表面活性物质,导致消化液表面张力减小,促进泡沫的形成.Subramanian和 Pagilla[28]的试验结果也证明,当剩余活性污泥消化系统中缺少泡沫产生的主要因子丝状菌时,通过提高OLR、降低混合等策略并不能成功诱导反应器产生泡沫.而且众所周知,丝状菌是污水厂污泥厌氧消化反应器中泡沫的主要原因之一[5].例如, Lienen等[7]发现工业规模有机废物厌氧消化反应器中出现过量泡沫时,与对照发酵罐相比,存在高丰度的微丝菌M. parvicella. Ganidi等[6]在污水污泥批次和工业规模厌氧消化反应器中检测到了几株属于Microthrix和Nostocoida属的丝状菌.然而,这些丝状菌是经进料剩余活性污泥或初沉污泥引入消化反应器的,而本研究中并未使用剩余活性污泥而是使用餐厨垃圾作为消化底物,故没有检测到这类丝状菌.而且这类丝状菌虽是好养微生物,但报道称它们也能在缺氧或厌氧条件下生存[5];并由于最佳温度较低而常在污水厂的春季和冬季大量繁殖.故而污泥厌氧反应器因进料污泥中丝状菌的季节性爆发随之产生季节性的泡沫现象[7].而本实验中发现的丝状菌均为严格厌氧菌,它们的大量繁殖可能是系统不稳定导致产酸菌的代谢产物(如VFAs)积累而诱发的.
C. fermentans最早由Bachmann[22]发现,是属于拟杆菌门的兼性厌氧黏细菌,可在发酵过程中产生大量粘液.虽然C. fermentans不具有丝状结构,但其代谢产物——含氮粘性物质对于起泡也有一定的贡献.因粘性物质趋于在沼气气泡周围形成一层液体膜,防止气泡破灭,从而促进泡沫的形成,提高其稳定性.Kougias等[8]在研究经受泡沫事件的粪便消化反应器时观察到的Lactobacillus, Bacillus等,也是通过所产生的具有表面活性的代谢产物(如乳酸或环脂肽表面活性素)对泡沫的形成产生贡献.另外,C. fermentans的条带强度随反应器的运行而逐渐增大,并在泡沫期和恢复期成为优势菌.而C. fermentans产生的大量粘液类似于含氮物质,故需要大量的氮源,而其最佳氮源——铵盐随时间的持续积累(图3(b)),可能是C. fermentans条带强度随时间增大的原因.虽然C. fermentans条带强度逐渐增大,但泡沫比例并未因此而上升,而是随着 VFAs的迅速降低以及丝状菌丰度的减小而减小,故推测C. fermentans的大量繁殖可能对系统最初的泡沫产生有一定的贡献,但在泡沫末期和恢复期,由于缺少了丝状菌和 VFAs积累两个因子,其起泡作用效果就不明显了.另外,由于C. fermentans大量的繁殖,并可发酵葡萄糖产生几乎等摩尔量的乙酸、丙酸和丁二酸,所以它也对失稳阶段的VFAs积累有一定程度贡献.
3.2.3 厌氧消化泡沫 停电扰动所造成的温度降低使得微生物尤其是产甲烷菌活性降低,反应器内进料底物和有机酸积累.恢复中温后,微生物恢复部分活性并高负荷降解系统中积累的有机物.7d后提升的进料负荷再次增大了微生物代谢负荷,由于增殖速率更大,产酸细菌为适应高底物环境而快速繁殖,其产酸量逐渐超过增殖速率较小的产甲烷菌所能消耗的量,故而系统内 VFAs开始积累[29-30].而由于酸积累导致的 pH值下降进一步抑制产甲烷菌的活性,加剧了 VFAs的积累,系统失稳.随后丝状菌 L. arvoryzae和Levilinea大量繁殖.而底物中的含氮物质转化为氨氮也在系统中持续积累,为粘细菌 C. fermentans的大量繁殖提供了充足的氮源.而泡沫的产生则是 VFAs积累和泡沫特征微生物的大量繁殖的结果.丝状菌利用其特有的丝状结构捕捉沼气气泡,而 VFAs和粘细菌产生的粘性物质则附着在气泡周围的液膜中,减小表面张力,增强气泡稳定性.最后气泡上升并在液面积累形成泡沫层.泡沫后期由于采取降低负荷等措施,随着系统中抑制物 VFAs逐渐被消耗,产甲烷菌活性缓慢恢复.而 VFAs浓度的降低以及丝状菌的减少导致沼气气泡不能得到足够的表面活性物质和丝状结构以形成泡沫,虽然粘细菌在充足的氮源下仍继续增长繁殖,但其产生粘性物质的量不足以形成泡沫,故系统中的泡沫量逐渐减少.而随着抑制物被逐渐消耗系统最终恢复稳定.
4 结论
4.1 经历泡沫事件后系统SBP、SMP和VS去除率显著降低,降幅分别为24.5%、24.8%和3.3%.
4.2 泡沫事件发生前,出现了VFAs的快速积累,且对泡沫趋势具有增强作用的丙酸比例上升;泡沫期,VFAs继续大量积累((13449 ± 4079) mg/L),且丙酸((67.21 ± 11.84)%)完全取代乙酸成为VFAs主要成分.
4.3 细菌群落结构经历泡沫后发生了变化,且多样性有所下降;发现泡沫特征微生物丝状菌 L. arvoryzae和Levilinea,黏细菌C. fermentans在泡沫期样品中条带强度明显增大.
4.4 引起泡沫出现的原因是复杂的,本研究中的泡沫可能是由大量 VFAs积累以及泡沫特征微生物大量繁殖的联合作用引起的.
[1] Tanimu M I, Ghazi T I M, Harun M R, et al. Effects of feedstock carbon to nitrogen ratio and organic loading on foaming potential in mesophilic food waste anaerobic digestion [J]. Applied Microbiology and Biotechnology, 2015,99(10):4509-4520.
[2] Kougias P G, Boe K, O-Thong S, et al. Anaerobic digestion foaming in full-scale biogas plants: a survey on causes and solutions [J]. Water Science and Technology, 2014,69:889-895.
[3] Moeller L, Görsch K, Müller R A, et al. Formation and suppression of foam in biogas plants-practical experiences [J]. Agricultural Engineering (Landtechnik), 2012,67(2):110-113.
[4] Boe K, Kougias P G, Pacheco F, et al. Effect of substrates and intermediate compounds on foaming in manure digestion systems [J]. Water Science and Technology, 2012,66(10):2146-2154.
[5] Ganidi N, Tyrrel S, Cartmel E. Anaerobic digestion foaming causes — A review [J]. Bioresource Technology, 2009,100:5546—5554.
[6] Ganidi N, Tyrrel S, Cartmell E. The effect of organic loading rate on foam initiation during mesophilic anaerobic digestion of municipal wastewater sludge [J]. Bioresource Technology, 2011,102(12):6637-6643.
[7] Lienen T, Kleyböcker A, Verstraete W, et al. Foam formation in a downstream digester of a cascade running full-scale biogas plant: Influence of fat, oil and grease addition and abundance of thefilamentous bacterium Microthrix parvicella [J]. Bioresource Technology, 2014,153:1-7.
[8] Kougias P G, Francisci D, Treu L, et al. Microbial analysis in biogas reactors suffering by foaming incidents [J]. Bioresource Technology, 2014,167:24—32.
[9] Zhang Y, Banks C J, Heaven S. Anaerobic digestion of two biodegradable municipal waste streams [J]. Journal of Environmental Management, 2012,104:166-174.
[10] APHA. Standard methods for the examination of water and wastewater [M]. Washington, DC: American Public Health Association, America Water Works Association, Water Environment Federation, 1999.
[11] Casamayor E O, Schäfer H, Bañeras L, et al. Identification of and spatio-temporal differences between microbial assemblages from two neighboring sulfurous lakes: comparison by microscopy and denaturing gradient gel electrophoresis [J]. Applied and Environmental Microbiology, 2000,66(2):499-508.
[12] Li L, He Q, Wei Y, et al. Early warning indicators for monitoring the process failure of anaerobic digestion system of food waste [J]. Bioresource Technology, 2014,171:491-494.
[13] Gupta P, Ahammad S Z, Sreekrishnan T R. Improving the cyanide toxicity tolerance of anaerobic reactor: Microbial interactions and toxin reduction [J]. Journal of Hazardous Materials, 2016,315:52-60.
[14] 唐 波,李 蕾,何 琴,等.总氨氮在餐厨垃圾厌氧消化系统中的积累及其抑制作用 [J]. 环境科学学报, 2016,36(1):210-216.
[15] Supaphol S, Jenkins S N, Intomo P, et al. Microbial community dynamics in mesophilic anaerobic co-digestion of mixed waste [J]. Bioresource Technology, 2011,102(5):4021—4027.
[16] Sorokin Y, Tourova T P, Mußmann M, et al. Dethiobacter alkaliphilus gen. nov. sp. nov., and Desulfurivibrio alkaliphilus gen. nov. sp. nov.: two novel representatives of reductive sulfur cycle from soda lakes [J]. Extremophiles, 2008,12(3):431-439.
[17] Sorokin Y, Panteleeva A N, Tourova T P, et al. Natronoflexus pectinivorans gen. nov. sp. nov., an obligately anaerobic and alkaliphilic fermentative member of Bacteroidetes from soda lakes [J]. Extremophiles, 2011,15(6):691-696.
[18] Zhao C, Gao Z, Qin Q, et al. Mangroviflexus xiamenensis gen. nov., sp. nov., a member of the family Marinilabiliaceae isolated from mangrove sediment [J]. International Journal of Systematic and Evolutionary Microbiology, 2012,62(8):1819-1824.
[19] Zhao B, Chen S. Alkalitalea saponilacus gen. nov., sp. nov., an obligately anaerobic, alkaliphilic, xylanolytic bacterium from a meromictic soda lake [J]. International Journal of Systematic and Evolutionary Microbiology, 2012,62(11):2618-2623.
[20] Grabowski A, Tindall B J, Bardin V, et al. Petrimonas sulfuriphila gen. nov., sp. nov., a mesophilic fermentative bacterium isolated from a biodegraded oil reservoir [J]. International Journal of Systematic and Evolutionary Microbiology, 2005,55(3):1113-1121.
[21] Weon H Y, Kim B Y, Lee C M, et al. Solitalea koreensis gen. nov., sp. nov. and the reclassification of [Flexibacter] canadensis as Solitalea canadensis comb. nov [J]. International Journal of Systematic and Evolutionary Microbiology, 2009,59(8):1969-1975.
[22] Bachmann B J. Studies on Cytophaga fermentans, n. sp., a facultatively anaerobic lower myxobacterium [J]. Microbiology, 1955,13(3):541-551.
[23] Yamada T. Anaerolinea thermolimosa sp. nov., Levilinea saccharolytica gen. nov., sp. nov., and Leptolinea tardivitalis gen. nov., sp. nov., novel filamentous anaerobes, and description of the new classes Anaerolineae classis nov., and Caldilineae classis nov. in the bacterial phylum Chloroflexi [J]. International Journal of Systematic and Evolutionary Microbiology, 2006,56(Part 6):1331-1340.
[24] Yamada T, Imachi H, Ohashi A, et al. Bellilinea caldifistulae gen. nov., sp. nov. and Longilinea arvoryzae gen. nov., sp. nov., strictly anaerobic, filamentous bacteria of the phylum Chloroflexi isolated from methanogenic propionate-degrading consortia [J]. International Journal of Systematic and Evolutionary Microbiology, 2007,57(6):2299-2306.
[25] Liu F H, Wang S B, Zhang J S, et al. The structure of the bacterial and archaeal community in a biogas digester as revealed by denaturing gradient gel electrophoresis and 16S rDNA sequencing analysis [J]. Journal of Applied Microbiology, 2009,106(3): 952-966.
[26] Kougias P G, Kotsopoulos T A, Martzopoulos G G. Effect of feedstock composition and organic loading rate during the mesophilic co-digestion of olive mill wastewater and swine manure [J]. Renewable Energy, 2014,69:202-207.
[27] Ali Shah F, Mahmood Q, Maroof Shah M, et al. Microbial ecology of anaerobic digesters: the key players of anaerobiosis [J]. The Scientific World Journal, 2014.
[28] Subramanian B, Pagilla K R. Anaerobic digester foaming in full-scale cylindrical digesters—Effects of organic loading rate, feed characteristics, and mixing [J]. Bioresource Technology, 2014,159:182-192.
[29] 李 蕾,何 琴,马 垚,等.厌氧消化过程稳定性与微生物群落的相关性 [J]. 中国环境科学, 2016,36(11):3397-3404.
[30] 王祥锟,闵祥发,李建政,等.产氢产乙酸和产甲烷反应对厌氧消化的限速作用 [J]. 中国环境科学, 2016,36(10):2997-3002.
Foaming phenomenon in anaerobic digestion system treating food waste.
HE Qin, LI Lei, PENG Shuang, ZHAO Xiao-fei, QU Li, WANG Xiao-ming, PENG Xu-ya*(Key Laboratory of Three Gorges Reservoir Region’s Eco-Environment, Ministry of Education, Chongqing University, Chongqing 400045, China). China Environmental Science, 2017,37(3):1040~1050
A serious foaming incident occurred in a mesophilic food waste digester. The effects of foaming on reactor efficiency parameters were investigated, including specific biogas production (SBP), specific methane production (SMP) and volatile solids (VS) removal rate. The possible causes of foaming were evaluated according to a series of stability parameters including volatile fatty acids (VFAs), the ratio of VFA to total alkalinity (VFA/TA) combined with the ammonia nitrogen concentration (TAN), as well as the bacterial community structure in pre- and post-foaming system. The SBP, SMP and VS removal rate during the stable stage were (0.950 ± 0.104) m3/kg VS, (0.574 ± 0.072) m3CH4/kg VS and (87.14 ± 2.76)%, respectively. However, those parameters decreased to (0.717 ± 0.100) m3/kg VS, (0.432 ± 0.070) m3CH4/kg VS and (84.24 ± 4.44)% with the appearance of the foaming incident, which indicated that the efficiency of the digester had been significantly influenced by the foaming incident. Prior to the foaming, there appeared to be a rapid accumulation of VFAs along with a reduction in the proportion of acetic acid in VFAs. The propionic acid, which is believed to play a major role in enhancing the foaming tendency, increased in its proportion in VFAs as well. The filamentous bacteria Longilinea arvoryzae, Levilinea and the myxobacterium Cytophaga fermentans had stronger band intensities after foaming. Their filamentous structure or mucilage can be a significant contributor to the initiation of foaming to some extent. In conclusion, foaming may be caused by the combination of VFAs accumulation and the proliferation of specific bacteria.
food waste;anaerobic digestion;foaming phenomenon;microorganism
X705
A
1000-6923(2017)03-1040-11
何 琴(1988-),女,四川遂宁人,重庆大学环境科学与工程专业博士研究生,研究方向为固体废物污染控制与资源化.
2016-08-02
国家“十一五”科技支撑计划资助项目(2010BAC67B01)
* 责任作者, 教授, xypeng33@126.com
