红松存在/不存在数据的多尺度空间分布格局
2017-04-10聂志文布仁仓李冰伦
聂志文, 布仁仓, 李冰伦
1 中国科学院沈阳应用生态研究所, 沈阳 110016 2 中国科学院大学, 北京 100049
红松存在/不存在数据的多尺度空间分布格局
聂志文1,2, 布仁仓1,*, 李冰伦1,2
1 中国科学院沈阳应用生态研究所, 沈阳 110016 2 中国科学院大学, 北京 100049
红松属小兴安岭地区地带性植被优势种,该地区也是其分布的北缘。在景观尺度上开展红松的分布格局研究有利于进一步了解红松分布机理、未来迁移过程等问题,对其经营和保护有重要意义。将景观指数法与点格局分析法结合,设定8个空间尺度,利用红松存在/不存在数据,通过计算各空间尺度上红松聚集程度和景观指数,分析小兴安岭地区红松种群在多尺度上的分布格局。研究结果表明,小尺度上红松聚集分布明显,随机分布区多处于其聚集分布区的边缘,均匀分布区则散布在其聚集分布区内。景观指数研究表明,通过景观指数可判断红松聚集分布格局趋势,而不能判断均匀分布、随机分布格局趋势,因为它们在多尺度下景观指数波动大,不能用景观指数来描述分布格局。研究得出如下结论:1)红松主要分布在其分布区的核心区域内,在分布区边缘和过渡带上呈随机分布,2)存在/不存在数据能够用来分析种群的多尺度空间分布格局,3)空间尺度的变化会引起树种分布格局的变化,随机分布随尺度增加,边缘化程度加强,4)单一尺度上,景观格局指数不能完全描述种群分布格局;而在多尺度上,变化趋势稳定的景观指数表明聚集分布存在,而波动剧烈的景观指数常与随机分布和均匀分布联系在一起,5)地形因子中,红松对坡度和海拔两个因子变化敏感。
红松;空间格局;景观指数;空间尺度;存在/不存在数据
种群空间分布格局描述种群的空间分布特性,是种群长期受温度、降水等环境因子以及种内种间竞争、扩散限制等生态过程影响所表现出的综合特征[1- 3]。开展种群空间格局分析有助于了解种群生物学特性、环境因子相互作用的基本规律以及与空间分布相联系的生态过程,对揭示种群的形成和维持机制有重要意义[4- 6],但采用某个尺度上的数据回答种群分布格局的全部特征非常难。
目前,分析种群分布格局的方法主要有样方法和距离法。传统样方法通过各项聚集强度指数判断种群的分布格局,但这种方法受限于样方大小,只能测定单一尺度的分布类型。改进的Greig-Smith相邻格子法克服了单一取样尺度对计算带来的影响,同时又保留了样方取样的优点,在研究中应用较广[7-8]。距离法中目前使用较多的是点格局方法。该方法通过将种群个体看作是二维空间中的一个点,组成种群点图,将其在空间中位置转化为二维坐标,计算并分析其分布格局[9]。点格局方法因其能够分析各种尺度下的种群格局和种群间空间相关性,克服了一些传统方法只能在单一尺度下进行分析的缺点,在生态学研究中得到广泛应用,在样地尺度上,分析了植物种群分布格局[2,5-6,9],在景观尺度上,开始研究森林景观、林火研究[10-11]。但在实际工作中,在大尺度上,很难获取每个个体的空间位置,无法确定物种分布格局。能否利用栅格化的林相图物种分布数据(即存在/不存在数据)或者遥感数据等确定物种分布格局,值得探讨。
使用景观格局指数描述景观格局及变化,分析景观格局和过程之间关系是景观生态学最常用的一种空间分析手段,在定量反映景观格局的现状及其变化过程中应用广泛[12-15]。单一景观指数法只能反映景观格局变化,并不能揭示景观要素的空间分布特征。简单地从景观格局指数很难判断物种或者景观的空间分布格局,景观格局指数和分布格局之间到底存在什么样的关系,需要进一步的统计分析。
本研究选取小兴安岭地区的优势树种红松(Pinuskoraiensis),分析其分布格局。目前对红松的空间分布研究,多集中在长白山地区[16-17]。小兴安岭地区研究大多在单个林场范围内[11],或者固定样地内[18- 20],分析红松分布核心区内,在小尺度上其分布格局特征。为了确定大尺度上红松分布格局,栅格化林相图为数据源,通过点格局分析(R 3.1.2)和景观格局指数分析(Fragstats 4.2),探讨了1)物种存在/不存在能否在大尺度上回答物种分布格局,2)物种分布格局在核心区和边缘地带有什么不同,3)景观格局指数能否完全说明物种分布格局。
1 材料和方法
1.1 研究区概况
本研究区位于我国黑龙江省境内小兴安岭中部、北部地区(47°03′—49°15′N,127°07′—129°46′E)。行政区域包括沾河林业局、丰林国家级自然保护区、汤旺河林业局、铁力林业局、鹤北林业局等22个林业局,总面积约4.23×106hm2。该区属北温带大陆季风气候区,四季分明,冬季严寒(1月平均气温-25℃)、夏季凉爽(7月平均气温21℃),年均气温-1—1℃,年降水量550—670mm。地带性植被类型为以红松为优势种的针阔混交林,主要的针叶乔木树种有:红松(Pinuskoraiensis)、红皮云杉(Piceakoraiensis)、鱼鳞云杉(Piceajezoensis)、冷杉(Abiesnephrolepis)、落叶松(Larixgmelinii)等,主要的阔叶乔木树种有:白桦(Betulaplatyphylla)、枫桦(Betulacostata)、黑桦(Betuladavurica)、山杨(Populusdavidiana)、紫椴(Tillaamurensis)、春榆(Ulmuspropinqua)、蒙古栎(Quercusmongolica)、黄檗(Phellodendronamurense)、色木槭(Acermono)、水曲柳(Fraxinusmandshurica)等。受气候和植被条件的影响,土壤母质风化作用较迅速,成土过程较强,土层较厚。地带性土壤为暗棕壤,并且以山地暗棕壤为主。山间谷底、山间盆地及河谷阶地等部位分布着面积较广的白浆土、草甸土、沼泽土和泥炭土。
1.2 数据来源
1.2.1 红松分布数据
栅格大小为90m × 90m的林相图(2000年),如果红松分布在栅格内,认为红松存在,不考虑其分布比例、年龄。采用林相图数据主要是因为,从小尺度样地到大尺度区域上,红松分布存在一个尺度外推问题,在小尺度样地内,容易获取红松个体位置信息,但当尺度增大到景观水平,获取个体位置信息就变得困难,所以只考虑在林相图的栅格上有或者没有红松分布,作为判定红松存在的依据。
1.2.2 环境要素数据
从林相图中提取1∶10万的高程,使用ArcGIS平台(Version9.3)生成数字高程模型(DEM),通过数字高程模型提取坡度、坡形、坡向等地形因子信息。并依据地形划分标准[21],对各地形要素进行分级和赋值(表1)。

表1 地形要素分级与赋值
1.3 研究方法
1.3.1 单变量空间点格局
点格局分析法是将每个个体视为二维空间的一个点,以全部个体组成的二维点图为基础进行格局分析[22]。在本研究中,红松存在栅格相当于一个红松的分布点。Ripley的K(t)函数是进行点格局分析的基础函数。其计算公式如下:
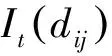
采用完全随机零模型(Complete spatial randomness,CSR),应用Monte-Carlo拟合检验用于计算上下包迹线(envelopes),即置信区间。在99%的置信水平上,拟合次数对为100次[9,23]。如果实际分布的K(t)值在包迹线之内,则表明变量是空间随机分布;在包迹线之上,则表明变量是空间聚集分布;在包迹线之下,表明变量是空间均匀分布。
1.3.2 多尺度分析
对红松分布图,按照50(边长为50个栅格长度的正方形,以下类推),100,150,200,250,300,350,400尺度水平切割,对切割所得的小图计算红松的分布格局。以50尺度为例,设置以50个栅格长度为边长的正方形框,对林相图进行切割,所得的切割数据转换成点形式,进行点格局分析,计算其聚集程度;同时对切割数据计算其景观指数,进行统计分析。将各景观指数均值与多尺度进行拟合,分析二者间相关性。
1.3.3 景观指数
景观指数主要选择聚集度指标及形状指标中所涉及的景观指数,选择时考虑指数的生态学意义,避免重复,并参考以往文献中对于景观指数相关性的研究[14,24],最终选择聚集指数(CONTIG)、分形维数(FRAC)、斑块数量(NP)、斑块密度(PD)以及相似临近百分比(PLADJ)5个指数。
1.3.4 分析工具
使用ArcGIS9.3平台对数据进行处理,调用统计软件R(Version 3.1.2)中spatstat包进行点格局计算, 使用FRAGSTATS(Version 4.2)软件对数据进行景观指数计算。
2 结果与分析
2.1 红松基本分布特征
由图1可以看出,红松在海拔和坡度上分布差别明显。红松主要分布在坡度不超过15°的平坡和缓坡上,只在斜坡、陡坡、急坡上有极少量分布。超过98%的红松分布在了200—800m海拔高度范围内,其中分布在400—600m海拔高度上的红松数量最多,超过了60%。红松在坡向和坡形方面分布差别不明显。其中坡向上,红松只在平地上分布较少,在其他各方位上分布比较均匀;坡形上,研究区内不存在平坡形,红松在凹坡形和凸坡形两个分级上差别不大。
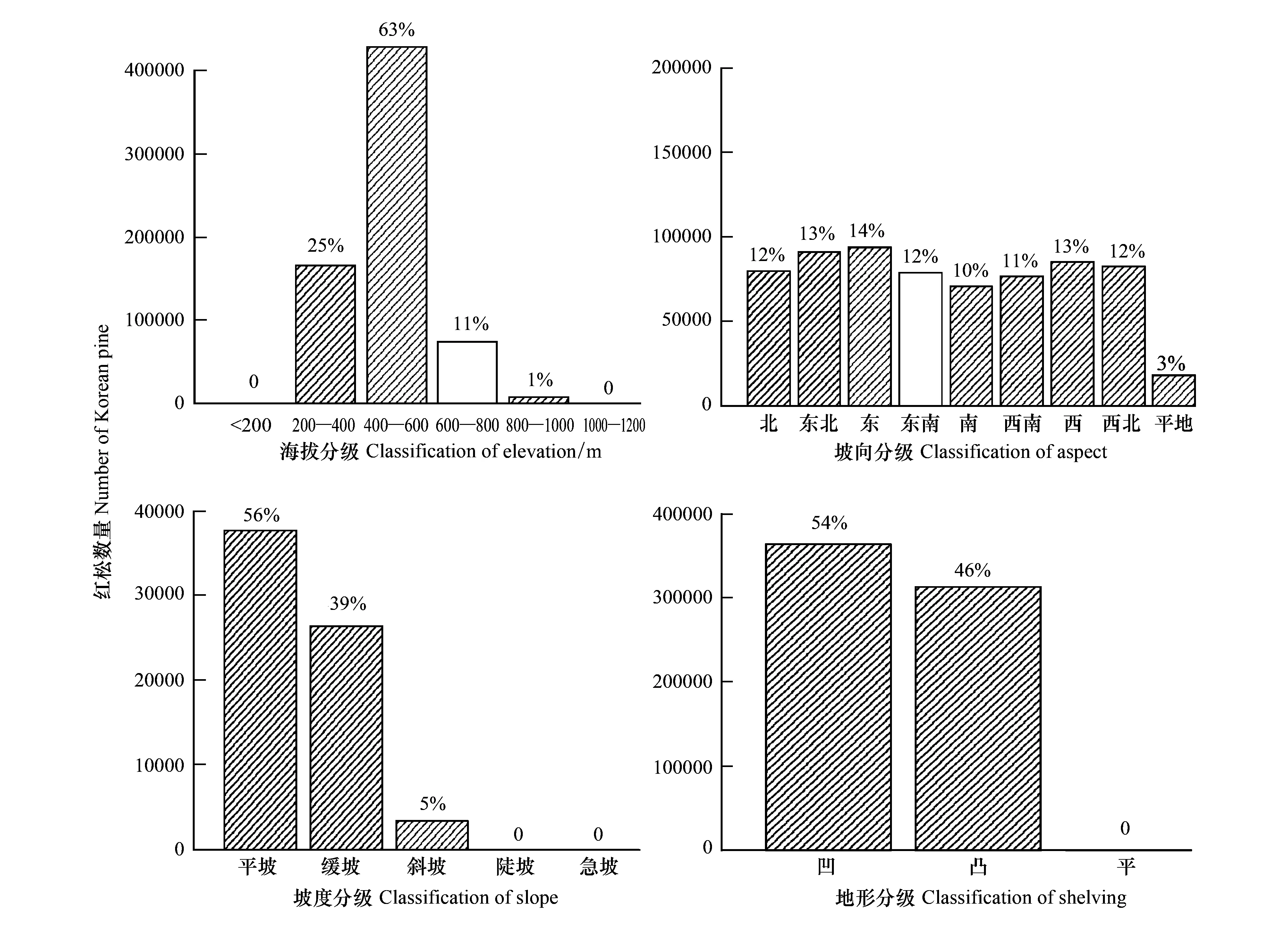
图1 红松地形分布特征Fig.1 The coverage of Korean pine on different geographical factors
2.2 红松多尺度分布特征
随着尺度增加,红松的聚集分布程度表现出增加的趋势,而随机分布和均匀分布斑块数量减少(图2)。在50尺度上,聚集分布是红松主要分布形式,在研究区的中南部地区表现明显;红松的随机分布主要发生在红松聚集分布与无分布区的边缘地带以及其他零星区域;均匀分布则主要发生在红松聚集分布区域内部,且在研究区中部比较明显。在100尺度上,聚集分布仍然是红松主要分布形式,均匀分布程度变化比较明显,出现比率提高,随机分布比率减小,零星分布在红松聚集区域周围。自150尺度至400尺度,红松的均匀分布程度降低,150尺度上存在一定数量分布,400尺度上均匀分布只存在一处;随着尺度变化,聚集分布程度持续增加,400尺度上占据了研究区的大部分区域;随机分布形式随尺度增加,主要出现在研究区的边缘地区,小尺度时在研究区内部零星散布,400尺度上出现在研究区的西北、东南两个区域,表现出随尺度增加,随机分布边缘化特征。
2.3 多尺度景观格局指数
总体上看(图3),聚集分布的景观指数随着尺度上升,变化趋势明显,且波动小,随机分布趋势变化不明显,指数波动较大,表现较杂乱,均匀分布对应的指数变化波动小,趋势较明显,聚集分布下各景观指数与均匀分布下各景观指数呈现相近的分布趋势。在聚集分布模式中,聚集指数(CONTIG)、分形维数(FRAC)、相似临近百分比(PLADJ)变化趋势平稳,斑块数量(NP)指数随尺度增加而增加,斑块密度(PD)随尺度增加逐渐减小。其中聚集指数在各尺度上均值大体上在0.6左右,且与随机分布和均匀分布形式下聚集指数相比较,表现出平稳特征,数值上下波动小。对比3种分布模式,分形维数和相似临近百分比两指数也表现出与聚集指数相同的变化趋势。随机分布形式中,5个指数随尺度增加,表现出杂乱的分布趋势,各指数数值上下波动明显。均匀分布中,随着尺度增加,聚集指数(CONTIG)、分形维数(FRAC)、相似临近百分比(PLADJ)大体表现出围绕某一数值上下波动趋势,斑块数量(NP)呈现先升后降趋势,斑块密度(PD)表现出下降趋势。
采用线性拟合对景观指数和多尺度进行分析(图3),从结果可以看出,景观指数和多尺度间存在着相关关系,且不同指数的相关程度表现不一。斑块密度,斑块数量和相似临近百分比拟合效果较好,其中在聚集分布格局下,斑块数量指数和斑块密度指数与尺度拟合效果好,R2值分别为0.967和0.973,变化趋势明显。相较于在均分分布和随机分布下,分形维数在聚集分布下拟合效果好。聚集指数与多尺度拟合效果一般,聚集指数与多尺度相关性弱。景观指数与分布格局间存在相关关系,聚集分布和随机分布随尺度变化,其聚集指数和分形维数表现出相反的变化趋势。在斑块数量、斑块密度和相似邻近比3个指数上,3种分布格局变化趋势相同,变化强度存在差异。
3 讨论
阔叶红松混交林是小兴安岭地区地带性顶级群落,红松作为其重要组成树种,分布势必受环境因子、人类干扰等因素影响。地形因子是影响植被分布的主要因素,海拔、坡度、坡向等可能会影响到局域生境中光照、水热条件及土壤养分分配,从而对植物种群生长发育及种群结构产生作用,影响其分布特征[2,25]。由结果中可以看出,红松大量生长在平坡、缓坡上,海拔多集中在200—800m范围内,在坡向和坡形各分级上数量差别不明显,这表明红松的分布主要受坡度和海拔的影响,这与Logistic回归模型所得结果一致[26]。坡度小的地区,土壤易于保水,土层较厚,利于植被生长。海拔高度一般会影响立地的温度和光照环境,从而影响树种生长。除地形因子,人类经济活动也会影响红松的分布。红松由于材质好,上世纪末遭到大面积砍伐,砍伐后存在抚育滞后问题,致使红松林斑块面积出现锐减,中幼龄林比重变大[27- 29]。我国自1998年起实施天然林保护工程,小兴安岭林区实施分类经营管理,红松作为保护树种不参与采伐,同时对其他树种的采伐为红松种群提供了生长空间,促进了红松种群的更新和生长[30]。
利用物种分布有/无数据对物种分布进行研究在物种保护和管理上的应用越来越广泛。 基于栅格数据提取的物种分布数据,在预测马尾松潜在分布区变化,探讨欧洲银冷杉的森林经营以及模拟小兴安岭主要阔叶树种的固碳速率研究中是研究主要数据源[31-33]。数据在大尺度上能够反映物种的分布范围,在物种分布模型、景观动态模型模拟中可以满足研究要求。本研究中,基于存在/不存在数据,在大尺度上计算物种的聚集程度,能够反映物种聚集程度变化,具有其合理性。
样地水平上的研究表明,随空间尺度(距离)增加,种群分布格局会发生变化,表现出尺度依赖性[34]。有研究认为,在较小尺度上,种群的空间分布格局受种内竞争、种子扩散方式等影响,在较大尺度上,可能受种群分布的异质性和环境异质性决定[35-36]。种群分布可能同时受一种或多种过程影响,因此随空间尺度变化,表现出不同分布特征。本文结果同样表明,随着空间尺度增加,物种分布格局也在发生变化。小尺度上,红松种
群聚集分布区主要是核心区域,随机分布区散步在核心区域周围,均匀分布区散布在研究区内,当空间尺度增大,聚集分布区占据研究区大部分区域,均匀分布区和随机分布区面积减小,同样表现出尺度依赖性。
自然群落中大部分种群表现为聚集分布格局,这是种群个体在生长和竞争过程中适应环境异质性的结果[19,37]。本研究中在各尺度上红松种群在大部分区域内呈现聚集分布形式,表明聚集分布是小兴安岭地区红松种群的主要分布形式。符合种群分布的一般规律,也与该地区已有的研究结果一致[19]。而在红松聚集分布区向无分布区域过渡带上,红松种群数量会出现减少趋势,所以在这个区域呈现随机分布形式,且多处于聚集分布区域边缘。表明物种在扩散过程中,种群在中心地带聚集,而在边缘地带随机分布。
聚集指数(CONTIG)反映景观中斑块间的邻近程度。随尺度增加,指数数值基本保持不变,表明在景观中同类斑块彼此间距离减小,红松种群聚集程度增加。相似临近百分比(PLADJ)同样描述景观中同类斑块的聚集程度,随着同类斑块的聚集,同类相临栅格的比例会增加,PLADJ指数数值会随之增大。研究中,PLADJ指数保持在80到90之间,反映景观中斑块聚集程度高。分形维数(FRAC)一般用来描述景观形状的复杂程度。斑块数量(NP)统计景观中同类斑块的总数,指数数值大,表明景观中占据面积大,从而决定景观中物种的空间分布特征。斑块密度(PD)主要描述景观内斑块数量和密度特征。在红松聚集分布形式下,其聚集指数和相似临近比指数趋势平稳,说明在整个研究区内,聚集分布是主要的分布形式,斑块数量和斑块密度的变化趋势则显示研究区内,随尺度增加,大量大面积的红松斑块出现,也说明聚集分布将是红松的主要分布形式。这与其他小尺度样地内红松格局研究结果相一致,红松种群在一定尺度上会表现为聚集分布,随尺度变化,聚集程度出现减弱趋势,进而表现为随机分布和均匀分布格局[19]。
“多种景观格局指数联合”、“多尺度景观格局分析”,是景观格局分析的一个发展方向,而结合具体的生态学过程,探讨和建立具有生态学意义的景观格局指数,并用来解释格局、过程间的关系也是亟需解决的问题[38-39]。单一景观指数法只能反映景观格局变化,并不能揭示景观要素的空间分布特征,点格局方法能进行多尺度格局分析,并且直观反映格局特征及变化,将多种景观指数法和点格局方法相结合,互为补充,用来反映景观要素空间分布特征,在研究中有一定的效果,仍有待深入研究。
本研究探索性地将点格局方法和物种存在/不存在数据结合,证明基于栅格数据提取的物种存在数据可以用于物种分布格局研究。但同时需要注意,将物种存在/不存在数据转成点形式的过程中,若只考虑物种是否存在,而未考虑种群其他信息时,所得的分布格局在反映物种分布时,可能会有所欠缺。在日后的研究中,将考虑把种群其他信息引入进来,同时丰富研究方法,从而在分析种群的空间分布特征中取得更好的效果。
4 结论
小兴安岭地区,红松种群在其分布核心区域内主要呈现聚集分布,在聚集区边缘以及过渡带上多为随机分布。地形因子中,坡度、海拔影响种群分布,坡向和坡形对种群分布影响不明显。在多尺度分析中,种群分布格局随着尺度变化,会表现出不同分布特征。在各尺度上,聚集分布形式是红松种群分布的主要形式。此外随尺度增加,随机分布呈边缘化分布趋势。在景观格局指数研究中,在单一尺度上,景观格局指数不能完全描述种群分布格局,而在多尺度上,景观指数变化趋势平稳暗示是聚集分布格局,变化浮动明显的趋势暗示是均匀分布或者随机分布。
[1] 惠刚盈, 李丽, 赵中华, 党普兴. 林木空间分布格局分析方法. 生态学报, 2007, 27(11): 4717- 4728.
[2] Lin Y C, Chang L W, Yang K C, Wang H H, Sun I F. Point patterns of tree distribution determined by habitat heterogeneity and dispersal limitation. Oecologia, 2011, 165(1): 175- 184.
[3] Shen G C, Yu M J, Hu X S, Mi X C, Ren H B, Sun I F, Ma K P. Species-area relationships explained by the joint effects of dispersal limitation and habitat heterogeneity. Ecology, 2009, 90(11): 3033- 3041.
[4] 侯向阳, 韩进轩. 长白山红松林主要树种空间格局的模拟分析. 植物生态学报, 1997, 21(3): 242- 249.
[5] Martínez I, Wiegand T, González-Taboada F, Obeso J R. Spatial associations among tree species in a temperate forest community in North-western Spain. Forest Ecology and Management, 2010, 260(4): 456- 465.
[6] Wiegand T, Gunatilleke S, Gunatilleke N. Species associations in a heterogeneous Sri Lankan dipterocarp forest. The American Naturalist, 2007, 170(4): E77-E95.
[7] 邓莉萍, 白雪娇, 李露露, 牛沙沙, 韩美娜, 秦胜金, 周永斌. 辽东山区次生林优势木本植物种间联结与相关分析. 生态学杂志, 2015, 34(6): 1473- 1479.
[8] 康华靖, 陈子林, 刘鹏, 郝朝运, 韦福民. 大盘山自然保护区香果树种群结构与分布格局. 生态学报, 2007, 27(1): 389- 396.
[9] 张金屯. 植物种群空间分布的点格局分析. 植物生态学报, 1998, 22(4): 344- 349.
[10] 刘志华, 杨健, 贺红士, 常禹. 黑龙江大兴安岭呼中林区火烧点格局分析及影响因素. 生态学报, 2011, 31(6): 1669- 1677.
[11] 王立海, 孟春. 小兴安岭带岭林区红松阔叶林景观多样性与稳定性研究. 应用生态学报, 2005, 16(12): 2267- 2270.
[12] 陈文波, 肖笃宁, 李秀珍. 景观空间分析的特征和主要内容. 生态学报, 2002, 22(7): 1080- 1087.
[13] 刘宇, 吕一河, 傅伯杰. 景观格局-土壤侵蚀研究中景观指数的意义解释及局限性. 生态学报, 2011, 31(1): 267- 275.
[14] 布仁仓, 王宪礼, 肖笃宁. 黄河三角洲景观组分判定与景观破碎化分析. 应用生态学报, 1999, 10(3): 321- 324.
[15] 布仁仓, 胡远满, 常禹, 李秀珍, 贺红士. 景观指数之间的相关分析. 生态学报, 2005, 25(10): 2764- 2775.
[16] 张健, 郝占庆, 宋波, 叶吉, 李步杭, 姚晓琳. 长白山阔叶红松林中红松与紫椴的空间分布格局及其关联性. 应用生态学报, 2007, 18(8): 1681- 1687.
[17] 杨华, 李艳丽, 沈林, 亢新刚. 长白山云冷杉针阔混交林主要树种空间分布及其关联性. 生态学报, 2014, 34(16): 4698- 4706.
[18] 徐丽娜, 金光泽. 小兴安岭凉水典型阔叶红松林动态监测样地: 物种组成与群落结构. 生物多样性, 2012, 20(4): 470- 481.
[19] 侯红亚, 王立海. 小兴安岭阔叶红松林物种组成及主要种群的空间分布格局. 应用生态学报, 2013, 24(11): 3043- 3049.
[20] Liu Y Y, Li F R, Jin G Z. Spatial patterns and associations of four species in an old-growth temperate forest. Journal of Plant Interactions, 2014, 9(1): 745- 753.
[21] 中华人民共和国林业部. 林业专业调查主要技术规定. 北京: 中国林业出版社, 1990.
[22] Ripley B D. Modelling spatial patterns. Journal of the Royal Statistical Society, Series B, 1977, 39(2): 172- 212.
[23] Wiegand T, Moloney K A. Rings, circles, and null-models for point pattern analysis in ecology. Oikos, 2004, 104(2): 209- 229.
[24] 王计平, 杨磊, 卫伟, 陈利顶, 黄志霖. 黄土丘陵区景观格局对水土流失过程的影响——景观水平与多尺度比较. 生态学报, 2011, 31(19): 5531- 5541.
[25] Webb E L, Stanfield B J, Jensen M L. Effects of topography on rainforest tree community structure and diversity in American Samoa, and implications for frugivore and nectarivore populations. Journal of Biogeography, 1999, 26(4): 887- 897.
[26] 布仁仓, 常禹, 胡远满, 李秀珍, 贺红士. 小兴安岭针叶树种在不同尺度上对环境因子的敏感性分析. 植物生态学报, 2008, 32(1): 80- 87.
[27] 王晓春, 孙龙, 周晓峰, 王天明, 国庆喜, 李淑娟. 黑龙江省森林景观的格局变化. 应用与环境生物学报, 2003, 9(2): 111- 116.
[28] 陈雄文. 1896年和1986年黑龙江省几种森林景观的特征变化. 植物学报, 2000, 42(9): 979- 984.
[29] 白卫国, 王祝雄. 论我国东北林区森林可持续经营. 林业资源管理, 2008, (6): 1- 7.
[30] 邓华卫, 布仁仓, 刘晓梅, 贺伟, 胡远满, 黄乃伟. 模拟分类经营对小兴安岭林区森林生物量的影响. 生态学报, 2012, 32(21): 6679- 6687.
[31] 马俊, 布仁仓, 邓华卫, 胡远满, 秦秦, 韩风林. 气候变化对小兴安岭主要阔叶树种地上部分固碳速率影响的模拟. 应用生态学报, 2014, 25(9): 2449- 2459.
[32] 张雷, 刘世荣, 孙鹏森, 王同立. 气候变化对马尾松潜在分布影响预估的多模型比较. 植物生态学报, 2011, 35(11): 1091- 1105.
[33] Falk Wolfgang, Mellert Karl H. Species distribution models as a tool for forest management planning under climate change: risk evaluation ofAbiesalbain Bavaria. Journal of Vegetation Science, 2011, 22(4): 621- 634.
[34] He F L, Duncan R P. Density-dependent effects on tree survival in an old-growth Douglas fir forest. Journal of Ecology, 2000, 88(4): 676- 688.
[35] Harms K E, Condit R, Hubbell S P, Foster R B. Habitat associations of trees and shrubs in a 50-ha neotropical forest plot. Journal of Ecology, 2001, 89(6): 947- 959.
[36] Zhang Z H, Hu G, Zhu J D, Luo D H, Ni J. Spatial patterns and interspecific associations of dominant tree species in two old-growth karst forests, SW China. Ecological Research, 2010, 25(6): 1151- 1160.
[37] 白雪娇, 邓莉萍, 李露露, 牛沙沙, 韩美娜, 秦胜金, 周永斌. 辽东山区次生林木本植物空间分布. 生态学报, 2015, 35(1): 98- 105.
[38] 陈利顶, 刘洋, 吕一河, 冯晓明, 傅伯杰. 景观生态学中的格局分析: 现状、困境与未来. 生态学报, 2008, 28(11): 5521- 5531.
[39] 陈利顶, 李秀珍, 傅伯杰, 肖笃宁, 赵文武. 中国景观生态学发展历程与未来研究重点. 生态学报, 2014, 34(12): 3129- 3141.
Multi-scale spatial pattern analysis of Korean pine based on presence/absence data
NIE Zhiwen1,2, BU Rencang1,*, LI Binglun1,2
1InstituteofAppliedEcology,ChineseAcademyofScience,Shenyang110016,China2UniversityofChineseAcademyofScience,Beijing100049,China
Spatial patterns reflect the distribution characteristics of populations and the adaptions of populations to environmental resources and ecological processes. Dispersal limitation, inter- and intraspecific competition, and habitat heterogeneity all influence species distribution. In addition, species are affected by local temperature, precipitation, and terrain. Therefore, the relationship among species distribution patterns, landscape indices, and environmental factors are important in population studies affected by changing climates. To reveal these relationships, Korean pine, the dominant species in the Small Khingan Mountains, at the northern boundary of its distribution range, was selected to analyze its distribution patterns at multi-spatial scales. The point pattern analysis and landscape pattern metrics were calculated at eight spatial scales (90 m × 90 m resolution) using R package (Spatstat 1.42- 1) and FRAGSTATS (Version 4.2), based on presence/absence data derived from forest inventory maps and attributed data. The results from the point pattern analysis showed that Korean pine was aggregated in small scales, random distribution areas surround an aggregated pattern, and uniform distribution areas were often mosaicked in aggregated pattern areas. However, the distribution pattern of Korean pine was more aggregated with an increasing spatial scale because the aggregated distribution area increased, especially in the core area of this region. Meanwhile, random distribution areas occurred at the edge of aggregated distribution areas, and uniform distribution areas disappeared from the core area. The results from the landscape pattern analysis indicated that landscape pattern indices could be used to describe an aggregated distribution at multi-spatial scales since the landscape metrics were most stable, or changes were logarithmic; whereas landscape pattern indices dramatically fluctuated with changing spatial scales for the others. The analyses also revealed that Korean pine was sensitive to slope and elevation; most individuals were distributed on flat slopes or elevations between 200 and 800 m. Korean pine distribution was not affected by this aspect. Based on the above results, we concluded that 1) Korean pine was primarily aggregated in the core area of its distribution, and was randomly distributed at the borders or ecotones; 2) presence/absence data of specie was more likely to be implemented in analysis of population distribution type at multi-spatial scales; 3) with changing spatial scales (or extents), species distribution patterns changed, e.g., the random and uniform distribution areas at small scales often merged into aggregated distribution areas at large scales; 4) landscape pattern indices could not completely describe species distribution patterns at a single scale, but at a multi-spatial scale, stable changes in landscape indices indicated an aggregated distribution, while unstable changes resulted in uniform or random distribution; 5) Korean pine was sensitive to slope and elevation. Elucidation of the spatial patterns of Korean pine aids in understanding its distribution mechanisms, future migration, climate change effects, and promotes strategies for its effective conservation and management.
Korean pine; spatial pattern analysis; landscape indices; spatial scales; presence/absence data
国家自然科学基金项目(41371198)
2015- 09- 07;
日期:2016- 06- 14
10.5846/stxb201509071846
*通讯作者Corresponding author.E-mail: burc@iae.ac.cn
聂志文, 布仁仓, 李冰伦.红松存在/不存在数据的多尺度空间分布格局.生态学报,2017,37(3):810- 818.
Nie Z W, Bu R C, Li B L.Multi-scale spatial pattern analysis of Korean pine based on presence/absence data.Acta Ecologica Sinica,2017,37(3):810- 818.
